
Identification of Quantitative Trait Loci for Leaf Rust and Stem Rust Seedling Resistance in Bread Wheat Using a Genome-Wide Association Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Screening of Infection Type of Pt and Pgt Races
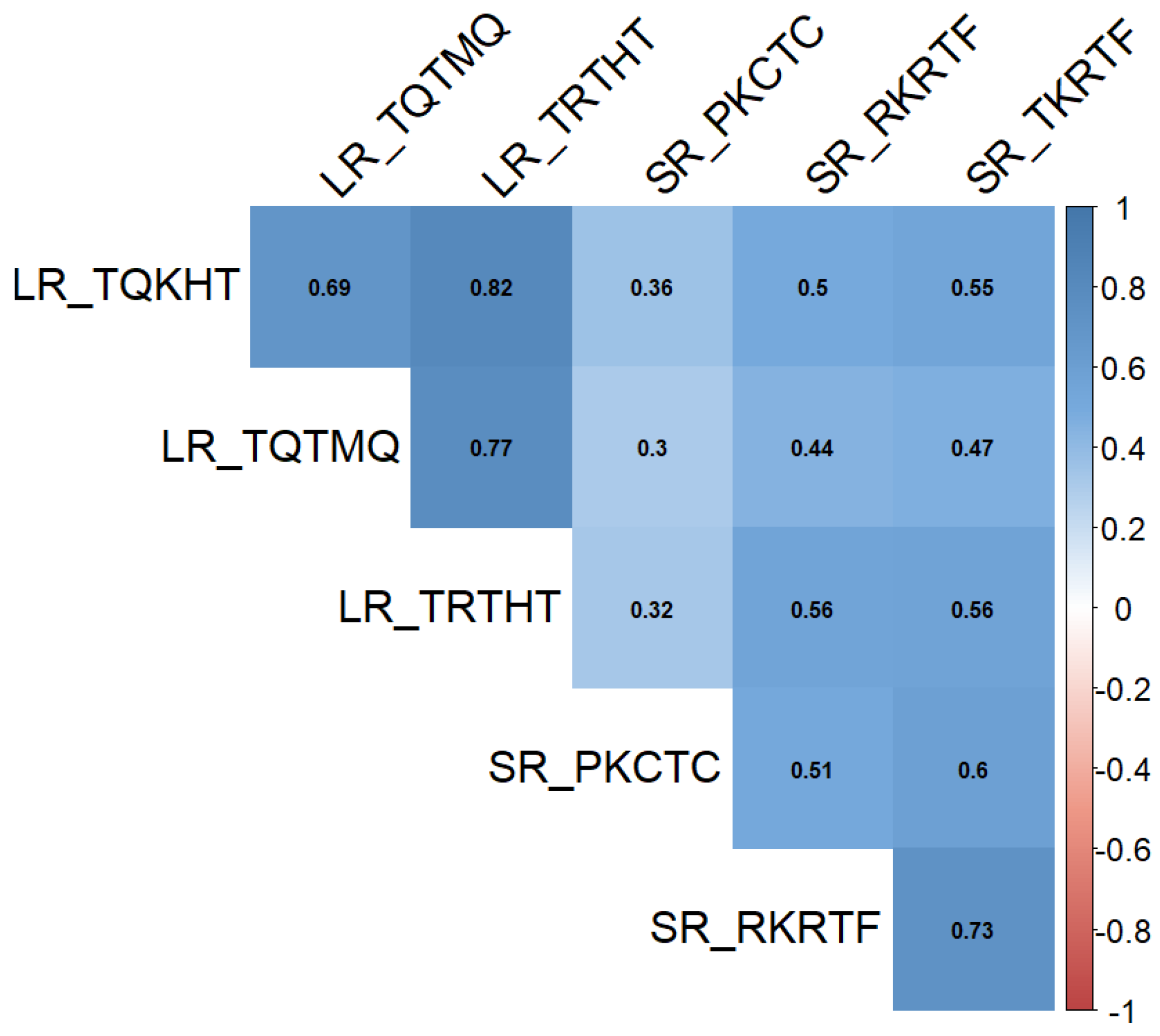
2.2. Genotyping Results and Analysis of the Population Structure
2.3. Association Mapping
3. Discussion
3.1. Patterns of Identified QTLs for Leaf and Stem Rust Resistances
3.2. Comparison of the Physical Positions of SNPs in Quantitative Trait Loci and Protein-Coding Genes
4. Materials and Methods
4.1. Genetic Material
4.2. Seedling LR and SR Evaluation
4.3. DNA Extracting and Genotyping
4.4. Association Mapping
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization Corporate Statistical Database (FAOSTAT). Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 20 August 2021).
- Statistics Committee. Ministry of National Economy of the Republic of Kazakhstan. Available online: https://stat.gov.kz/ (accessed on 13 September 2021).
