
Root-Zone Heat Priming Effects on Maximum Quantum Efficiency of PSII, Productivity, Root Morphology and Nutritional Quality of Two Aeroponically Grown Leafy Greens in a Tropical Greenhouse



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Maximum Quantum Efficiency of PSII Measured by Midday Chlorophyll (Chl) Fluorescence Fv/Fm Ratio and Leaf Expansion during RZ Heat Priming
2.2. Effects of High RZT on Maximum Quantum Efficiency of PSII of RZ Heat Primed and Non-Primed Plants
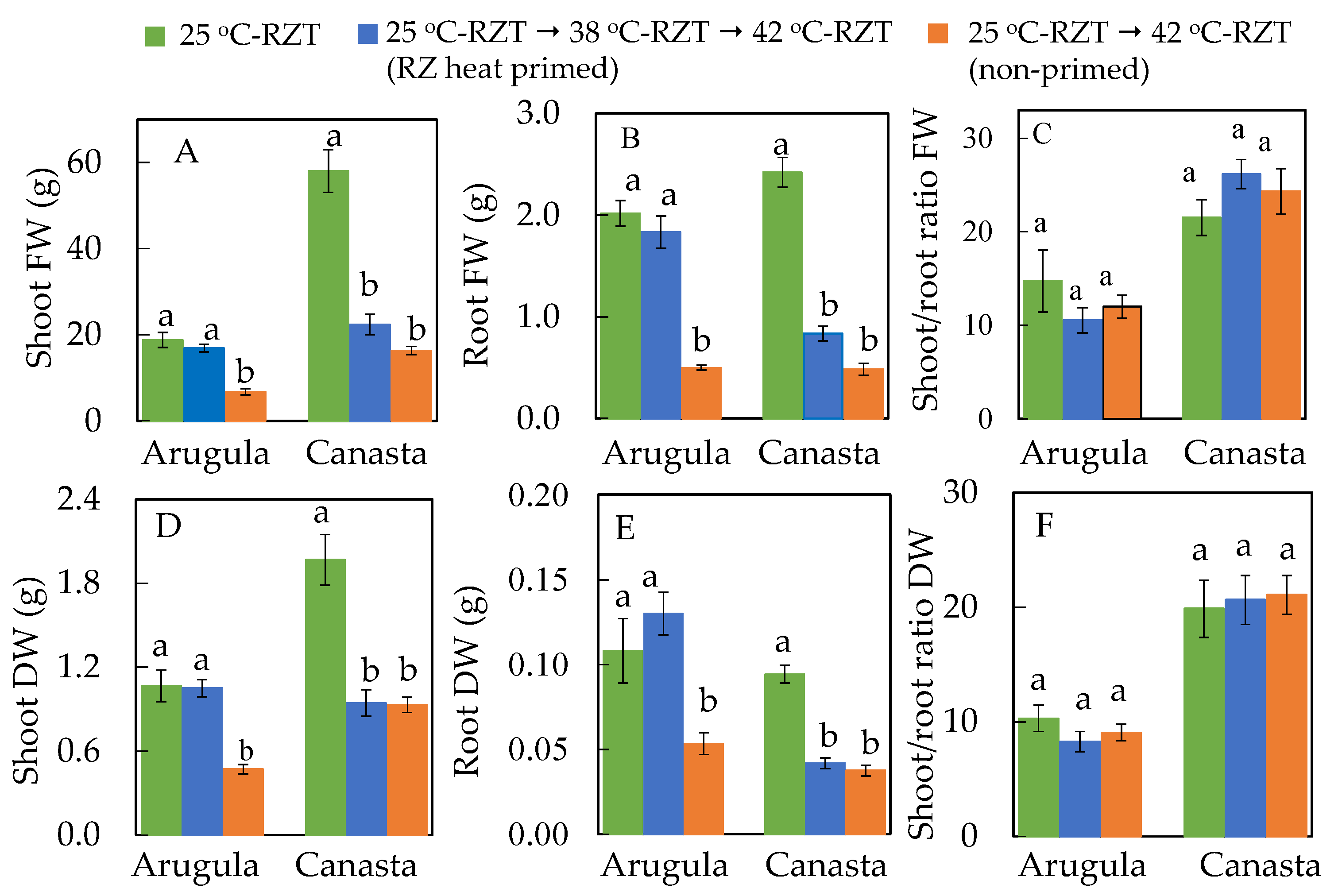
2.3. Effects of High RZT on Productivity of RZ Heat Primed Plants and Non-Primed Plants
2.4. Effects of High RZT on Root Morphology of RZ Heat Primed and Non-Primed Plants
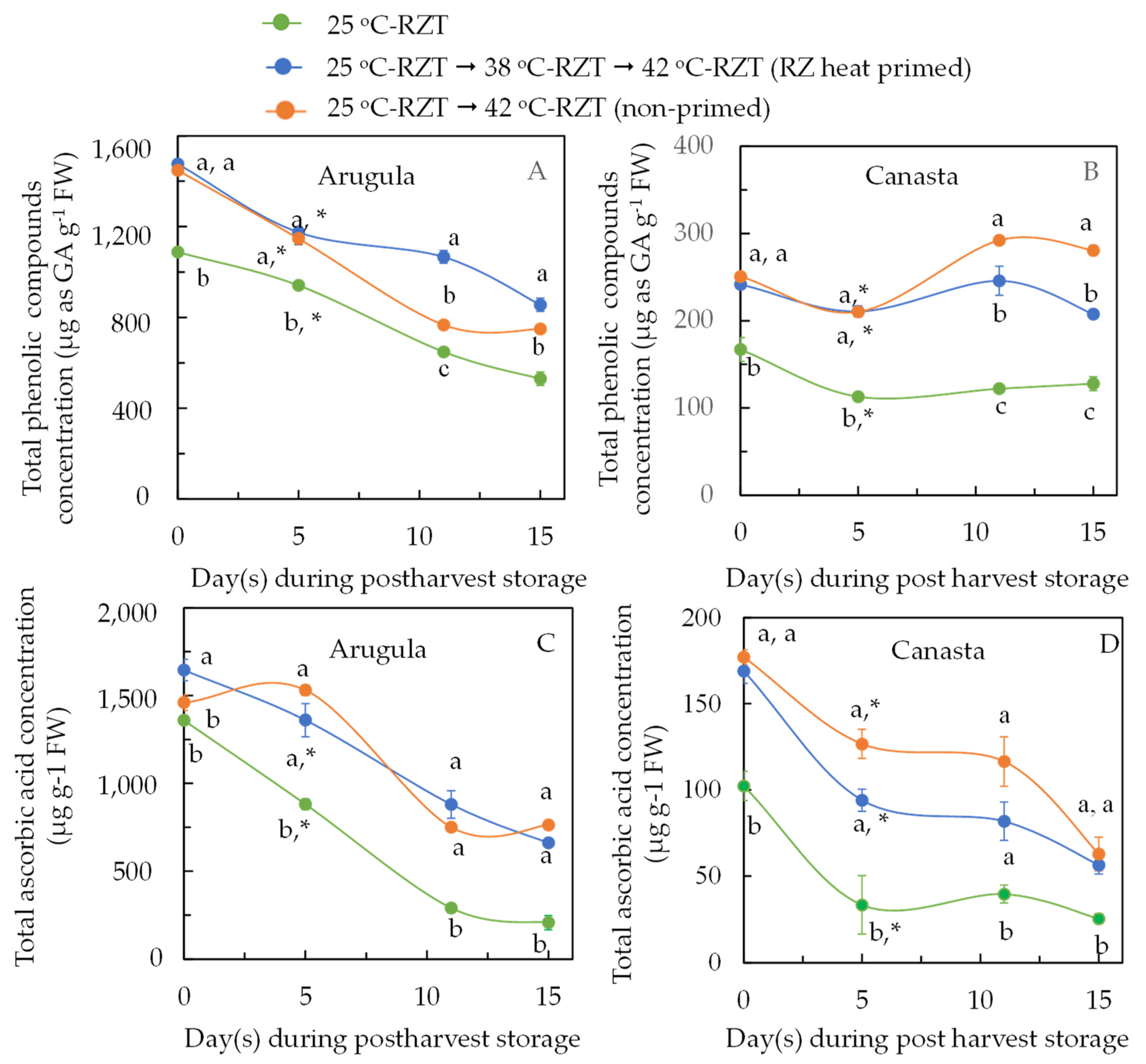
2.5. Nutritional Quality at Harvest and during Postharvest Storage of RZ Heat Primed and Non-Primed Plants after Subjecting to High RZT
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. RZT Treatments
4.3. Postharvest Shoot Sampling and Storage
4.4. Measurement of Fv/Fm Ratio
4.5. Measurement of Leaf Area Expansion
4.6. Root Morphology Analysis
4.7. Measurements of Shoot and Root Productivity
4.8. Measurements of Shoot Dietary Minerals
4.9. Determination of Total Phenolic Compounds
4.10. Determination of Total Ascorbic Acid
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheelbeek, P.F.D.; Birda, F.A.; Tuomisto, H.L.; Green, R.; Harris, F.B.; Joy, E.J.M.; Chalabi, Z.; Allen, E.; Haines, A.; Dangour, A.D. Effect of environmental changes on vegetable and legume yields and nutritional quality. Proc. Nat. Acad. Sci. USA 2018, 115, 6804–6809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and responses of chloroplasts to heat stress in plants. Front. Plant Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuletić, M.V.; Mihaljević, I.; Tomaš, V.; Horvat, D.; Zdunić, Z.; Vuković, D. Physiological response to short-term heat stress in the leaves of traditional and modern plum (Prunus domestica L.) cultivars. Horticulturae 2022, 8, 72. [Google Scholar] [CrossRef]
