
Brevilin A Isolated from Centipeda minima Induces Apoptosis in Human Gastric Cancer Cells via an Extrinsic Apoptotic Signaling Pathway


 , , and
, , and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Network Pharmacology Analysis
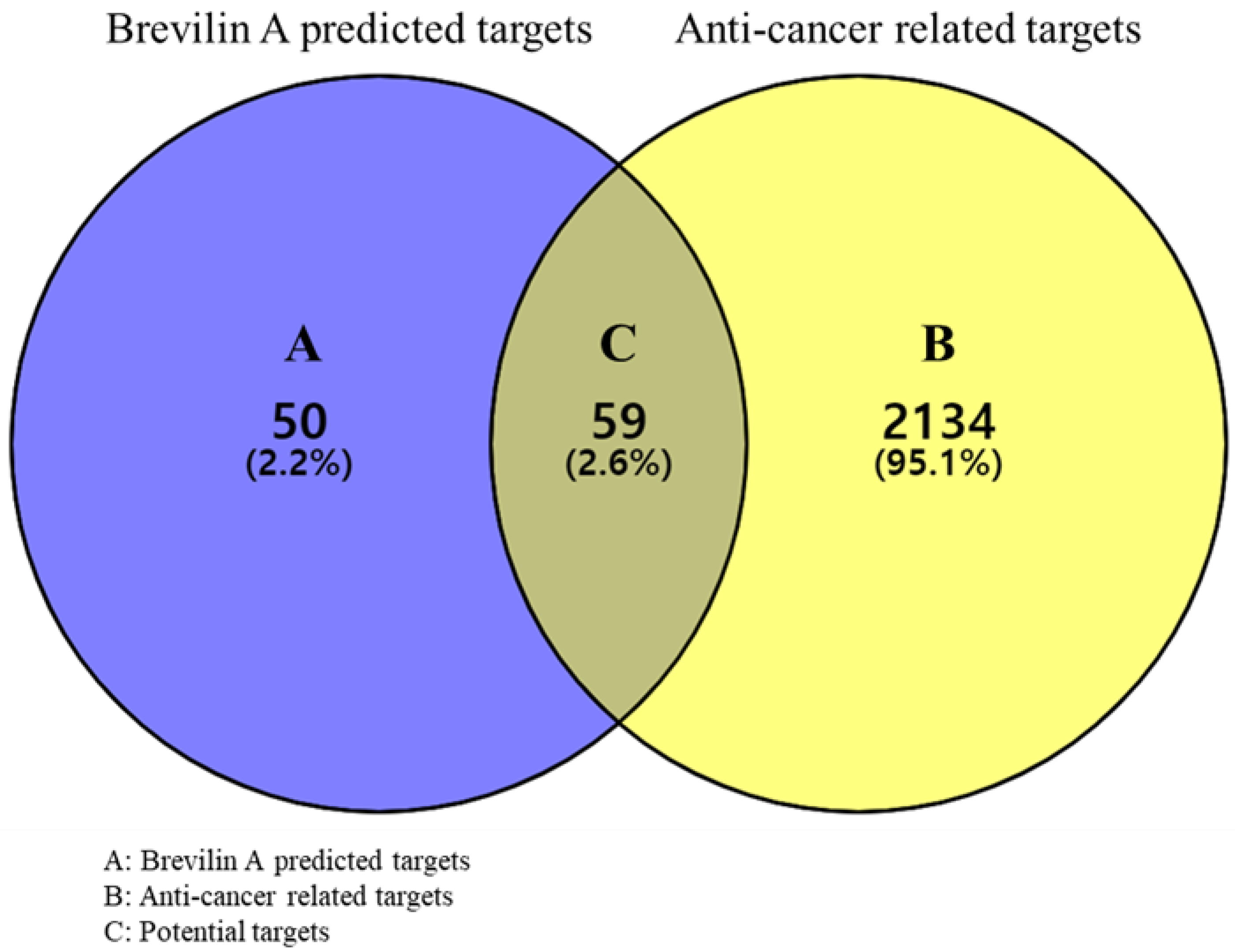
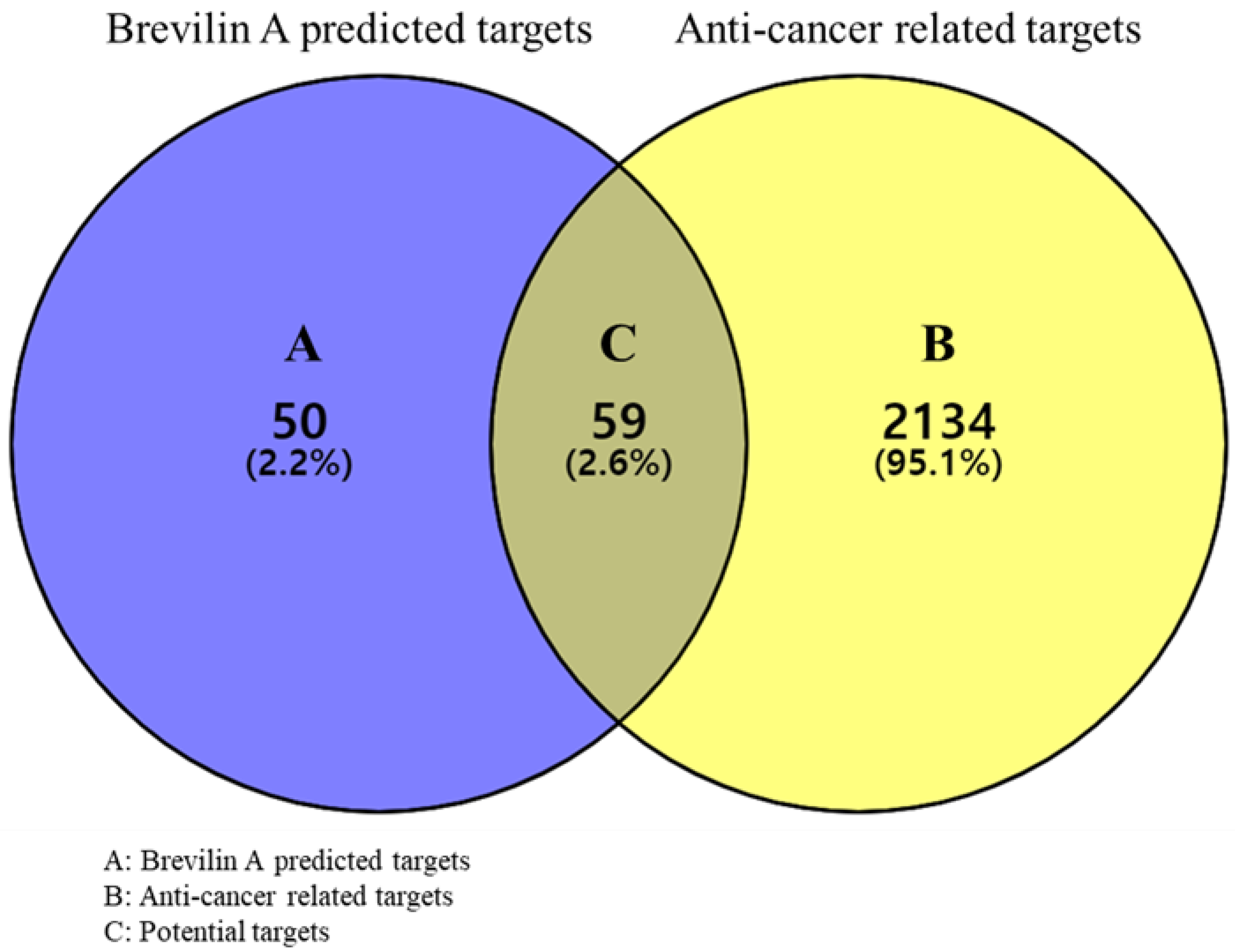
2.1.1. Prediction of Targets and Screening of Potential Targets
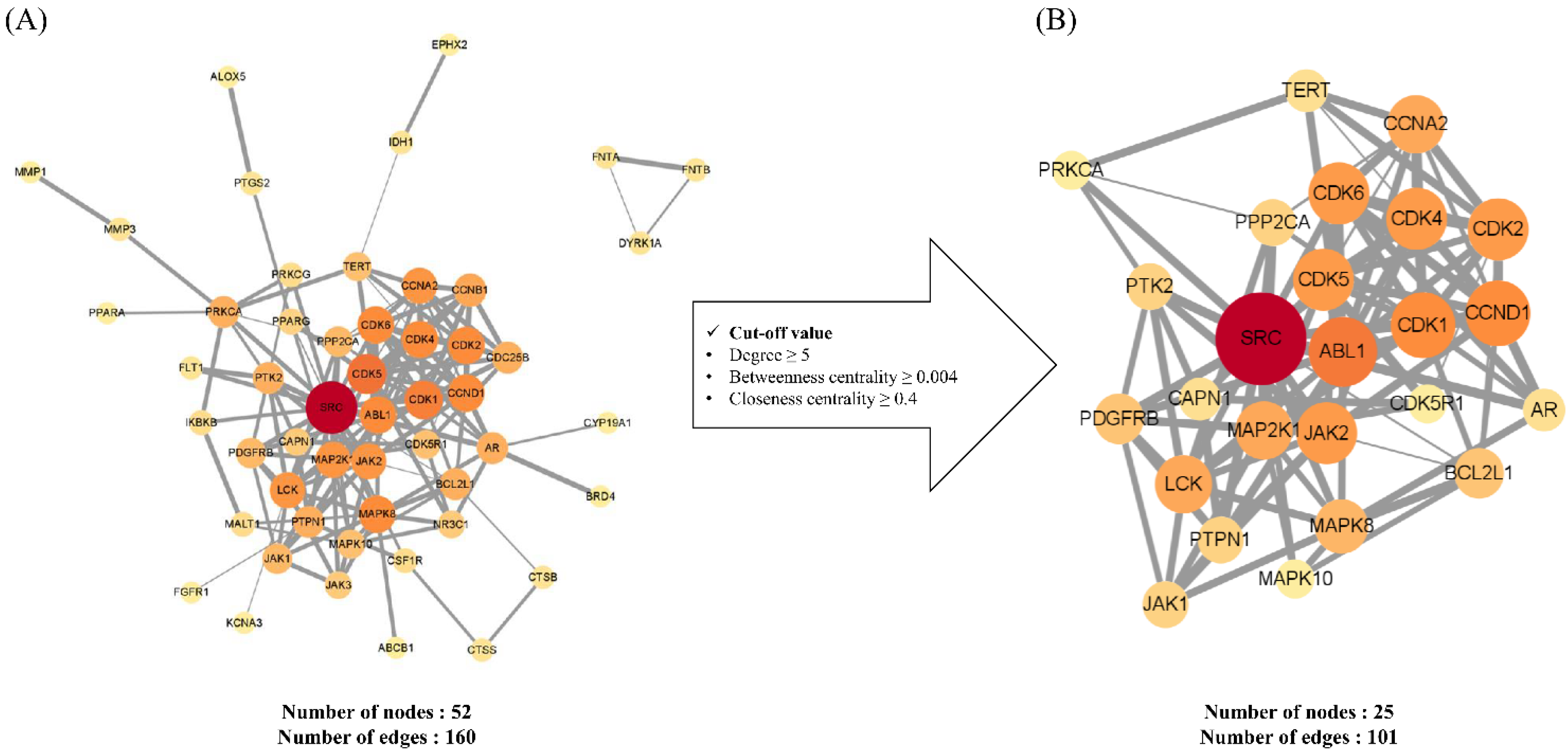
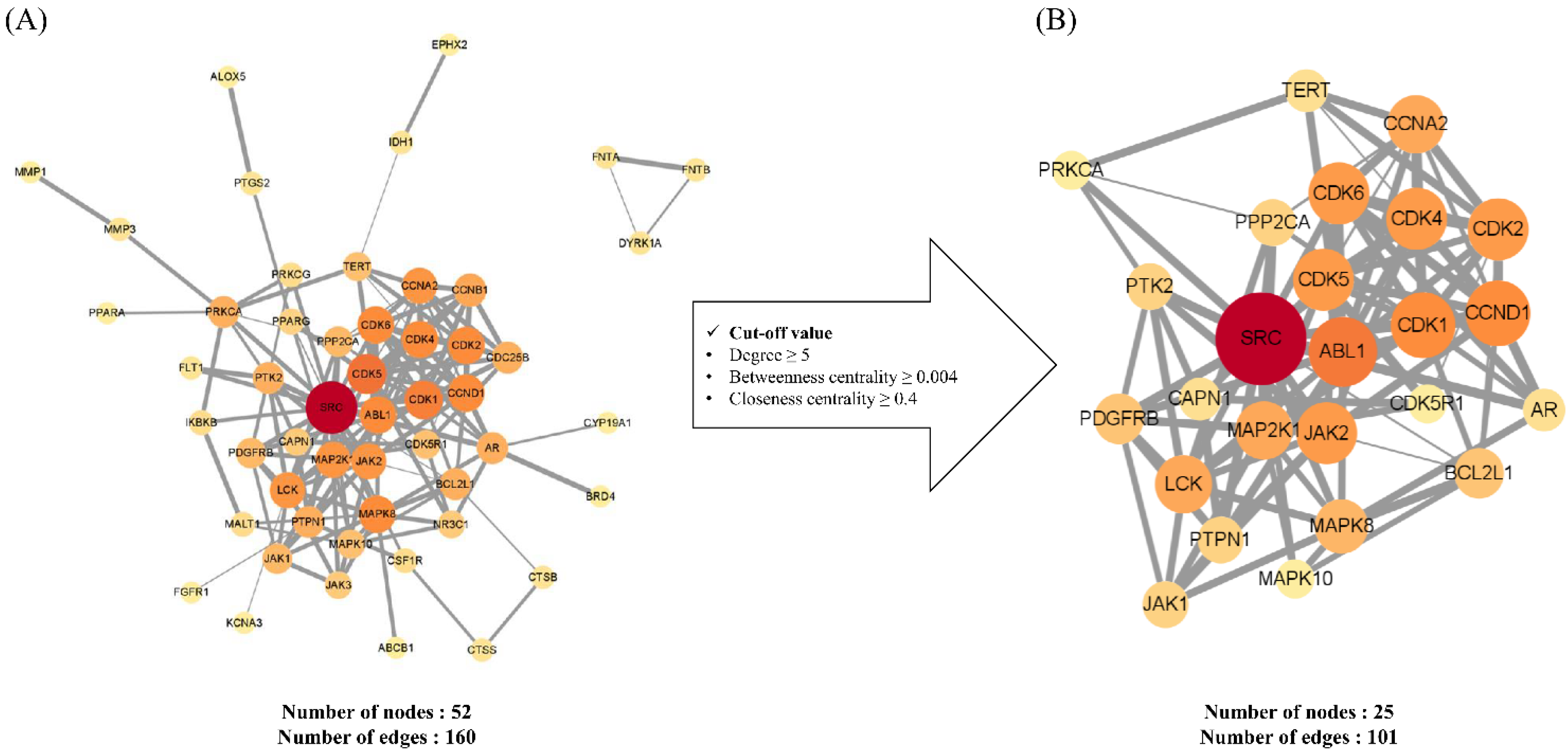
2.1.2. Construction and Analysis of Protein–Protein Interaction Networks of Potential and Key Targets
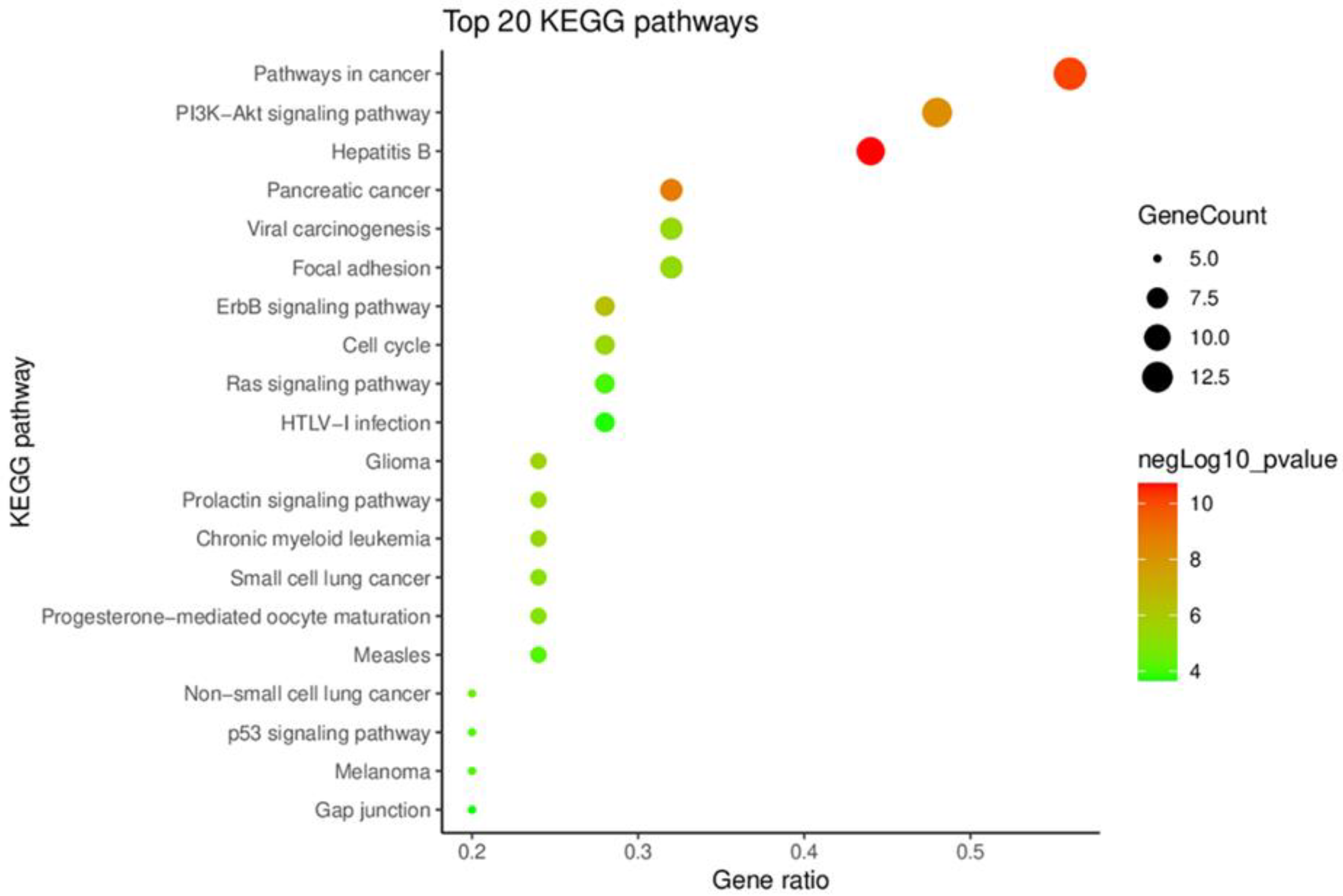
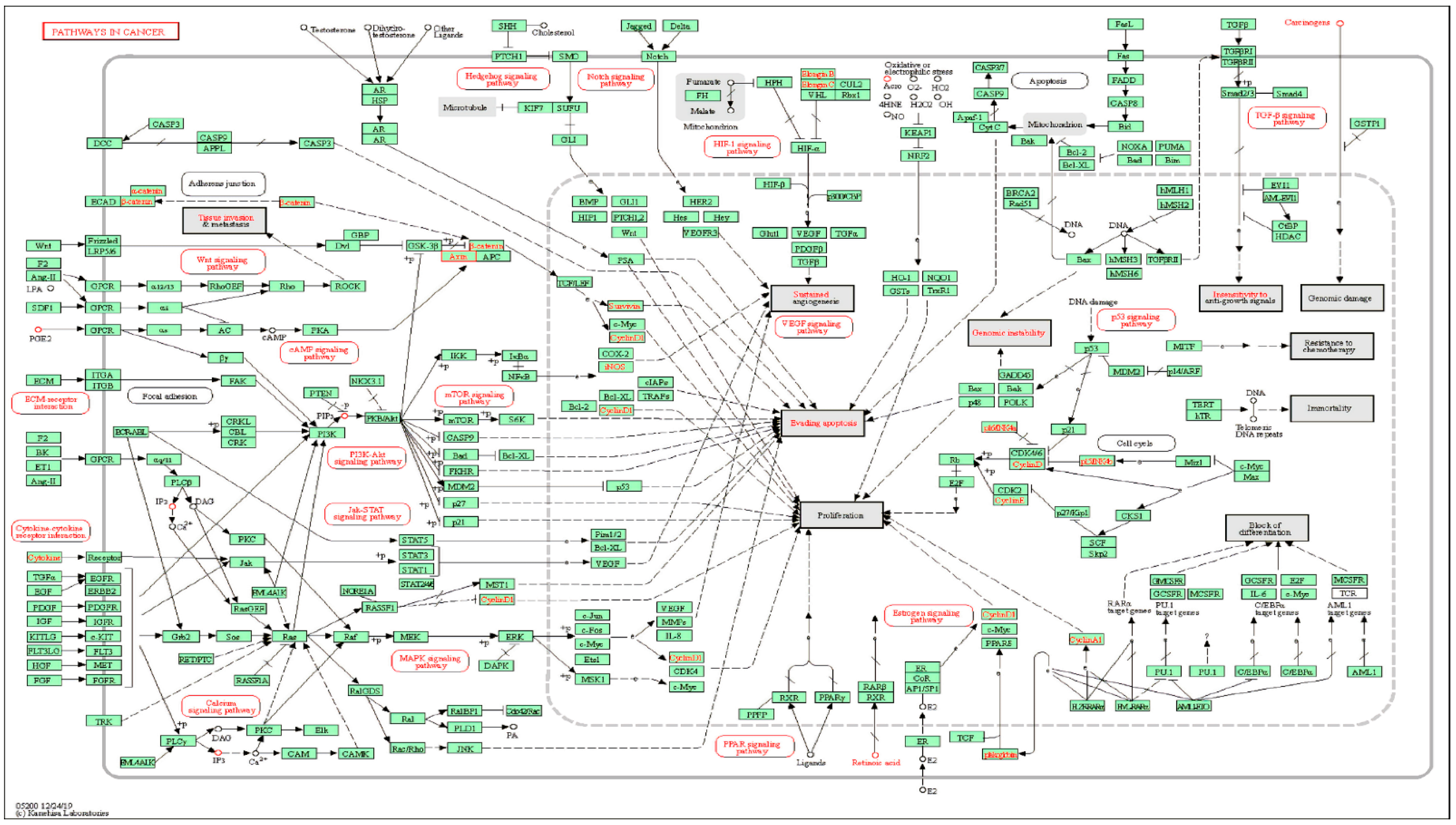
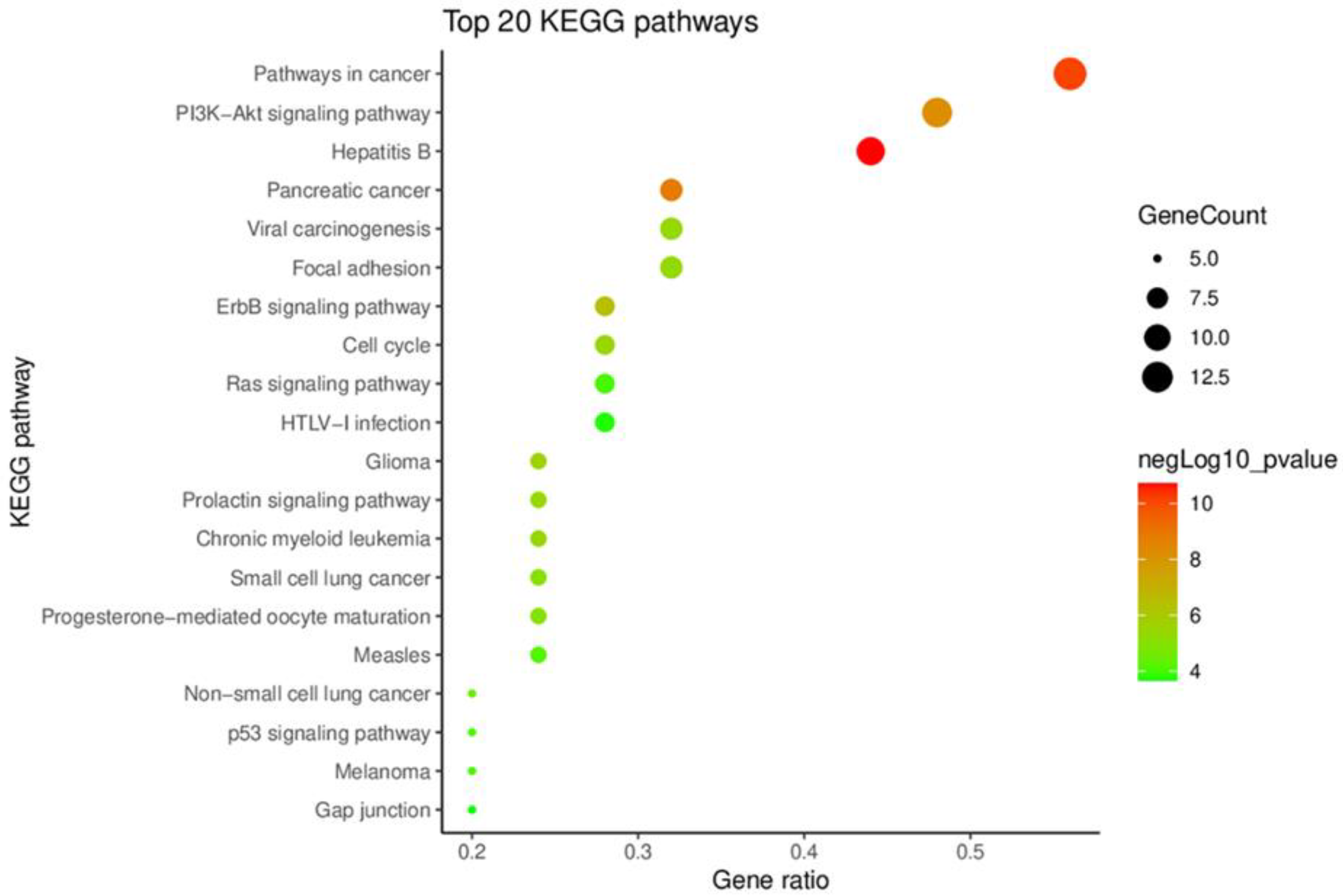
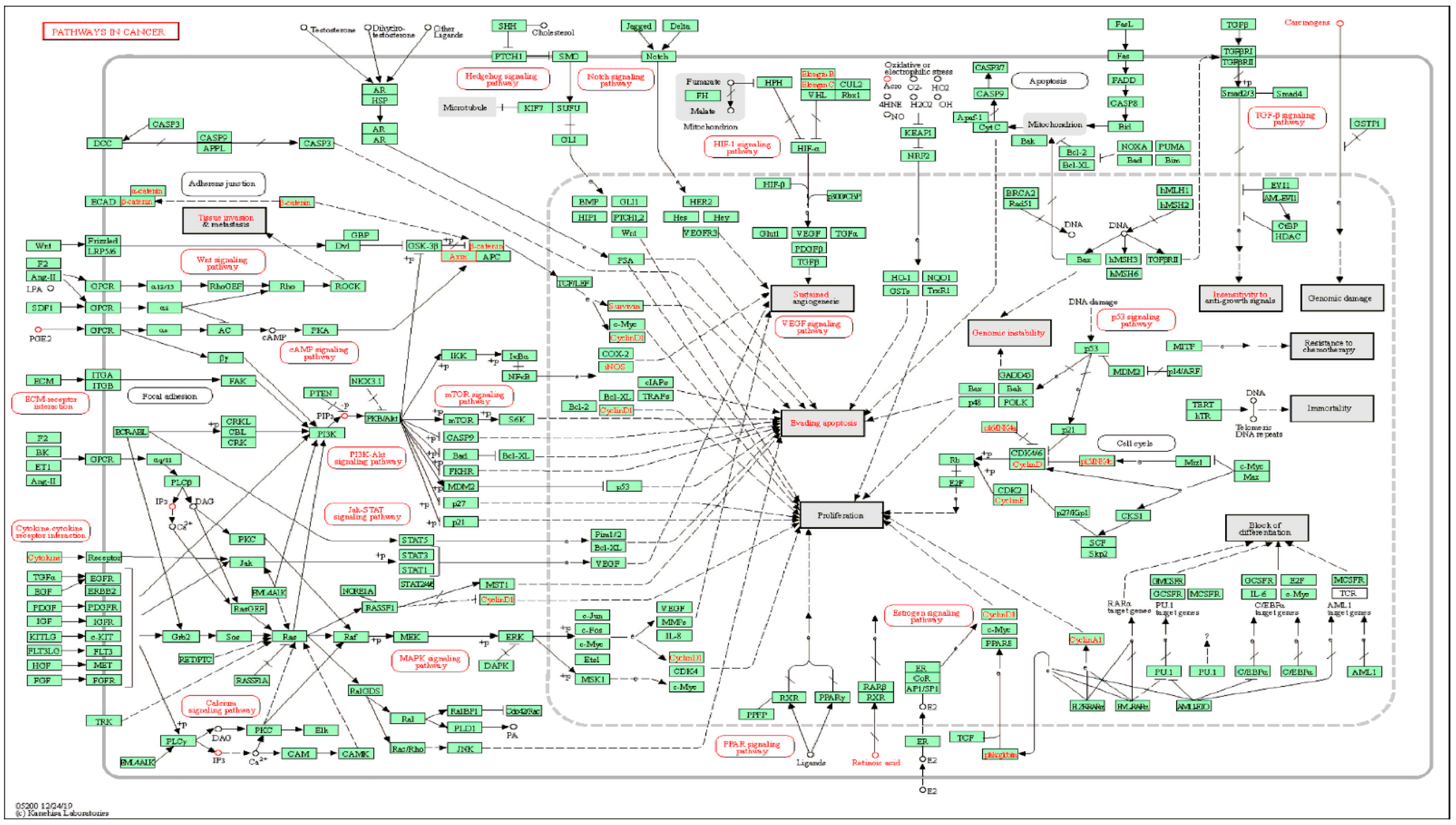
2.1.3. KEGG Pathway Enrichment Analysis of Key Targets
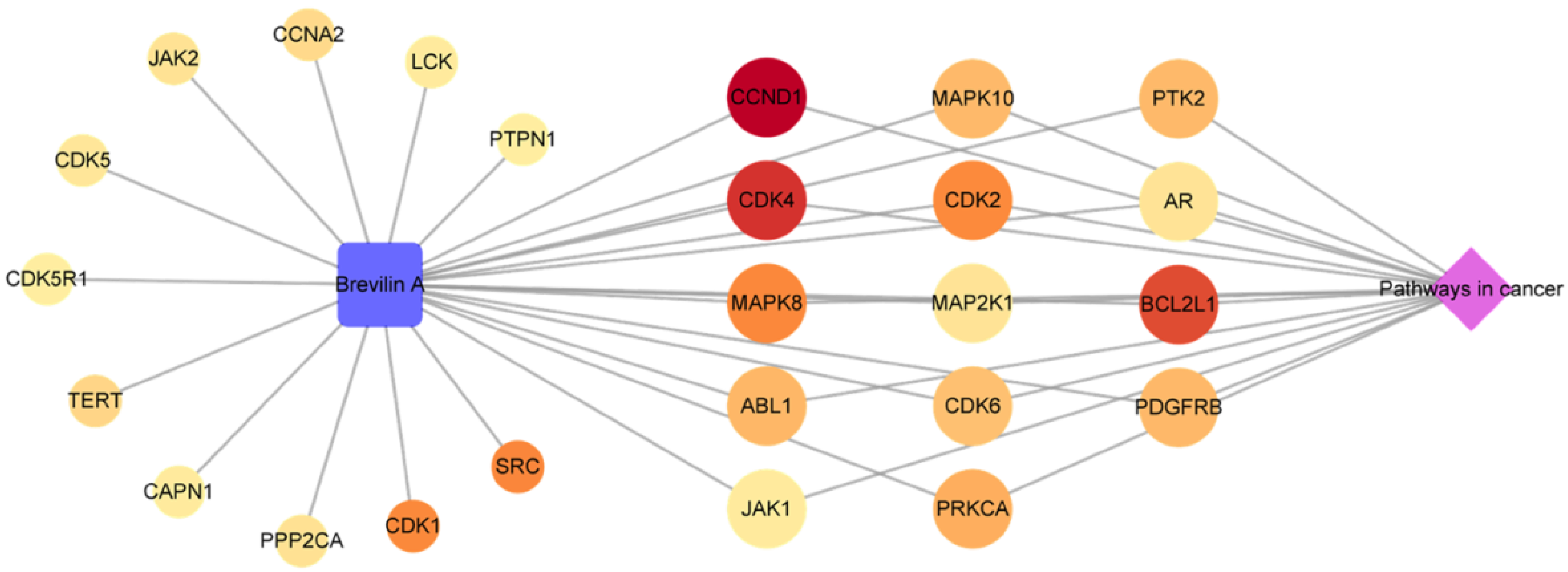
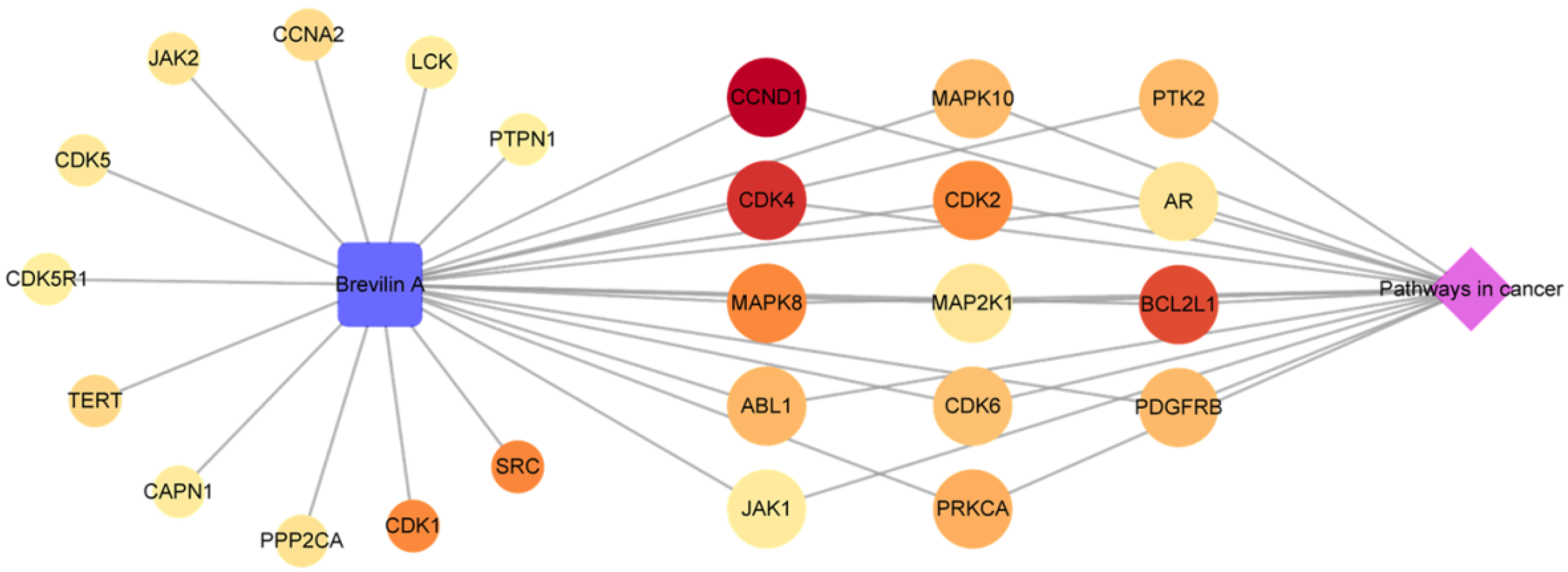
2.1.4. Analysis of a Compound-Key Target-Pathway Network
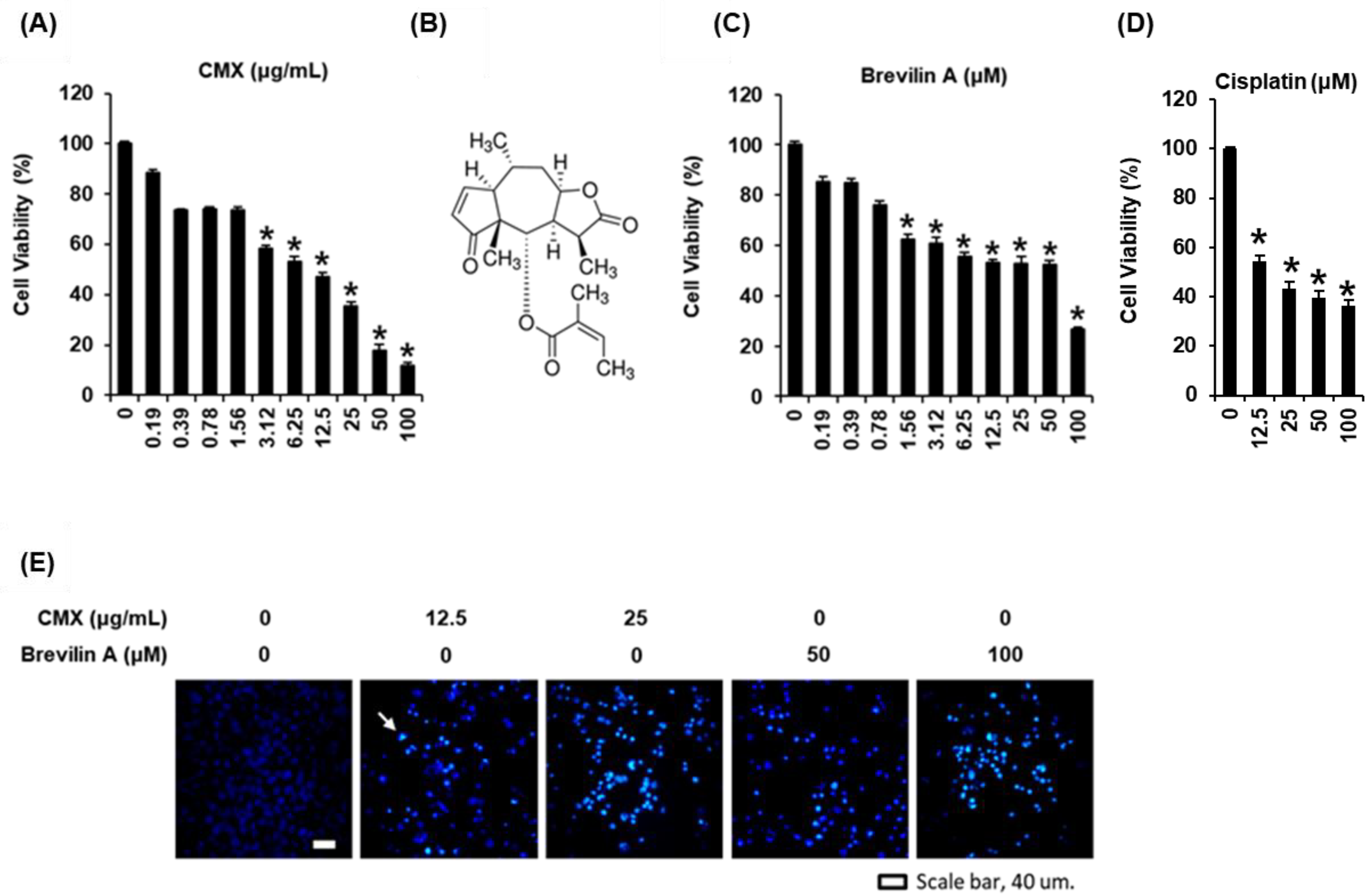
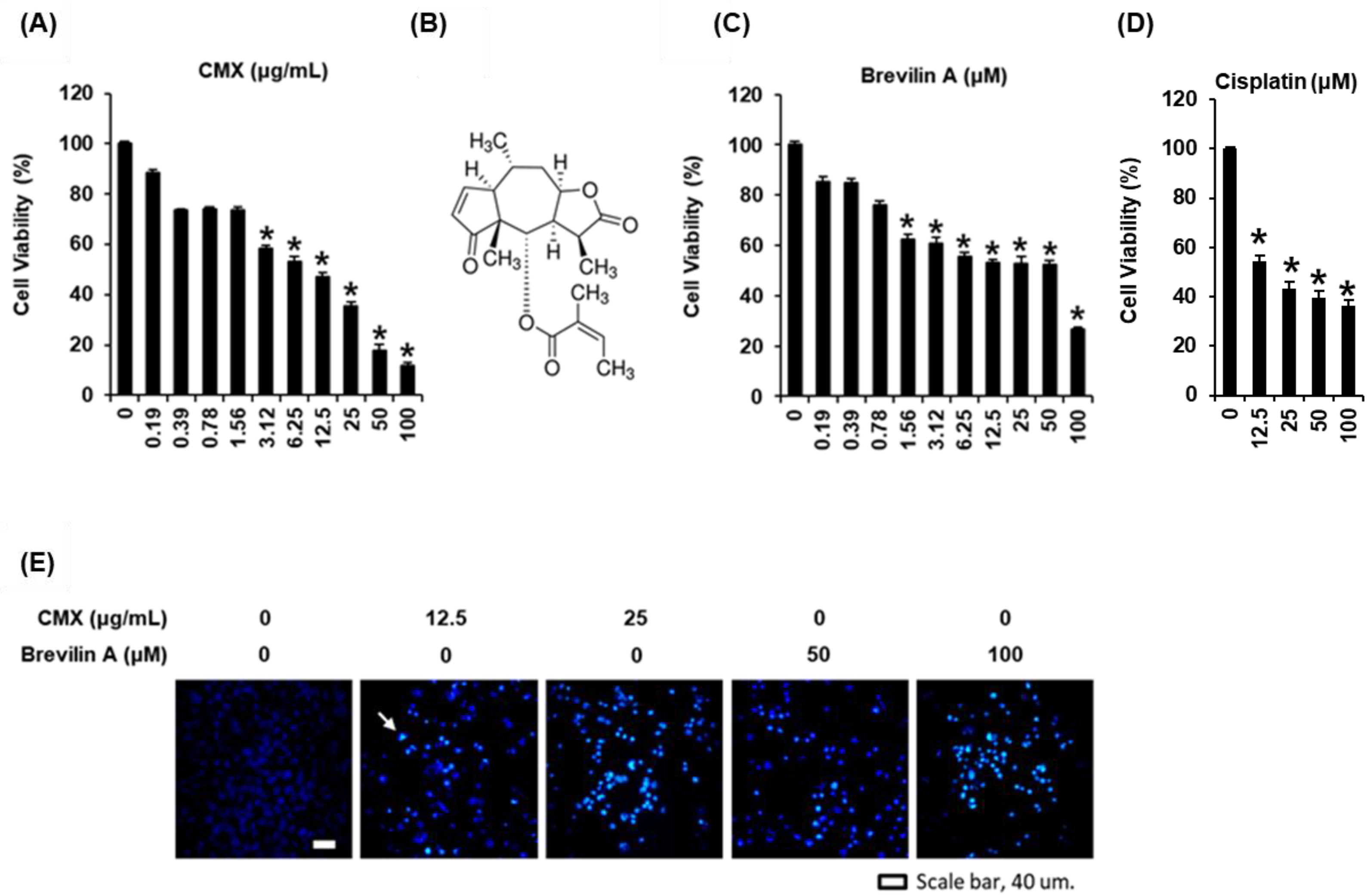
2.2. The Cytotoxic Effects of Centipeda minima (CMX) and Brevilin A on AGS Human Gastric Cancer Cells
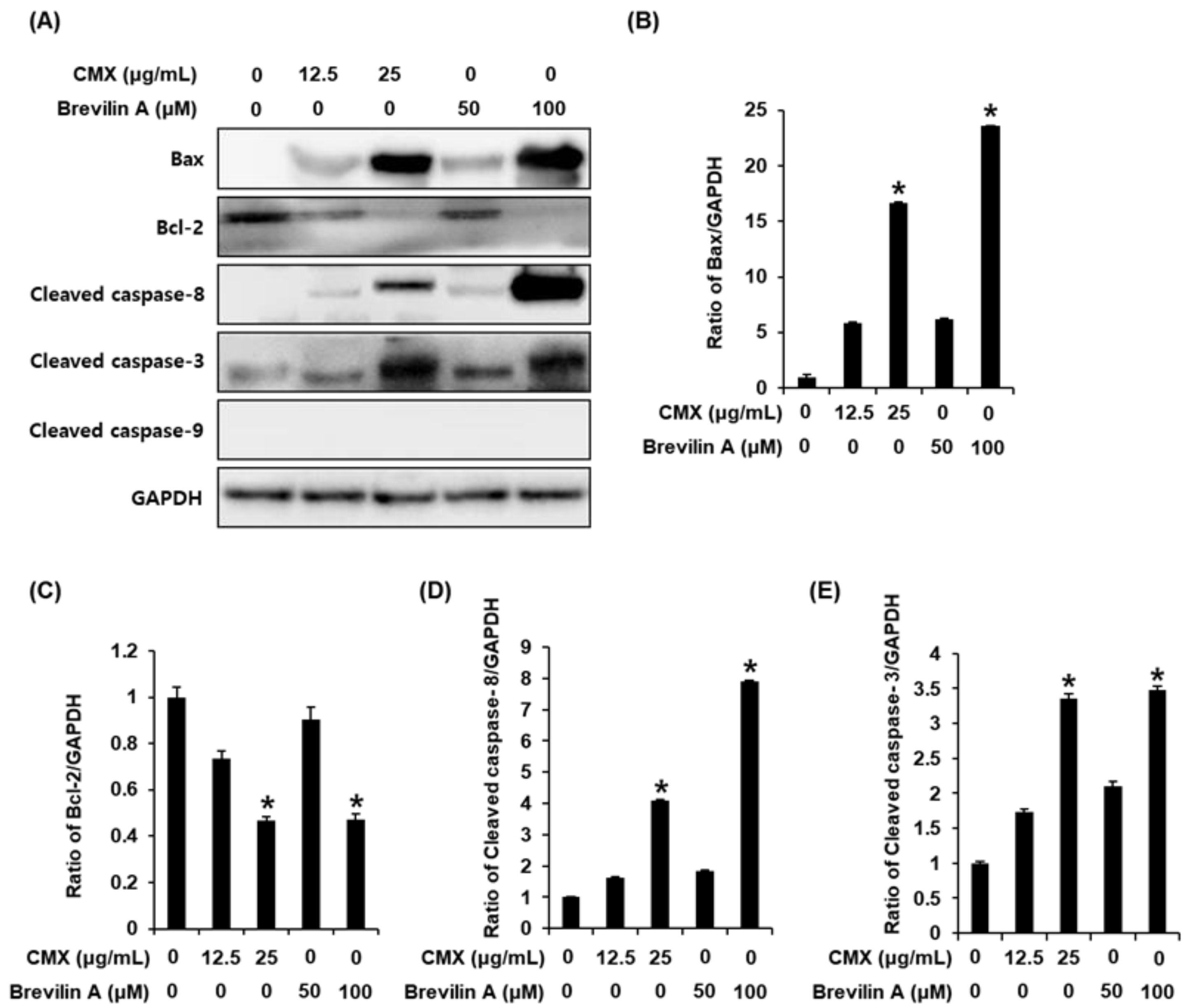
2.3. The Effects of Centipeda minima (CMX) and Brevilin A on the Expression of Apoptosis-Related Proteins in AGS Human Gastric Cancer Cells
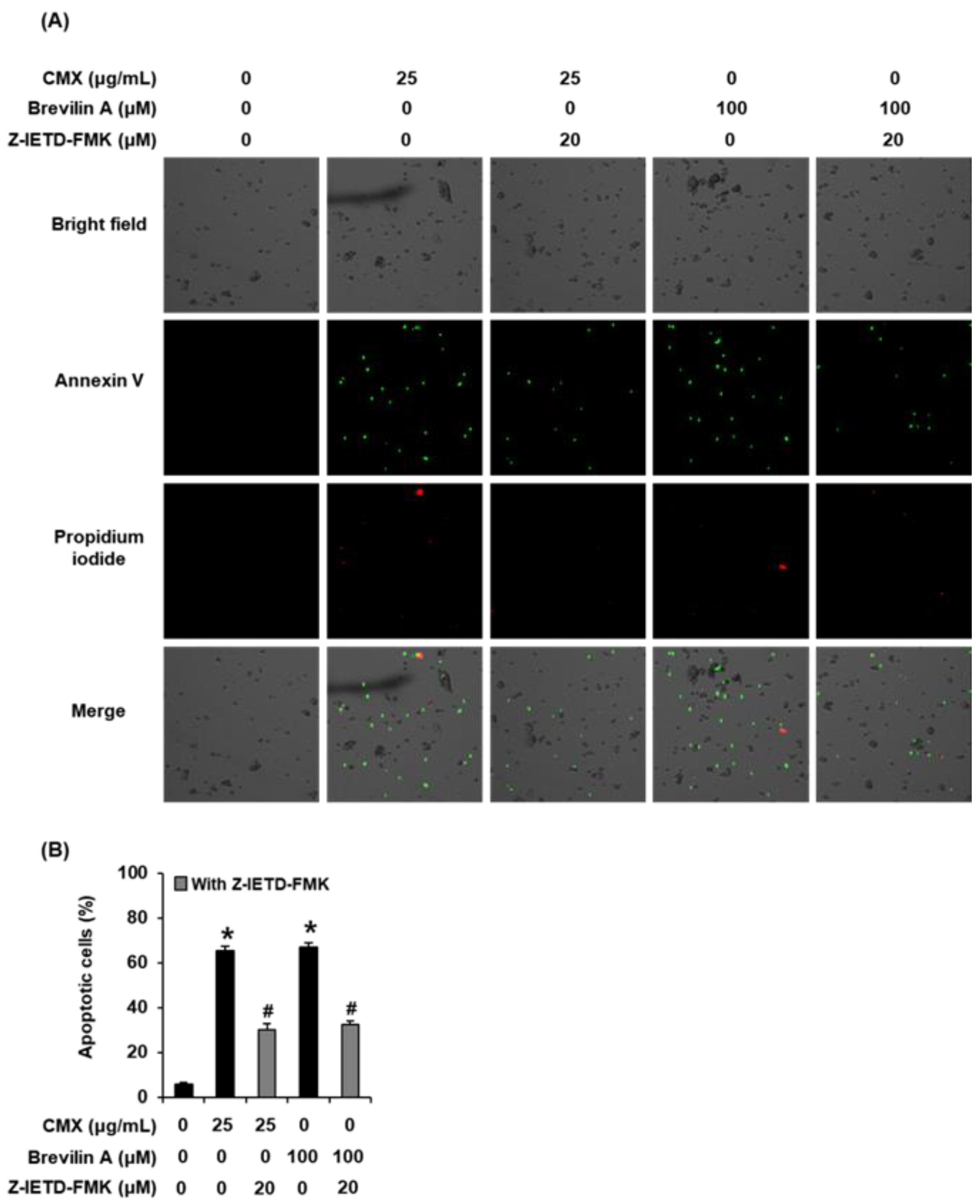
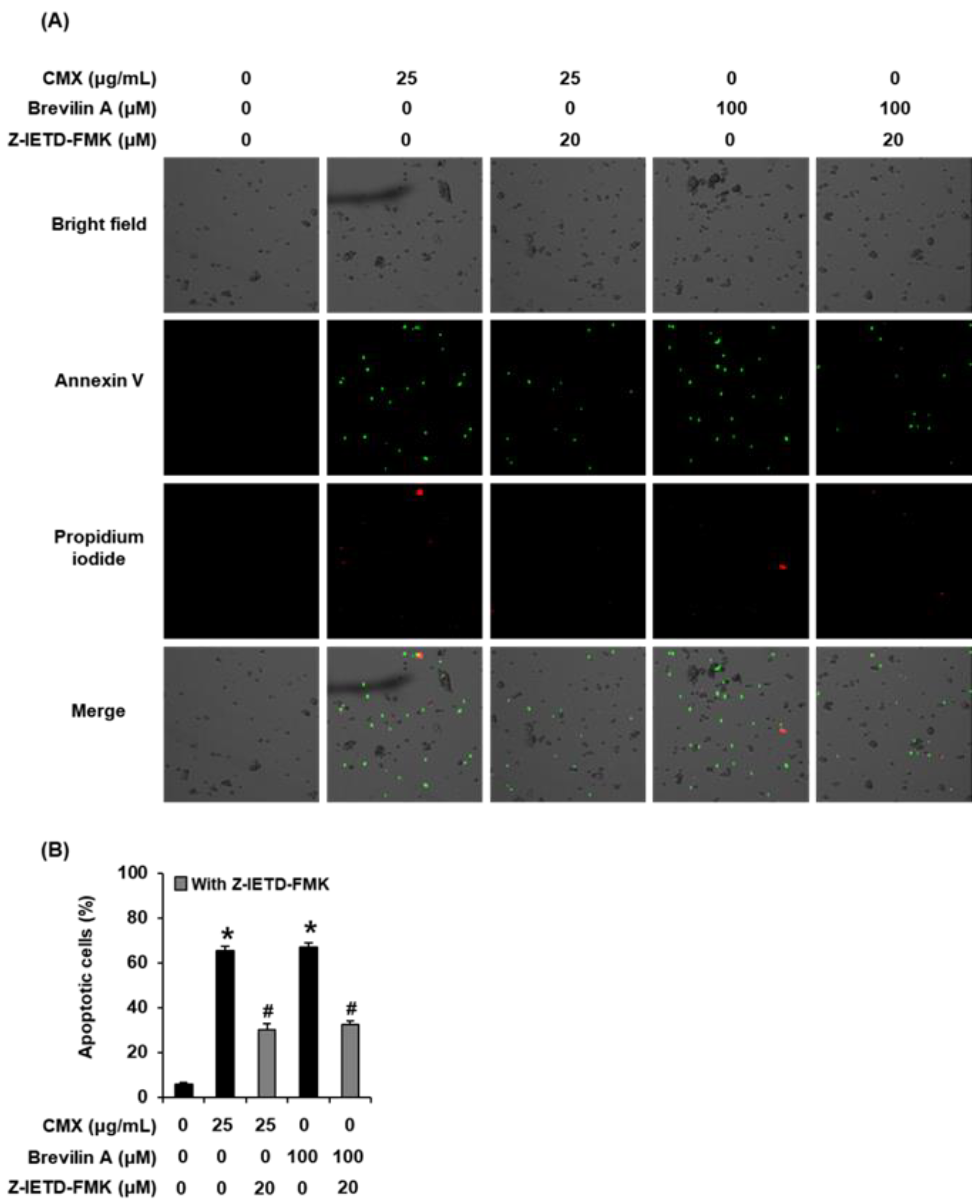
2.4. The Effects of Centipeda minima (CMX) and Brevilin A on Apoptotic Cell Death in AGS Human Gastric Cancer Cells
3. Materials and Methods