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: Mexico City, Mexico, 1992; p. 81. [Google Scholar]
- Kolmer, J. Leaf rust of wheat: Pathogen biology, variation and host resistance. Forests 2013, 4, 70–84. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.P.; Hodson, D.P.; Jin, Y.; Lagudah, E.S.; Ayliffe, M.A.; Bhavani, S.; Rouse, M.N.; Pretorius, Z.A.; Szabo, L.J.; Huerta-Espino, J.; et al. Emergence and spread of new races of wheat stem rust fungus: Continued threat to food security and prospects of genetic control. Phytopathology 2015, 10, 872–884. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Espino, J.; Singh, R.; German, S.; Mccallum, B.; Park, R.; Chen, W.Q.; Bhardwaj, S.C.; Goyeau, H. Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 2011, 179, 143–160. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Bhavani, S.; Njau, P.; Herrera-Foessel, S.; Singh, P.K.; Singh, S.; Velu, G. The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Annu. Rev. Phytopathol. 2011, 49, 465–481. [Google Scholar] [CrossRef] [Green Version]
- Figlan, S.; Ntushelo, K.; Mwadzingeni, L.; Terefe, T.; Tsilo, T.J.; Shimelis, H. Breeding Wheat for Durable Leaf Rust Resistance in Southern Africa: Variability; Distribution; Current Control Strategies; Challenges and Future Prospects. Front. Plant Sci. 2020, 11, 549. [Google Scholar] [CrossRef] [PubMed]
- Samborski, D.J. Wheat leaf rust. In The Cereal Rusts Vol. II: Diseases, Distribution, Epidemiology and Control; Roelfs, A.P., Bushnell, W.R., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 39–59. [Google Scholar]
- Morgounov, A.; Rosseeva, L.; Koyshibayev, M. Leaf rust of spring wheat in Northern Kazakhstan and Siberia: Incidence, virulence, and breeding for resistance. Aust. J. Agric. Res. 2007, 58, 847. [Google Scholar] [CrossRef]
- Koyshybaev, M. Wheat Diseases, 1st ed.; FAO: Ankara, Turkey, 2018; p. 365. [Google Scholar]
- Shamanin, V.; Salina, E.; Wanyera, R.; Zelenskiy, Y.; Olivera, P.; Morgounov, A.I. Genetic diversity of spring wheat from Kazakhstan and Russia for resistance to stem rust Ug99. Euphytica 2016, 212, 287–296. [Google Scholar] [CrossRef]
- Rsaliyev, A.S.; Rsaliyev, S.S. Principal approaches and achievements in studying race composition of wheat stem rust. Vavilov J. Genet. Breed. 2018, 22, 967–977. [Google Scholar] [CrossRef]
- Shamanin, V.P.; Pototskaya, I.V.; Shepelev, S.S.; Pozherukova, V.E.; Salina, E.A.; Skolotneva, E.S.; Morgounov, A.I. Stem rust in Western Siberia–race composition and effective resistance genes. Vavilov J. Genet. Breed. 2020, 24, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Rsaliyev, A.; Yskakova, G.; Maulenbay, A.; Zakarya, K.; Rsaliyev, S. Virulence and race structure of Puccinia graminis f. sp. tritici in Kazakhstan. Plant Prot. Sci. 2020, 56, 275–284. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.F.; Xia, X.C. Catalogue of Gene Symbols for Wheat: 2017 Supplement. Available online: https://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf (accessed on 23 September 2021).
- Ellis, J.G.; Lagudah, E.; Spielmeyer, W.; Dodds, P.N. The past, present and future of breeding rust resistant wheat. Front. Plant Sci. 2014, 5, 641. [Google Scholar] [CrossRef] [Green Version]
- Bajgain, P.; Rouse, M.; Bulli, P.; Bhavani, S.; Gordon, T.; Wanyera, R.; Njau, P.N.; Legesse, W.; Andersen, J.A.; Pumphrey, M.O. Association mapping of North American spring wheat breeding germplasm reveals loci conferring resistance to Ug99 and other African stem rust races. BMC Plant Biol. 2015, 15, 249. [Google Scholar] [CrossRef]
- Bariana, H.S.; Hayden, M.J.; Ahmed, N.U.; Bell, J.A.; Sharp, P.J.; McIntosh, R.A. Mapping of durable adult plant and seedling resistances to stripe rust and stem rust diseases in wheat. Aust. J. Agric. Res. 2001, 52, 1247. [Google Scholar] [CrossRef]
- Olivera, P.D.; Sikharulidze, Z.; Dumbadze, R.; Szabo, L.J.; Newcomb, M.; Natsarishvili, K.; Rouse, M.N.; Luster, D.G.; Jin, Y. Presence of a sexual population of Puccinia graminis f.sp. tritici in Georgia provides a hotspot for genotypic and phenotypic diversity. Phytopathology 2019, 109, 2152–2160. [Google Scholar] [CrossRef] [Green Version]
- Li, T.Y.; Ma, Y.C.; Wu, X.X.; Chen, S.; Xu, X.F.; Wang, H.; Cao, Y.Y.; Xuan, Y.H. Race and virulence characterization of Puccinia graminis f. sp. tritici in China. PLoS ONE 2018, 13, e0197579. [Google Scholar] [CrossRef] [PubMed]