- Nievola, C.C.; Carvalho, C.P.; Carvalho, V.; Rodrigues, E. Rapid responses of plants to temperature changes. Temperature 2017, 4, 371–405. [Google Scholar] [CrossRef]
- Bäurle, I. Plant heat adaptation: Priming in response to heat stress. F1000Research 2016, 5, 694. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, L.; Rusalepp, L.; Kaurilind, E.; Sulaiman, H.Y.; Püssa, T.; Niinemets, Ü. Heat priming improved heat tolerance of photosynthesis, enhanced terpenoid and benzenoid emission and phenolics accumulation in Achillea millefolium. Plant Cell Environ. 2021, 44, 2365–2385. [Google Scholar] [CrossRef]
- Kim, M.; McLoughlin, F.; Eman Basha, E.; Vierling, E. Assessing plant tolerance to acute heat stress. Bio-Protocol 2017, 7, e2405. [Google Scholar] [CrossRef]
- Vierling, E. The role of heat shock proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F.L.; Jiang, D. Priming: A promising strategy for crop production in response to future climate. J. Integr. Agric. 2017, 16, 2709–2716. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Liu, F.; Jin, M.; Yu, H.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Pre-anthesis high temperature acclimation alleviates the negative effects of post-anthesis heat stress on stem stored carbohydrates remobilization and grain starch accumulation in wheat. J. Cereal Sci. 2012, 55, 331–336. [Google Scholar] [CrossRef]