3.1. Extraction and Isolation
3.2. Network Pharmacology Analysis
3.2.1. Acquisition of the Predicted and Disease-Related Targets of Brevilin A
3.2.2. Acquisition of Potential Targets
3.2.3. Construction and Analysis of a Protein–Protein Interaction Network
3.2.4. Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Analysis
3.2.5. Construction and Analysis of a Compound-Key Target-Pathway (C-T-P) Network
3.3. Cell Culture
3.4. Ez-Cytox Assay
3.5. Cell Staining Using Hoechst 33342
3.6. Western Blot Analysis
3.7. Cell Staining with Annexin V and Propidium Iodide
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lee, M.M.-L.; Chan, B.D.; Wong, W.-Y.; Qu, Z.; Chan, M.-S.; Leung, T.-W.; Lin, Y.; Mok, D.K.-W.; Chen, S.; Tai, W.C.-S. Anti-cancer activity of Centipeda minima extract in triple negative breast cancer via inhibition of AKT, NF-κB, and STAT3 signaling pathways. Front. Oncol. 2020, 10, 491. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.-Q.; Sun, H.-Y.; Chan, C.-O.; Liu, B.-B.; Wu, J.-H.; Chan, S.-W.; Mok, D.K.-W.; Tse, A.K.-W.; Yu, Z.-L.; Chen, S.-B. Centipeda minima (Ebushicao) extract inhibits PI3K-Akt-mTOR signaling in nasopharyngeal carcinoma CNE-1 cells. Chin. Med. 2015, 10, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maqbool, M.F.; Ashaq, A.; Maryam, A.; Khan, M.; Ali, M.A.; Shakir, H.A.; Gul, S.; Shakoori, F.R.; Irfan, M.; Ara, C. Unraveling the Anticancer Components of Centipeda minima and their Cellular Targets in Human Cancers. J. Biol. Act. Prod. Nat. 2022, 12, 42–64. [Google Scholar] [CrossRef]