- Omara, R.I.; Nehela, Y.; Mabrouk, O.I.; Elsharkawy, M.M. The Emergence of New Aggressive Leaf Rust Races with the Potential to Supplant the Resistance of Wheat Cultivars. Biology 2021, 10, 925. [Google Scholar] [CrossRef]
- Admass, B.; Friedt, W.; Ordon, F. Stem rust seedling resistance genes in Ethiopian wheat cultivars and breeding lines. Afr. Crop Sci. J. 2012, 20, 149–162. [Google Scholar]
- Zhang, P.P.; Gebrewahid, T.W.; Yue, Z.; Li, Q.L.; Li, Z.F.; Liu, D.Q. Seedling and adult plant resistance to leaf rust in 46 Chinese bread wheat landraces and 39 wheat lines with known Lr genes. J. Integr. Agric. 2019, 18, 1014–1023. [Google Scholar] [CrossRef]
- Atia, M.A.; El-Khateeb, E.A.; El-Maksoud, A.; Reem, M.; Abou-Zeid, M.A.; Salah, A.; Abdel-Hamid, A.M. Mining of Leaf Rust Resistance Genes Content in Egyptian Bread Wheat Collection. Plants 2021, 10, 1378. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.G.; Friesen, T.L.; Xu, S.S.; Faris, J.D.; Kolmer, J.A. Identification of novel QTLs for seedling and adult plant leaf rust resistance in a wheat doubled haploid population. Theor. Appl. Genet. 2009, 119, 263–269. [Google Scholar] [CrossRef]
- Li, G.; Xu, X.; Bai, G.; Carver, B.F.; Hunger, R.; Bonman, J.M.; Kolmer, J.; Dong, H. Genome-wide association mapping reveals novel QTL for seedling leaf rust resistance in a worldwide collection of winter wheat. Plant Genome 2016, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Rollar, S.; Serfling, A.; Geyer, M.; Hartl, L.; Mohler, V.; Ordon, F. QTL mapping of adult plant and seedling resistance to leaf rust (Puccinia triticina Eriks.) in a multiparent advanced generation intercross (MAGIC) wheat population. Theor. Appl. Genet. 2021, 134, 37–51. [Google Scholar] [CrossRef]
- Afzal, A.; Ali, S.R.; Ijaz, M.; Saeed, M. Combating Ug99-Current Scenario. Int. J. Phytopathol. 2021, 10, 57–70. [Google Scholar] [CrossRef]
- Maulenbay, A.D.; Yskakova, G.S.; Rsaliyev, A. Virulence and racial composition of Puccina triticina in Kazakhstan in 2018. Her. Sci. S. Seifullin Kazakh Agro Tech. Univ. 2020, 3, 25–35. (In Russian) [Google Scholar]
- Genievskaya, Y.; Abugalieva, S.; Rsaliyev, A.; Yskakova, G.; Turuspekov, Y. QTL Mapping for Seedling and Adult Plant Resistance to Leaf and Stem Rusts in Pamyati Azieva à Paragon Mapping Population of Bread Wheat. Agronomy 2020, 10, 1285. [Google Scholar] [CrossRef]
- Semagn, K.; Bjørnstad, Å.; Ndjiondjop, M.N. Principles; requirements and prospects of genetic mapping in plants. Afr. J. Biotechnol. 2006, 5, 2569–2587. [Google Scholar]
- Genievskaya, Y.; Turuspekov, Y.; Rsaliyev, A.; Abugalieva, S. Genome-wide association mapping for resistance to leaf; stem; and yellow rusts of common wheat under field conditions of South Kazakhstan. PeerJ 2020, 8, e9820. [Google Scholar] [CrossRef]
- Amalova, A.; Abugalieva, S.; Babkenov, A.; Babkenova, S.; Turuspekov, Y. Genome-wide association study of yield components in spring wheat collection harvested under two water regimes in Northern Kazakhstan. PeerJ 2021, 9, e11857. [Google Scholar] [CrossRef]
- Dyck, P.L.; Samborski, D.J.; Anderson, R.G. Inheritance of adult-plant leaf rust resistance derived from the common wheat varieties Exchange and Frontana. Can. J. Genet. Cytol. 1966, 8, 665–671. [Google Scholar] [CrossRef]
- Singh, R.P.; McIntosh, R.A. Complementary genes for reaction to Puccinia recondita tritici in Triticum aestivum. II. Cytogenetic studies. Can. J. Genet. Cytol. 1984, 26, 736–742. [Google Scholar] [CrossRef]
- Bansal, U.K.; Hayden, M.J.; Venkata, B.P.; Khanna, R.; Saini, R.G.; Bariana, H.S. Genetic mapping of adult plant leaf rust resistance genes Lr48 and Lr49 in common wheat. Theor. Appl. Genet. 2008, 117, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Juliana, P.; Singh, R.P.; Singh, P.K.; Poland, J.A.; Bergstrom, G.C.; Huerta-Espino, J.; Bhavani, S.; Crossa, J.; Sorrells, M.E. Genome-wide association mapping for resistance to leaf rust; stripe rust and tan spot in wheat reveals potential candidate genes. Theor. Appl. Genet. 2018, 131, 1405–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Turner, M.K.; Chao, S.; Kolmer, J.; Anderson, J.A. Genome Wide Association Study of Seedling and Adult Plant Leaf Rust Resistance in Elite Spring Wheat Breeding Lines. PLoS ONE 2016, 11, e0148671. [Google Scholar] [CrossRef] [Green Version]