- Bokszczanin, K.L.; Solanaceae Pollen Thermotolerance Initial Training Network (SPOT-ITN) Consortium; Fragkostefanakis, S. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 2013, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Suzuki, T. Effect of rootzone temperature on growth and quality of hydroponically grown red leaf lettuce (Lactuca sativa L. cv. Red Wave). Am. J. Plant Sci. 2015, 6, 2350–2360. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Lee, S.K. Growth and photosynthetic characteristics of lettuce (Lactuca sativa L.) under fluctuating hot ambient temperatures with the manipulation of cool root-zone temperature. J. Plant Physiol. 1998, 152, 387–391. [Google Scholar] [CrossRef]
- He, J.; Lee, S.K. Growth and photosynthetic responses of three aeroponically grown lettuce cultivars (Lactuca sativa L.) to different rootzone temperatures and growth irradiances under tropical aerial conditions. J. Hortic. Sci. Biotechnol. 1998, 73, 173–180. [Google Scholar] [CrossRef]
- He, J.; Lee, S.K.; Dodd, I.C. Limitations to photosynthesis of lettuce grown under tropical conditions: Alleviation by rootzone cooling. J. Exp. Bot. 2001, 52, 1323–1330. [Google Scholar] [CrossRef]
- He, J.; Lee, S.K. Relationship among photosynthesis, ribulose-1,5-bisphosphate carboxylase (Rubisco) and water relations of subtropical vegetable Chinese broccoli grown in the tropics by manipulation of root-zone temperature. Environ. Exp. Bot. 2001, 46, 119–128. [Google Scholar] [CrossRef]
- He, J. Impact of RZT on photosynthetic efficiency of aeroponically grown temperate and subtropical vegetable crops in the tropics. In Theory and Applications in Energy, Biotechnology and Nanotechnology; Buchner, T.B., Ewingen, N.H., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2009; Chapter 4; pp. 111–144. Available online: http://trove.nla.gov.au/work/27991465?versionId=45343866 (accessed on 23 March 2022).