- Liu, R.; Dow Chan, B.; Mok, D.K.-W.; Lee, C.-S.; Tai, W.C.-S.; Chen, S. Arnicolide D, from the herb Centipeda minima, is a therapeutic candidate against nasopharyngeal carcinoma. Molecules 2019, 24, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.M.-L.; Chan, B.D.; Wong, W.-Y.; Leung, T.-W.; Qu, Z.; Huang, J.; Zhu, L.; Lee, C.-S.; Chen, S.; Tai, W.C.-S. Synthesis and evaluation of novel anticancer compounds derived from the natural product Brevilin A. ACS Omega 2020, 5, 14586–14596. [Google Scholar] [CrossRef]
- Qin, Y.; Lu, H. In vitro evaluation of anti-hepatoma activity of brevilin A: Involvement of Stat3/Snail and Wnt/β-catenin pathways. RSC Adv. 2019, 9, 4390–4396. [Google Scholar] [CrossRef] [Green Version]
- Su, T.; Wang, Y.-P.; Wang, X.-N.; Li, C.-Y.; Zhu, P.-L.; Huang, Y.-M.; Yang, Z.-Y.; Chen, S.-B.; Yu, Z.-L. The JAK2/STAT3 pathway is involved in the anti-melanoma effects of brevilin A. Life Sci. 2020, 241, 117169. [Google Scholar] [CrossRef]
- Zhang, X.; Xia, Y.; Yang, L.; He, J.; Li, Y.; Xia, C. Brevilin a, a sesquiterpene lactone, inhibits the replication of influenza a virus in vitro and in vivo. Viruses 2019, 11, 835. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Wright, C.; Cai, Y.; Yang, S.; Phillipson, J.; Kirby, G.; Warhurst, D. Antiprotozoal activities of Centipeda minima. Phytother. Res. 1994, 8, 436–438. [Google Scholar] [CrossRef]