- Kertho, A.; Mamidi, S.; Bonman, J.M.; McClean, P.E.; Acevedo, M. Genome-Wide Association Mapping for Resistance to Leaf and Stripe Rust in Winter-Habit Hexaploid Wheat Landraces. PLoS ONE 2015, 10, e0129580. [Google Scholar] [CrossRef] [Green Version]
- Covarrubias-Pazaran, G.E. Optimizing Breeding Schemes. Manual. Heritability: Meaning and Computation, 1st ed.; CGIAR Excellence in Breeding Platform (EiB): Montpellier, France, 2019; p. 10. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation. Version II. Plant Mol. Biol. Report. 1983, 4, 19–21. [Google Scholar] [CrossRef]
- Saccomanno, A.; Matny, O.; Marone, D.; Laidò, G.; Petruzzino, G.; Mazzucotelli, E.; Desiderio, F.; Blanco, A.; Gadaleta, A.; Pecchioni, N.; et al. Genetic Mapping of Loci for Resistance to Stem Rust in a Tetraploid Wheat Collection. Int. J. Mol. Sci. 2018, 19, 3907. [Google Scholar] [CrossRef] [Green Version]
- Dakouri, A.; McCallum, B.D.; Radovanovic, N.; Cloutier, S. Molecular and phenotypic characterization of seedling and adult plant leaf rust resistance in a world wheat collection. Mol. Breed. 2013, 32, 663–677. [Google Scholar] [CrossRef] [Green Version]
- Aoun, M.; Kolmer, J.A.; Rouse, M.N.; Elias, E.M.; Breiland, M.; Bulbula, W.D.; Chao, S.; Acevedo, M. Mapping of Novel Leaf Rust and Stem Rust Resistance Genes in the Portuguese Durum Wheat Landrace PI 192051. Genes Genomes Genet. 2019, 9, 2535–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlin, M.H.; Andrews, J.; Toh, S.S. Essential Letters in the Fungal Alphabet: ABC and MFS Transporters and Their Roles in Survival and Pathogenicity. Adv. Genet. 2014, 85, 201–253. [Google Scholar] [CrossRef]
- Gou, J.Y.; Li, K.; Wu, K.; Wang, X.; Lin, H.; Cantu, D.; Uauy, C.; Dobon-Alonso, A.; Midorikawa, T.; Inoue, K.; et al. Wheat Stripe Rust Resistance Protein WKS1 Reduces the Ability of the Thylakoid-Associated Ascorbate Peroxidase to Detoxify Reactive Oxygen Species. Plant Cell 2015, 27, 1755–1770. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Wang, B.; Zhang, Q.; Fu, Y.; Huang, L.; Wang, X.; Kang, Z. Exploration of microRNAs and their targets engaging in the resistance interaction between wheat and stripe rust. Front. Plant Sci. 2015, 6, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowell, J.B. Controlled infection by Puccinia graminis f.sp. tritici under artificial conditions. In The Cereal Rusts, Vol 1: Origins, Specificity, Structure and Physiology; Bushnell, W.R., Roelfs, A.P., Eds.; Academic Press: Orlando, FL, USA, 1984; pp. 292–332. [Google Scholar]
- Spanic, V.; Rouse, M.N.; Kolmer, J.A.; Anderson, J.A. Leaf and stem seedling rust resistance in wheat cultivars grown in Croatia. Euphytica 2015, 203, 437–448. [Google Scholar] [CrossRef]
- Pretorius, Z.A.; Jin, Y.; Bender, C.M.; Herselman, L.; Prin, R. Seedling resistance to stem rust race Ug99 and marker analysis for Sr2, Sr24 and Sr31 in South African wheat cultivars and lines. Euphytica 2012, 186, 15–23. [Google Scholar] [CrossRef]
- Stakman, E.C.; Stewart, D.M.; Loegering, W.Q. Identification of physiologic races of Puccinia graminis var. tritici. US Agric. Res. Serv. 1962, 617, 1–53. [Google Scholar]
- Zhang, D.; Bowden, R.; Bai, G. A method to linearize Stakman infection type ratings for statistical analysis. In Proceedings of the Borlaug Global Rust Initiative 2011 Technical Workshop, Saint Paul, MN, USA, 13–16 June 2011. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Zhang, Z.; Ersoz, E.; Lai, C.; Todhunter, R.; Tiwari, H.; Gore, M.; Bradbury, P.; Yu, J.; Arnett, D.K.; Ordovas, J.M.; et al. Mixed linear model approach adapted for genome-wide association studies. Nat. Genet. 2010, 42, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Edae, E.A.; Bowden, R.L.; Poland, J. Application of population sequencing (POPSEQ) for ordering and imputing genotyping-by-sequencing markers in hexaploid wheat. G3 Genes Genomes Genet. 2015, 5, 2547–2553. [Google Scholar] [CrossRef] [Green Version]
- BLAST Tool. Triticum Aestivum. Available online: https://plants.ensembl.org/Triticum_aestivum/Tools/Blast (accessed on 4 October 2021).
- EnsemblPlant. Triticum Aestivum. Available online: https://plants.ensembl.org/Triticum_aestivum/Info/Index (accessed on 7 October 2021).