- Zhang, J.-H.; Huang, W.-D.; Liu, Y.-P.; Pan, Q.-H. Effects of temperature acclimation pretreatment on the ultrastructure of mesophyll cells in young grape plants (Vitis vinifera L. cv. Jingxiu) under cross-temperature stresses. J. Integr. Plant Biol. 2005, 47, 959–970. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- He, J.; Lai, C.H.; Lim, Y.J.; Qin, L. Heat priming impacts on root morphology, productivity and photosynthesis of temperate vegetables crops grown in the tropics. J. Adv. Agric. Technol. 2019, 6, 14–19. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rysiak, A.; Dresler, S.; Hanaka, A.; Hawrylak-Nowak, B.; Strzemski, M.; Kováčik, J.; Sowa, I.; Latalski, M.; Wójciak, M. High temperature alters secondary metabolites and photosynthetic efficiency in Heracleum sosnowskyi. Int. J. Mol. Sci. 2021, 22, 4756. [Google Scholar] [CrossRef] [PubMed]
- Loaiza-Velarde, J.G.; Tomás-Barberán, F.A.; Salveit, M.E. Effect of intensity and duration of heat-shock treatment on wound-induced phenolic metabolism in iceberg lettuce. J. Am. Soc. Hortic. Sci. 1997, 122, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Loaiza-Velarde, J.G.; Saltveit, M.E. Heat shocks applied either before or after wounding reduce browning of lettuce leaf tissue. J. Am. Soc. Hortic. Sci. 2001, 126, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Arah, I.K.; Amaglo, H.; Kumah, E.K.; Ofori, H. Preharvest and postharvest factors affecting the quality and shelf life of harvested tomatoes: A mini review. Int. J. Agron. 2015, 2015, 478041. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.E.; Wang, C.L.; Shi, B.B.; Pan, X.J. Effect of storage temperature and time on the nutritional quality of walnut male inflorescences. J. Food Drug Anal. 2017, 25, 374–384. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Lee, S.K. Photosynthetic utilization of radiant energy by temperate lettuce grown under natural tropical condition with manipulation of root-zone temperature. Photosynthetica 2004, 42, 457–463. [Google Scholar] [CrossRef]
- Werner, C.; Correia, O.; Beyschlag, W. Characteristic patterns of chronic and dynamic photoinhibition of different functional groups in a Mediterranean ecosystem. Funct. Plant Biol. 2002, 29, 999–1011. [Google Scholar] [CrossRef]
- Kitao, M.; Harayama, H.; Yazaki, K.; Tobita, H.; Agathokleous, E.; Furuya, N.; Hashimoto, T. Photosynthetic and growth responses in a pioneer tree (Japanese white birch) and competitive perennial weeds (Eupatorium sp.) grown under different regimes with limited water supply to waterlogging. Front. Plant Sci. 2022, 13, 835068. [Google Scholar] [CrossRef]
- Lai, C.H.; He, J. Physiological performances of temperate vegetables with response to chronic and acute heat stress. Am. J. Plant Sci. 2016, 7, 2055–2071. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Ma, C.; Huang, Z.; Abid, M.; Jiang, S.; Dai, T.; Han, X. Heat priming during early reproductive stages enhances thermo-tolerance to post-anthesis heat stress via improving photosynthesis and plant productivity in winter wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 805. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Zhang, G.; Zhang, S.; Wang, Q.; Fu, Y.; Lu, H. Efficacy of root zone temperature increase in root and shoot development and hormone changes in different maize genotypes. Agriculture 2021, 11, 477. [Google Scholar] [CrossRef]