- Qin, Q.; Xu, G.; Zhan, X.; Wang, Z.; Wang, Y.; Liu, H.; Hou, X.; Shi, W.; Ma, J.; Bai, Z. Brevilin A inhibits NLRP3 inflammasome activation in vivo and in vitro by acting on the upstream of NLRP3-induced ASC oligomerization. Mol. Immunol. 2021, 135, 116–126. [Google Scholar] [CrossRef]
- Kim, B.H.; Lee, M.J.; Lee, W.Y.; Pyo, J.; Shin, M.S.; Hwang, G.S.; Shin, D.; Kim, C.E.; Park, E.S.; Kang, K.S. Hair Growth Stimulation Effect of Centipeda minima Extract: Identification of Active Compounds and Anagen-Activating Signaling Pathways. Biomolecules 2021, 11, 976. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kwak, H.J.; Kim, B.H.; Kim, S.H.; Kim, D.-W.; Kang, K.S. Combined Anti-Adipogenic Effects of Hispidulin and p-Synephrine on 3T3-L1 Adipocytes. Biomolecules 2021, 11, 1764. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Wang, T.; Shi, C.; Wang, Z.; Fu, X. Network pharmacology-based approach to understand the effect and mechanism of Danshen against anemia. J. Ethnopharmacol. 2022, 282, 114615. [Google Scholar] [CrossRef] [PubMed]
- Golbeck, J. Introduction to Social Media Investigation: A Hands-On Approach; Syngress: Rockland, MA, USA, 2015. [Google Scholar]
- Hansen, D.; Shneiderman, B.; Smith, M.A. Analyzing Social Media Networks with NodeXL: Insights from a Connected World; Morgan Kaufmann: Los Angeles, CA, USA, 2010. [Google Scholar]
- Metcalf, L.; Casey, W. Cybersecurity and Applied Mathematics; Syngress: Rockland, MA, USA, 2016. [Google Scholar]