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
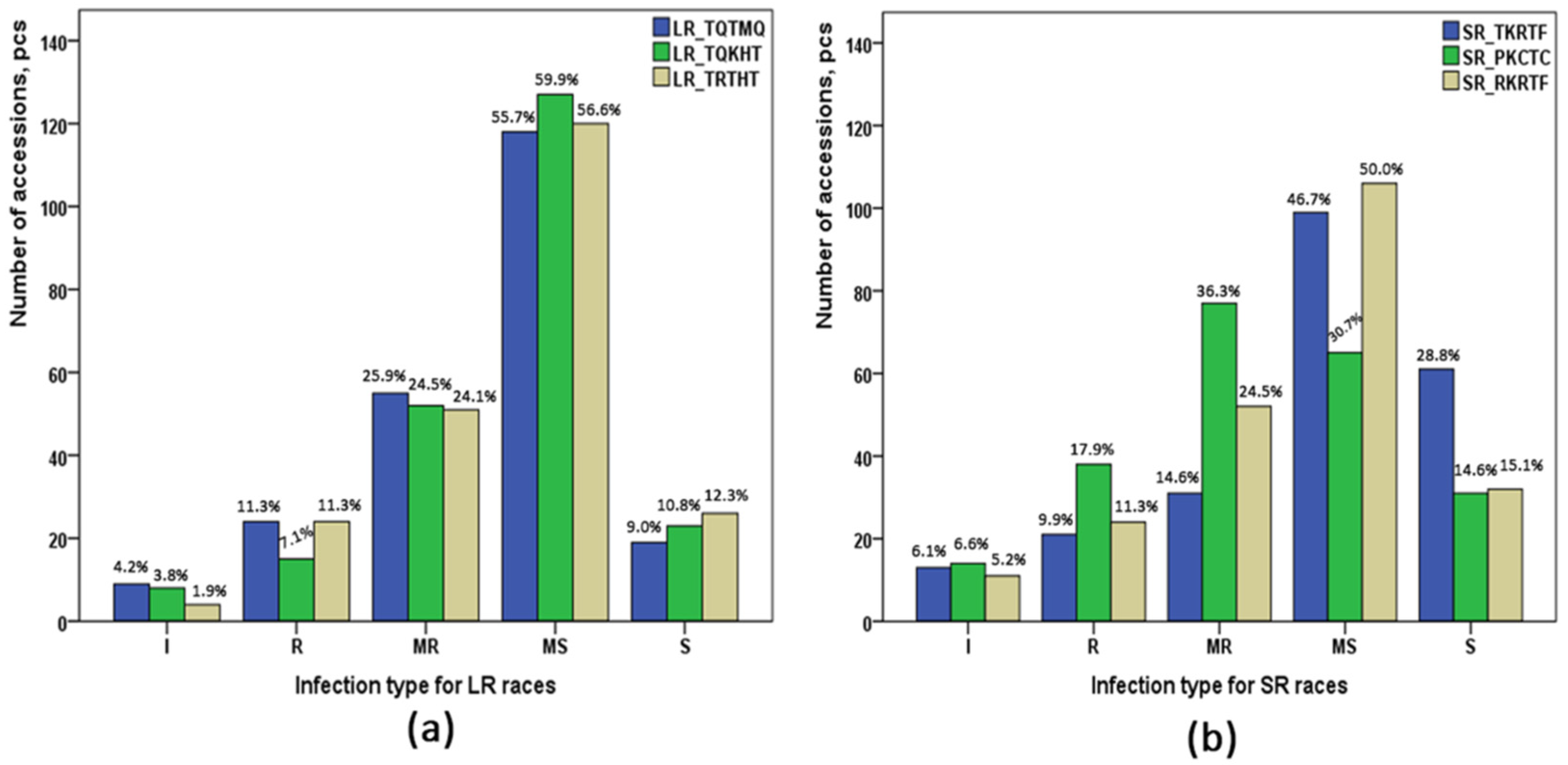

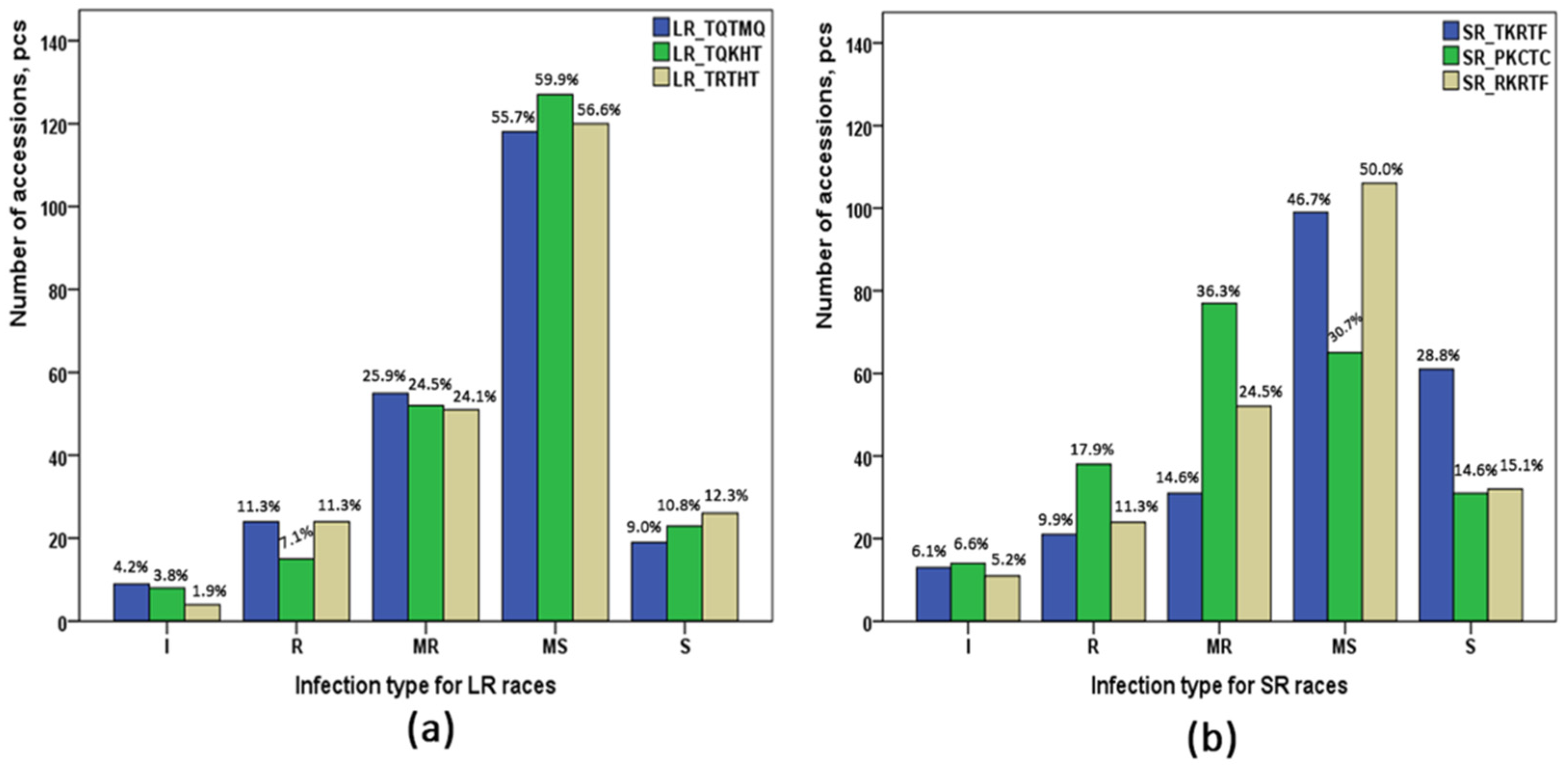{kind=link}
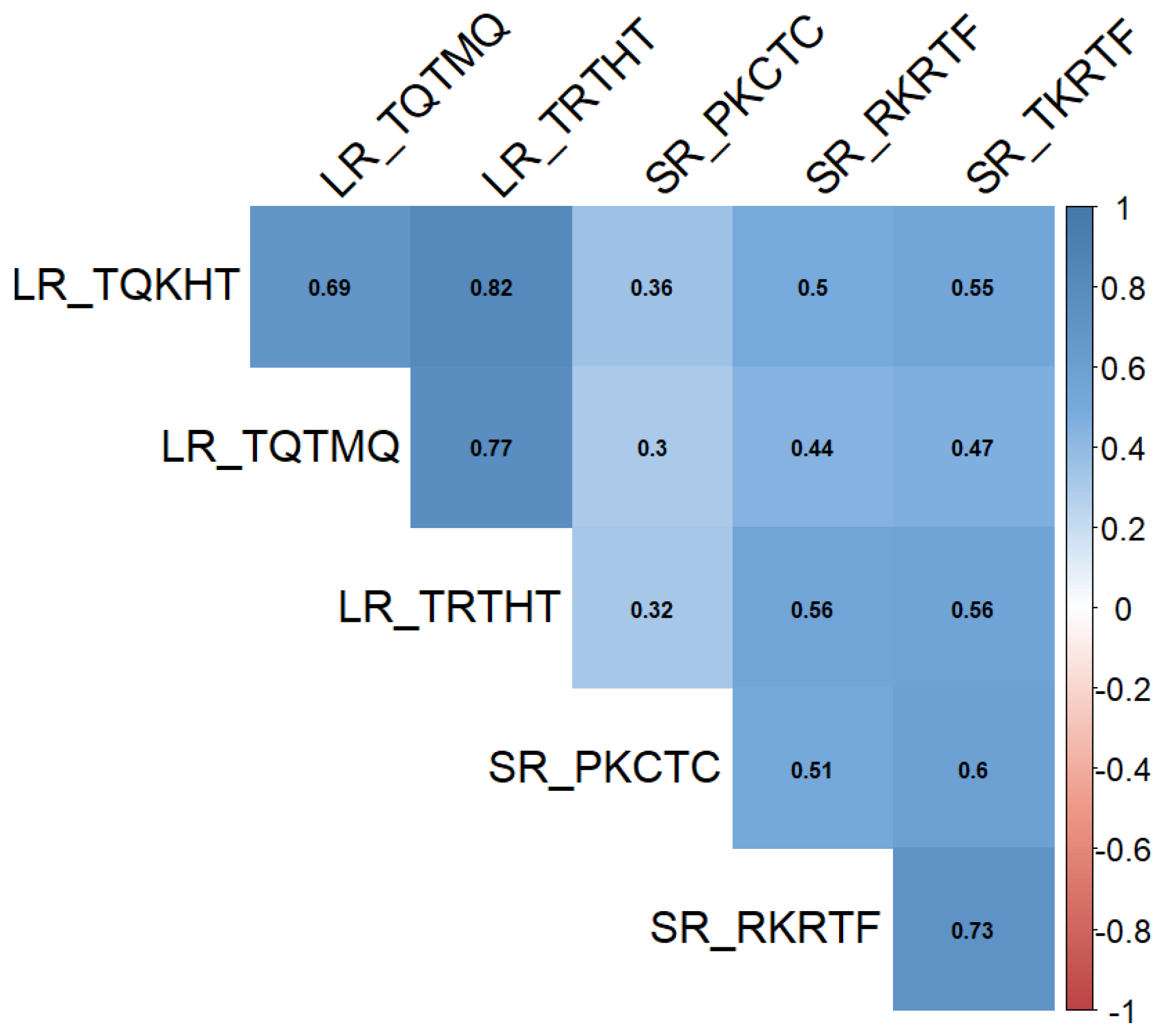{kind=link}
| Disease | Factor | df | SS | MS | F-Value | p-Value | Genotype (H2) % | Race % | Genotype × Race % |
|---|---|---|---|---|---|---|---|---|---|
| LR | Genotype | 211 | 5222 | 24.8 | 20.9 | <2 × 10−16 | 74.60 | 0.47 | 14.19 |
| Race | 2 | 33 | 16.7 | 14.1 | 9.97 × 10−07 | ||||
| Genotype × Race | 422 | 993 | 2.4 | 2.0 | 1.99 × 10−15 | ||||
| Residuals | 636 | 752 | 1.2 | ||||||
| SR | Genotype | 211 | 5852 | 27.7 | 16.9 | <2 × 10−16 | 63.35 | 3.18 | 22.14 |
| Race | 2 | 294 | 146.9 | 89.4 | <2 × 10−16 | ||||
| Genotype × Race | 422 | 2045 | 4.9 | 3.0 | <2 × 10−16 | ||||
| Residuals | 636 | 1046 | 1.6 |
| APR LR | APR SR | LR_TQTMQ | LR_TQKHT | LR_TRTHT | SR_TKRTF | SR_PKCTC | |
|---|---|---|---|---|---|---|---|
| APR SR | 0.5580 *** | ||||||
| LR_TQTMQ | 0.6037 *** | 0.4290 *** | |||||
| LR_TQKHT | 0.6549 *** | 0.4748 *** | 0.6936 *** | ||||
| LR_TRTHT | 0.6672 *** | 0.4994 *** | 0.7734 *** | 0.8155 *** | |||
| SR_TKRTF | 0.5563 *** | 0.6838 *** | 0.4687 *** | 0.5494 *** | 0.5633 *** | ||
| SR_PKCTC | 0.2702 *** | 0.5607 *** | 0.3002 *** | 0.3590 *** | 0.3228 *** | 0.5968 *** | |
| SR_RKRTF | 0.5483 *** | 0.6563 *** | 0.4415 *** | 0.5033 *** | 0.5566 *** | 0.7261 *** | 0.5115 *** |
| # | SNP Marker | Chr. | Pos1. (cM) | Pos2. (bp) | Leaf Rust (LR) | Stem Rust (SR) | Allele | Effect | R2 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TQTMQ | TQKHT | TRTHT | TKRTF | PKCTC | RKRTF | ||||||||
| 1 | Kukri_c41943_535 | 1A | 38.1 | 13,230,954 | - | - | 1.29 × 10−05 | - | - | - | A | 1.61 | 0.10 |