- He, J.; Tan, L.P.; Lee, S.K. Root-zone temperature effects on photosynthesis, 14C-photoassimilate partitioning and growth of temperate lettuce (Lactuca sativa cv. ‘Panama’) in the tropics. Photosynthetica 2009, 47, 95–103. [Google Scholar] [CrossRef]
- He, J.; See, X.E.; Qin, L.; Choong, T.W. Effects of root-zone temperature on photosynthesis, productivity and nutritional quality of aeroponically grown salad rocket (Eruca sativa) vegetable. Am. J. Plant Sci. 2016, 7, 1993–2005. [Google Scholar] [CrossRef] [Green Version]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.P.; He, J.; Lee, S.K. Effects of root-zone temperature on the root development and nutrient uptake of Lactuca sativa L. cv. ’Panama’ grown in an aeroponic system in the tropics. J. Plant Nutr. 2002, 25, 297–314. [Google Scholar] [CrossRef]
- Qin, L.; He, J.; Lee, S.K.; Dodd, I.C. An assessment of ethylene mediation of lettuce (Lactuca sativa) root growth at high temperatures. J. Exp. Bot. 2007, 58, 3017–3024. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, E.; Hikosaka, S.; Goto, E. Effects of nutrient solution temperature on the concentration of major bioactive compounds in red perilla. J. Agric. Meteorol. 2018, 74, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.T.P.; Lu, N.; Kagawa, N.; Takagaki, M. Optimization of photosynthetic photon flux density and root-zone temperature for enhancing secondary metabolite accumulation and production of coriander in plant factory. Agronomy 2019, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Sęczyk, Ł.; Ozdemir, E.A.; Kołodziej, B. In vitro bioaccessibility and activity of basil (Ocimum basilicum L.) phytochemicals as affected by cultivar and postharvest preservation method—Convection drying, freezing, and freeze-drying. Food Chem. 2020, 382, 132363. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Bocchini, M.; Bartucca, M.L.; Ciancaleoni, S.; Mimmo, T.; Cesco, S.; Pii, Y.; Albertini, E.; Del Buono, D. Iron deficiency in barley plants: Phytosiderophore release, iron translocation, and DNA methylation. Front. Plant Sci. 2015, 6, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragee, S.; Abdel-Aal, E.M.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Leipner, J.; Fracheboud, Y.; Stamp, P. Acclimation by suboptimal temperature diminishes photooxidative damage in maize leaves. Plant Cell Environ. 1997, 20, 366–372. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Tan, C.; Qin, L. Root-Zone Heat Priming Effects on Maximum Quantum Efficiency of PSII, Productivity, Root Morphology and Nutritional Quality of Two Aeroponically Grown Leafy Greens in a Tropical Greenhouse. Plants 2022, 11, 1684. https://doi.org/10.3390/plants11131684
He J, Tan C, Qin L. Root-Zone Heat Priming Effects on Maximum Quantum Efficiency of PSII, Productivity, Root Morphology and Nutritional Quality of Two Aeroponically Grown Leafy Greens in a Tropical Greenhouse. Plants. 2022; 11(13):1684. https://doi.org/10.3390/plants11131684
Chicago/Turabian StyleHe, Jie, Cassander Tan, and Lin Qin. 2022. "Root-Zone Heat Priming Effects on Maximum Quantum Efficiency of PSII, Productivity, Root Morphology and Nutritional Quality of Two Aeroponically Grown Leafy Greens in a Tropical Greenhouse" Plants 11, no. 13: 1684. https://doi.org/10.3390/plants11131684
APA StyleHe, J., Tan, C., & Qin, L. (2022). Root-Zone Heat Priming Effects on Maximum Quantum Efficiency of PSII, Productivity, Root Morphology and Nutritional Quality of Two Aeroponically Grown Leafy Greens in a Tropical Greenhouse. Plants, 11(13), 1684. https://doi.org/10.3390/plants11131684