- Zhang, S.; Yu, D. Targeting Src family kinases in anti-cancer therapies: Turning promise into triumph. Trends Pharmacol. Sci. 2012, 33, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Simatou, A.; Simatos, G.; Goulielmaki, M.; Spandidos, D.A.; Baliou, S.; Zoumpourlis, V. Historical retrospective of the SRC oncogene and new perspectives. Mol. Clin. Oncol. 2020, 13, 1–10. [Google Scholar] [CrossRef]
- Montalto, F.I.; De Amicis, F. Cyclin D1 in cancer: A molecular connection for cell cycle control, adhesion and invasion in tumor and stroma. Cells 2020, 9, 2648. [Google Scholar] [CrossRef]
- Chou, J.; Quigley, D.A.; Robinson, T.M.; Feng, F.Y.; Ashworth, A. Transcription-associated cyclin-dependent kinases as targets and biomarkers for cancer therapy. Cancer Discov. 2020, 10, 351–370. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Zhu, Y.; Mu, R.; Xu, J.; Zhang, X.; Wang, C.; Li, Q.; Huang, Z.; Zhang, J.; Pan, Y. A Jak2-selective inhibitor potently reverses the immune suppression by modulating the tumor microenvironment for cancer immunotherapy. Biochem. Pharmacol. 2017, 145, 132–146. [Google Scholar] [CrossRef]
- Luttman, J.H.; Colemon, A.; Mayro, B.; Pendergast, A.M. Role of the ABL tyrosine kinases in the epithelial–mesenchymal transition and the metastatic cascade. Cell Commun. Signal. 2021, 19, 1–16. [Google Scholar] [CrossRef]
- Dey, N.; De, P.; Leyland-Jones, B. PI3K-AKT-mTOR inhibitors in breast cancers: From tumor cell signaling to clinical trials. Pharmacol. Ther. 2017, 175, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Polivka, J., Jr.; Janku, F. Molecular targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol. Ther. 2014, 142, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.J.; Snowden, J.A.; Zeidler, M.P.; Danson, S.J. The role of JAK/STAT signalling in the pathogenesis, prognosis and treatment of solid tumours. Br. J. Cancer 2015, 113, 365–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; He, G.; Hou, M.; Chen, L.; Chen, S.; Xu, A.; Fu, Y. Cell cycle regulation by alternative polyadenylation of CCND1. Sci. Rep. 2018, 8, 6824. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.J.; Reddy, E.P. CDK4: A key player in the cell cycle, development, and cancer. Genes Cancer 2012, 3, 658–669. [Google Scholar] [CrossRef] [Green Version]
- Shan, Y.S.; Hsu, H.P.; Lai, M.D.; Hung, Y.H.; Wang, C.Y.; Yen, M.C.; Chen, Y.L. Cyclin D1 overexpression correlates with poor tumor differentiation and prognosis in gastric cancer. Oncol. Lett. 2017, 14, 4517–4526. [Google Scholar] [CrossRef] [Green Version]
- Sillars-Hardebol, A.H.; Carvalho, B.; Beliën, J.A.; de Wit, M.; Delis-van Diemen, P.M.; Tijssen, M.; van de Wiel, M.A.; Pontén, F.; Fijneman, R.J.; Meijer, G.A. BCL2L1 has a functional role in colorectal cancer and its protein expression is associated with chromosome 20q gain. J. Pathol. 2012, 226, 442–450. [Google Scholar] [CrossRef]
- Elbadawy, M.; Usui, T.; Yamawaki, H.; Sasaki, K. Development of an experimental model for analyzing drug resistance in colorectal cancer. Cancers 2018, 10, 164. [Google Scholar] [CrossRef] [Green Version]