| 2 | TA001473-0980 | 1B | 62.4 | 54,075,692 | - | 1.32 × 10−05 | 1.37 × 10−05 | - | - | - | A | 2.41 | 0.09 |
| 3 | BS00078431_51 | 1B | 70.8 | 346,871,402 | 3.61 × 10−05 | 9.85 × 10−06 | 5.45 × 10−08 | 7.21 × 10−07 | - | - | C | 1.72 | 0.11 |
| 4 | BS00063511_51 | 1D | 167.1 | 485,708,706 | - | - | 2.46 × 10−05 | 5.33 × 10−05 | - | - | A | 1.46 | 0.08 |
| 5 | BobWhite_c14476_80 | 2A | 102 | 86,901,274 | 2.09 × 10−06 | 3.23 × 10−06 | 6.33 × 10−08 | 2.47 × 10−07 | - | 8.40 × 10−05 | A | −1.67 | 0.11 |
| 6 | Excalibur_c27349_166 | 4B | 77.9 | 619,448,536 | 9.81 × 10−08 | 1.07 × 10−05 | 2.43 × 10−11 | 1.59 × 10−05 | - | - | C | −1.93 | 0.15 |
| 7 | GENE-2307_1216 | 5B | 147.4 | 531,888,962 | - | - | 4.50 × 10−05 | - | - | - | G | 1.53 | 0.08 |
| 8 | wsnp_Ex_rep_c68175_66950387 | 6A | 31.9 | 13,600,765 | 1.67 × 10−05 | 4.37 × 10−05 | 8.37 × 10−07 | 1.87 × 10−05 | - | - | C | 1.70 | 0.09 |
| 9 | TA003021-1057 | 6A | 56.1 | 34,974,650 | 9.58 × 10−06 | 3.51 × 10−05 | 1.25 × 10−07 | 6.51 × 10−06 | - | - | A | −1.83 | 0.10 |
| 10 | BobWhite_c24063_231 | 7A | 127.7 | 232,746,015 | 1.46 × 10−06 | 4.24 × 10−05 | 3.42 × 10−07 | 4.83 × 10−07 | - | 7.16 × 10−05 | C | 1.82 | 0.10 |
| 11 | TA003458-0086 | 7A | 133.9 | 565,347,833 | - | - | 5.44 × 10−06 | 3.39 × 10−06 | - | - | C | 1.77 | 0.10 |
| # | SNP Marker | QTL ref | Candidate Gene(s) | Overlapping Gene(s) | Protein | Orthologue Gene(s) | Identity (%) |
|---|---|---|---|---|---|---|---|
| 1 | Kukri_c41943_535 | QLr.ipbb-1A.3 | - | TraesCS1A02G027800 | Uncharacterized protein | TRIUR3_10793 (T. urartu) | 97.8 |
| 2 | TA001473-0980 | QLr.ipbb-1B.5 | - | TraesCS1A02G071800 | Uncharacterized protein | AET1Gv20182900 (A. tauschii) | 99.0 |
| 3 | BS00078431_51 | QLr.ipbb-1B.2 *; QSr.ipbb-1B.2 *; QTKW.ta.ipbb-1D †, QNPS.ta.ipbb-1B † | - | - | - | - | - |
| 4 | BS00063511_51 | QLr.ipbb-1D.2 *; QSr.ipbb-1D.2; QTKW.ta.ipbb-1D †, QNPS.ta.ipbb-1B † | - | TraesCS1D02G439800 | Trimethylguanosine synthase | AET1Gv21018700 (A. tauschii) | 96.3 |
| 5 | BobWhite_c14476_80 | QLr.ipbb-2A.2 *; QSr.ipbb-2A.3 | - | TraesCS2A02G141400 | Cation efflux protein | TRITD_2Av1G039940 (T. turgidum) | 99.0 |
| 6 | Excalibur_c27349_166 | QLr.ipbb-4B.2 *; QSr.ipbb-4B.1; QTKW.ta.ipbb-4B † | Lr12 [35], Lr31 [36], Lr49 [37] | TraesCS4B02G328500 | MFS domain-containing protein | TRITD_4Bv1G186380 (T. turgidum) | 99.7 |
| 7 | GENE-2307_1216 | QLr.ipbb-5B.1 * | - | TraesCS5D02G505900 | Uncharacterized protein | AET5Gv21130000 (A. tauschii) | 100 |
| 8 | wsnp_Ex_rep_c68175_66950387 | QLr.ipbb-6A.6 *; QSr.ipbb-6A.3 | - | TraesCS6D02G032300 | Protein kinase domain-containing protein | AET6Gv20067300 (A. tauschii) | 99.9 |
| 9 | TA003021-1057 | QLr.ipbb-6A.2 *; QSr.ipbb-6A.1 *; QTKW.ta.ipbb-6A †, QNPS.ta.ipbb-6A † | - | - | - | - | - |
| 10 | BobWhite_c24063_231 | QLr.ipbb-7A.2 *; QSr.ipbb-7A.2 | - | TraesCS7A02G250500 | L-ascorbate peroxidase 6 | TRITD_7Av1G095170 (T. turgidum) | 99.6 |
| 11 | TA003458-0086 | QLr.ipbb-7A.2 *; QSr.ipbb-7A.3; QNPS.ta.ipbb-7A †, QTKW.ta.ipbb-7A † | - | TraesCS7A02G389100 | Rab-GAP TBC domain-containing protein | TRITD7Av1G209600 (T. turgidum) | 100.0 |
| Disease (Pathogen) | Race | Avirulent (Effective) Genes | Virulent (Ineffective) Genes |
|---|---|---|---|
| LR (Puccinia triticina Eriks.) | TQTMQ | Lr24, 26, 20, 25, 14a, 15 | Lr1, 2a, 2c, 3, 9, 16, 3ka, 11, 17, 30, 19, 29, 2b, 3bg |
| TQKHT | Lr24, 26, 3ka, 19, 25 | Lr1, 2a, 2c, 3, 9, 16, 11, 17, 30, 20, 29, 2b, 3bg, 14a, 15 | |
| TRTHT | Lr24, 19, 25 | Lr1, 2a, 2c, 3, 9, 16, 26, 3ka, 11, 17, 30, 20, 29, 2b, 3bg, 14a, 15 | |