- Trisciuoglio, D.; Tupone, M.G.; Desideri, M.; Di Martile, M.; Gabellini, C.; Buglioni, S.; Pallocca, M.; Alessandrini, G.; D’Aguanno, S.; Del Bufalo, D. BCL-X L overexpression promotes tumor progression-associated properties. Cell Death Dis. 2017, 8, 3216. [Google Scholar] [CrossRef] [Green Version]
- Toné, S.; Sugimoto, K.; Tanda, K.; Suda, T.; Uehira, K.; Kanouchi, H.; Samejima, K.; Minatogawa, Y.; Earnshaw, W.C. Three distinct stages of apoptotic nuclear condensation revealed by time-lapse imaging, biochemical and electron microscopy analysis of cell-free apoptosis. Exp. Cell Res. 2007, 313, 3635–3644. [Google Scholar] [CrossRef] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Zhang, J.; O’Neill, K.L.; Gurumurthy, C.B.; Quadros, R.M.; Tu, Y.; Luo, X. Cleavage by caspase 8 and mitochondrial membrane association activate the BH3-only protein Bid during TRAIL-induced apoptosis. J. Biol. Chem. 2016, 291, 11843–11851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish, A.B.; Freel, C.D.; Kornbluth, S. Cellular mechanisms controlling caspase activation and function. Cold Spring Harb. Perspect. Biol. 2013, 5, a008672. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [PubMed]
- Gfeller, D.; Michielin, O.; Zoete, V. Shaping the interaction landscape of bioactive molecules. Bioinformatics 2013, 29, 3073–3079. [Google Scholar] [CrossRef] [PubMed]
- Venny, O.J.C. An Interactive Tool for Comparing Lists with Venn Diagrams. 2007. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 18 August 2021).
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | UniProt ID | Gene | Targets | Protein Class |
|---|---|---|---|---|
| 1 | P08183 | ABCB1 | ATP binding cassette subfamily B member 1 | Transporter |
| 2 | P09874 | PARP1 | poly (ADP-ribose) polymerase 1 | - |
| 3 | P24385 | CCND1 | cyclin D1 | Enzyme modulator |
| 4 | P11802 | CDK4 | cyclin-dependent kinase 4 | Kinase |
| 5 | Q07817 | BCL2L1 | BCL2 like 1 | Signaling |
| 6 | P35354 | PTGS2 | prostaglandin-endoperoxide synthase 2 | Enzyme |
| 7 | P12931 | SRC | SRC proto-oncogene, non-receptor tyrosine kinase | Kinase |
| 8 | P45983 | MAPK8 | mitogen-activated protein kinase 8 | Kinase |
| 9 | P06493 | CDK1 | cyclin-dependent kinase 1 | Kinase |
| 10 | P24941 | CDK2 | cyclin-dependent kinase 2 | Kinase |
| 11 | P09917 | ALOX5 | arachidonate 5-lipoxygenase | Enzyme |
| 12 | P11511 | CYP19A1 | cytochrome P450 family 19 subfamily A member 1 | Enzyme |
| 13 | P52333 | JAK3 | Janus kinase 3 | Kinase |
| 14 | P17252 | PRKCA | protein kinase C alpha | Kinase |
| 15 | O14920 | IKBKB | inhibitor of nuclear factor-kappa B kinase subunit beta | Kinase |
| 16 | P14635 | CCNB1 | cyclin B1 | Enzyme modulator |
| 17 | P00519 | ABL1 | ABL proto-oncogene 1, non-receptor tyrosine kinase | Kinase |
| 18 | P09619 | PDGFRB | platelet-derived growth factor receptor beta | Kinase |
| 19 | P53779 | MAPK10 | mitogen-activated protein kinase 10 | Kinase |
| 20 | Q05397 | PTK2 | protein tyrosine kinase 2 | Kinase |
| 21 | P37231 | PPARG | peroxisome proliferator-activated receptor gamma | Nuclear receptor |
| 22 | Q07869 | PPARA | peroxisome proliferator-activated receptor alpha | Nuclear receptor |
| 23 | P07858 | CTSB | cathepsin B | Enzyme |
| 24 | Q00534 | CDK6 | cyclin-dependent kinase 6 | Kinase |
| 25 | Q9NR96 | TLR9 | toll-like receptor 9 | - |
| 26 | P17948 | FLT1 | fms-related receptor tyrosine kinase 1 | Kinase |
| 27 | P04150 | NR3C1 | nuclear receptor subfamily 3 group C member 1 | Nuclear receptor |
| 28 | P05129 | PRKCG | protein kinase C gamma | Kinase |
| 29 | P30305 | CDC25B | cell division cycle 25B | Enzyme |
| 30 | O14746 | TERT | telomerase reverse transcriptase | Enzyme |
| 31 | P20248 | CCNA2 | cyclin A2 | Enzyme modulator |
| 32 | P34913 | EPHX2 | epoxide hydrolase 2 | Enzyme |
| 33 | P67775 | PPP2CA | protein phosphatase 2 catalytic subunit alpha | - |
| 34 | O60674 | JAK2 | Janus kinase 2 | Kinase |
| 35 | O60885 | BRD4 | bromodomain containing 4 | Epigenetic regulator |
| 36 | Q02750 | MAP2K1 | mitogen-activated protein kinase kinase 1 | Kinase |
| 37 | P10275 | AR | androgen receptor | Nuclear receptor |
| 38 | P11362 | FGFR1 | fibroblast growth factor receptor 1 | Kinase |
| 39 | Q00535 | CDK5 | cyclin-dependent kinase 5 | Kinase |
| 40 | P03956 | MMP1 | matrix metallopeptidase 1 | Enzyme |
| 41 | P08254 | MMP3 | matrix metallopeptidase 3 | Enzyme |
| 42 | Q96RI1 | NR1H4 | nuclear receptor subfamily 1 group H member 4 | Nuclear receptor |
| 43 | Q99572 | P2RX7 | purinergic receptor P2X 7 | Ion channel |
| 44 | P49356 | FNTB | farnesyltransferase, CAAX box, beta | - |
| 45 | P30536 | TSPO | translocator protein | - |
| 46 | P23458 | JAK1 | Janus kinase 1 | Kinase |
| 47 | O75874 | IDH1 | isocitrate dehydrogenase (NADP(+)) 1 | - |
| 48 | P06239 | LCK | LCK proto-oncogene, SRC family tyrosine kinase | Kinase |
| 49 | P07333 | CSF1R | colony-stimulating factor 1 receptor | Kinase |
| 50 | Q13627 | DYRK1A | dual-specificity tyrosine phosphorylation regulated kinase 1A | Kinase |
| 51 | P07384 | CAPN1 | calpain 1 | Enzyme |
| 52 | Q9UDY8 | MALT1 | MALT1 paracaspase | Enzyme |
| 53 | P48039 | MTNR1A | melatonin receptor 1A | G-protein-coupled receptor |
| 54 | P49354 | FNTA | farnesyltransferase, CAAX box, alpha | Enzyme |
| 55 | Q12884 | FAP | fibroblast activation protein alpha | Enzyme |
| 56 | P18031 | PTPN1 | protein tyrosine phosphatase non-receptor type 1 | - |
| 57 | Q15078 | CDK5R1 | cyclin-dependent kinase 5 regulatory subunit 1 | Enzyme modulator |
| 58 | P25774 | CTSS | cathepsin S | Enzyme |
| 59 | P22001 | KCNA3 | potassium voltage-gated channel subfamily A member 3 | Ion channel |
| No. | UniProt ID | Gene | Degree | Relevance Score | Betweenness Centrality | Closeness Centrality |
|---|---|---|---|---|---|---|
| 1 | P12931 | SRC | 18 | 1.557 | 0.216 | 0.800 |
| 2 | P00519 | ABL1 | 12 | 0.882 | 0.085 | 0.667 |
| 3 | P06493 | CDK1 | 11 | 1.528 | 0.034 | 0.649 |
| 4 | P24385 | CCND1 | 11 | 2.873 | 0.024 | 0.649 |
| 5 | P24941 | CDK2 | 10 | 1.517 | 0.037 | 0.600 |
| 6 | Q00534 | CDK6 | 10 | 0.748 | 0.018 | 0.632 |
| 7 | P11802 | CDK4 | 10 | 2.379 | 0.022 | 0.632 |
| 8 | Q00535 | CDK5 | 10 | 0.196 | 0.039 | 0.615 |
| 9 | O60674 | JAK2 | 10 | 0.259 | 0.046 | 0.615 |
| 10 | P20248 | CCNA2 | 9 | 0.392 | 0.016 | 0.571 |
| 11 | Q02750 | MAP2K1 | 9 | 0.240 | 0.058 | 0.600 |
| 12 | P06239 | LCK | 9 | 0.139 | 0.027 | 0.615 |
| 13 | P45983 | MAPK8 | 8 | 1.536 | 0.026 | 0.600 |
| 14 | P09619 | PDGFRB | 7 | 0.868 | 0.009 | 0.571 |
| 15 | Q07817 | BCL2L1 | 7 | 2.128 | 0.018 | 0.585 |
| 16 | P18031 | PTPN1 | 6 | 0.098 | 0.012 | 0.571 |
| 17 | Q05397 | PTK2 | 6 | 0.854 | 0.014 | 0.545 |
| 18 | P23458 | JAK1 | 6 | 0.139 | 0.009 | 0.522 |
| 19 | P67775 | PPP2CA | 6 | 0.277 | 0.016 | 0.571 |
| 20 | P10275 | AR | 5 | 0.240 | 0.003 | 0.533 |
| 21 | P07384 | CAPN1 | 5 | 0.139 | 0.021 | 0.545 |
| 22 | O14746 | TERT | 5 | 0.439 | 0.011 | 0.500 |
| 23 | Q15078 | CDK5R1 | 4 | 0.098 | 0.005 | 0.500 |
| 24 | P17252 | PRKCA | 4 | 1.007 | 0.011 | 0.511 |
| 25 | P53779 | MAPK10 | 4 | 0.854 | 0.001 | 0.522 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Kwak, H.J.; Kim, B.H.; Kim, D.-W.; Kim, H.Y.; Kim, S.H.; Kang, K.S. Brevilin A Isolated from Centipeda minima Induces Apoptosis in Human Gastric Cancer Cells via an Extrinsic Apoptotic Signaling Pathway. Plants 2022, 11, 1658. https://doi.org/10.3390/plants11131658
Lee D, Kwak HJ, Kim BH, Kim D-W, Kim HY, Kim SH, Kang KS. Brevilin A Isolated from Centipeda minima Induces Apoptosis in Human Gastric Cancer Cells via an Extrinsic Apoptotic Signaling Pathway. Plants. 2022; 11(13):1658. https://doi.org/10.3390/plants11131658
Chicago/Turabian StyleLee, Dahae, Hee Jae Kwak, Byoung Ha Kim, Dong-Wook Kim, Hyun Young Kim, Seung Hyun Kim, and Ki Sung Kang. 2022. "Brevilin A Isolated from Centipeda minima Induces Apoptosis in Human Gastric Cancer Cells via an Extrinsic Apoptotic Signaling Pathway" Plants 11, no. 13: 1658. https://doi.org/10.3390/plants11131658
APA StyleLee, D., Kwak, H. J., Kim, B. H., Kim, D.-W., Kim, H. Y., Kim, S. H., & Kang, K. S. (2022). Brevilin A Isolated from Centipeda minima Induces Apoptosis in Human Gastric Cancer Cells via an Extrinsic Apoptotic Signaling Pathway. Plants, 11(13), 1658. https://doi.org/10.3390/plants11131658