| SR (Puccinia graminis Pers. f. sp. tritici Eriks. & E. Henn.) | TKRTF | Sr11, 30, 24, 31 | Sr5, 21, 9e, 7b, 6, 8a, 9g, 36, 9b, 17, 9a, 9d, 10, 38, Tmp, McN |
| PKCTC | Sr21, 11, 36, 9b, 30, 24, 31, 38 | Sr5, 9e, 7b, 6, 8a, 9g, 17, 9a, 9d, 10, Tmp, McN | |
| RKRTF | Sr9e, 11, 30, 24, 31 | Sr5, 21, 7b, 6, 8a, 9g, 36, 9b, 17, 9a, 9d, 10, 38, Tmp, McN |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zatybekov, A.; Genievskaya, Y.; Rsaliyev, A.; Maulenbay, A.; Yskakova, G.; Savin, T.; Turuspekov, Y.; Abugalieva, S. Identification of Quantitative Trait Loci for Leaf Rust and Stem Rust Seedling Resistance in Bread Wheat Using a Genome-Wide Association Study. Plants 2022, 11, 74. https://doi.org/10.3390/plants11010074
Zatybekov A, Genievskaya Y, Rsaliyev A, Maulenbay A, Yskakova G, Savin T, Turuspekov Y, Abugalieva S. Identification of Quantitative Trait Loci for Leaf Rust and Stem Rust Seedling Resistance in Bread Wheat Using a Genome-Wide Association Study. Plants. 2022; 11(1):74. https://doi.org/10.3390/plants11010074
Chicago/Turabian StyleZatybekov, Alibek, Yuliya Genievskaya, Aralbek Rsaliyev, Akerke Maulenbay, Gulbahar Yskakova, Timur Savin, Yerlan Turuspekov, and Saule Abugalieva. 2022. "Identification of Quantitative Trait Loci for Leaf Rust and Stem Rust Seedling Resistance in Bread Wheat Using a Genome-Wide Association Study" Plants 11, no. 1: 74. https://doi.org/10.3390/plants11010074
APA StyleZatybekov, A., Genievskaya, Y., Rsaliyev, A., Maulenbay, A., Yskakova, G., Savin, T., Turuspekov, Y., & Abugalieva, S. (2022). Identification of Quantitative Trait Loci for Leaf Rust and Stem Rust Seedling Resistance in Bread Wheat Using a Genome-Wide Association Study. Plants, 11(1), 74. https://doi.org/10.3390/plants11010074