
Influence of Seed Source and Soil Contamination on Ecophysiological Responses of Lavandula pedunculata in Rehabilitation of Mining Areas




Abstract
:1. Introduction
2. Results
2.1. Soil Characterization
2.2. Germination and Growth of L. pedunculata
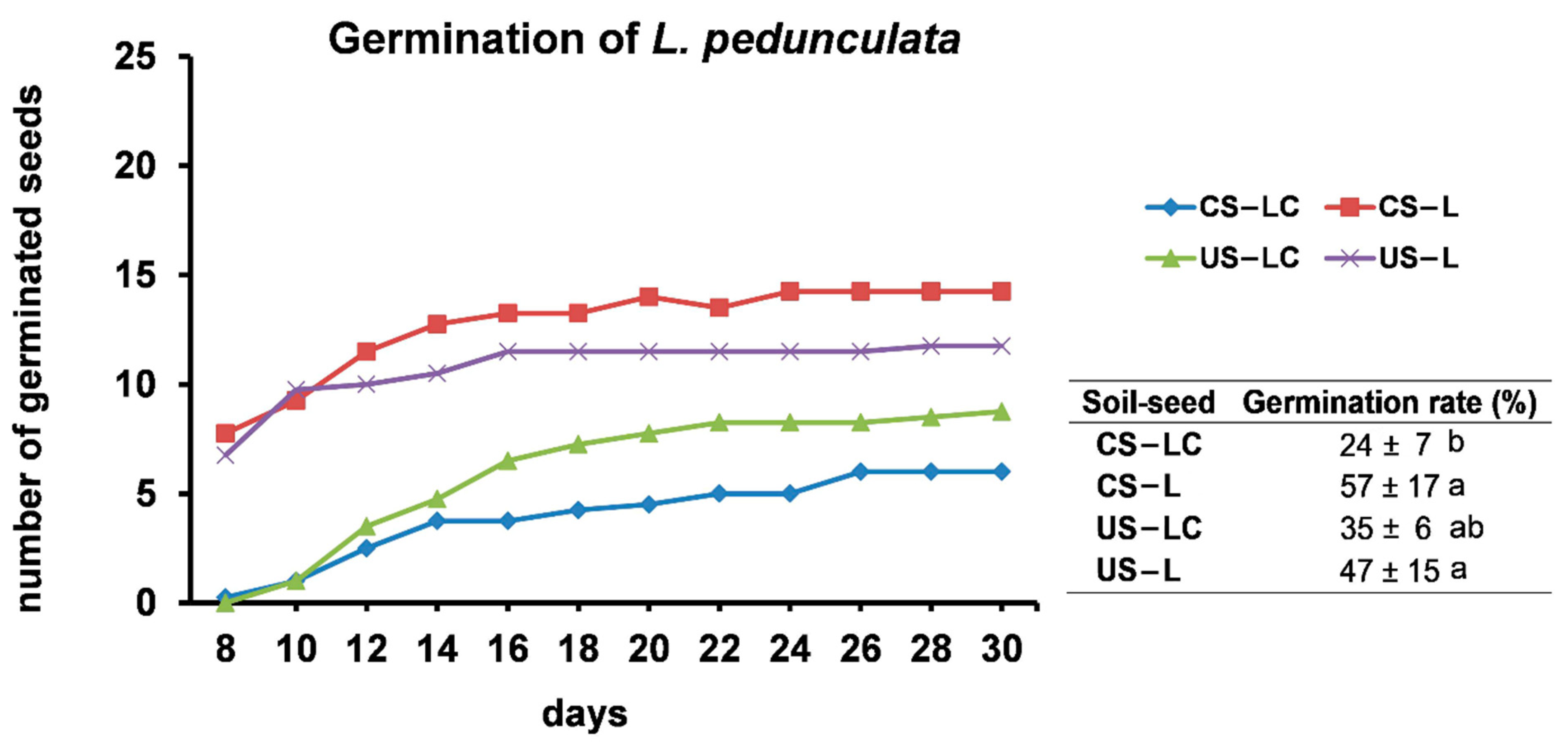
2.2.1. Germination Assays
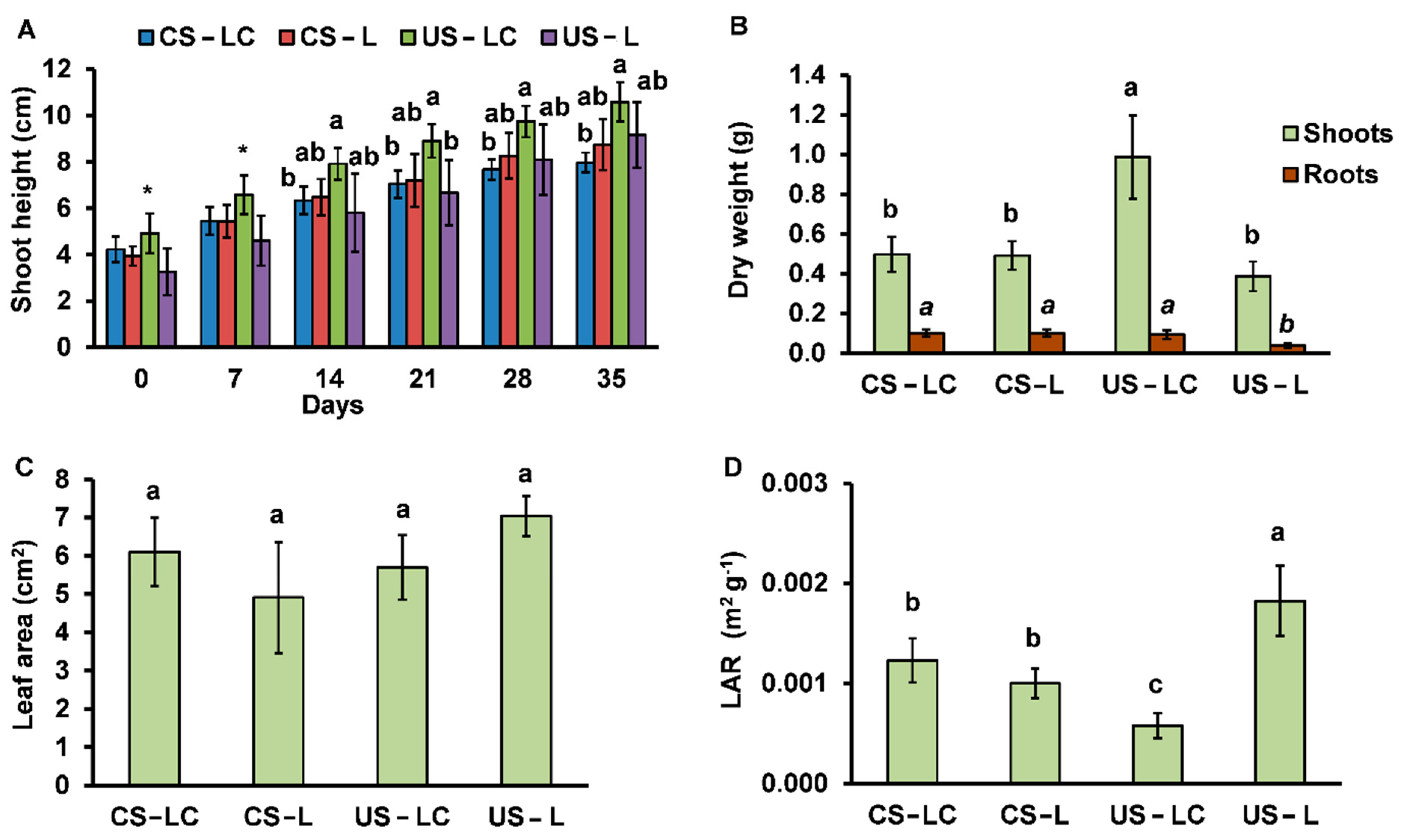
2.2.2. Plant Development
2.3. Metal(loid)s Concentration in Roots and Shoots
2.4. Physiologcal Parameters
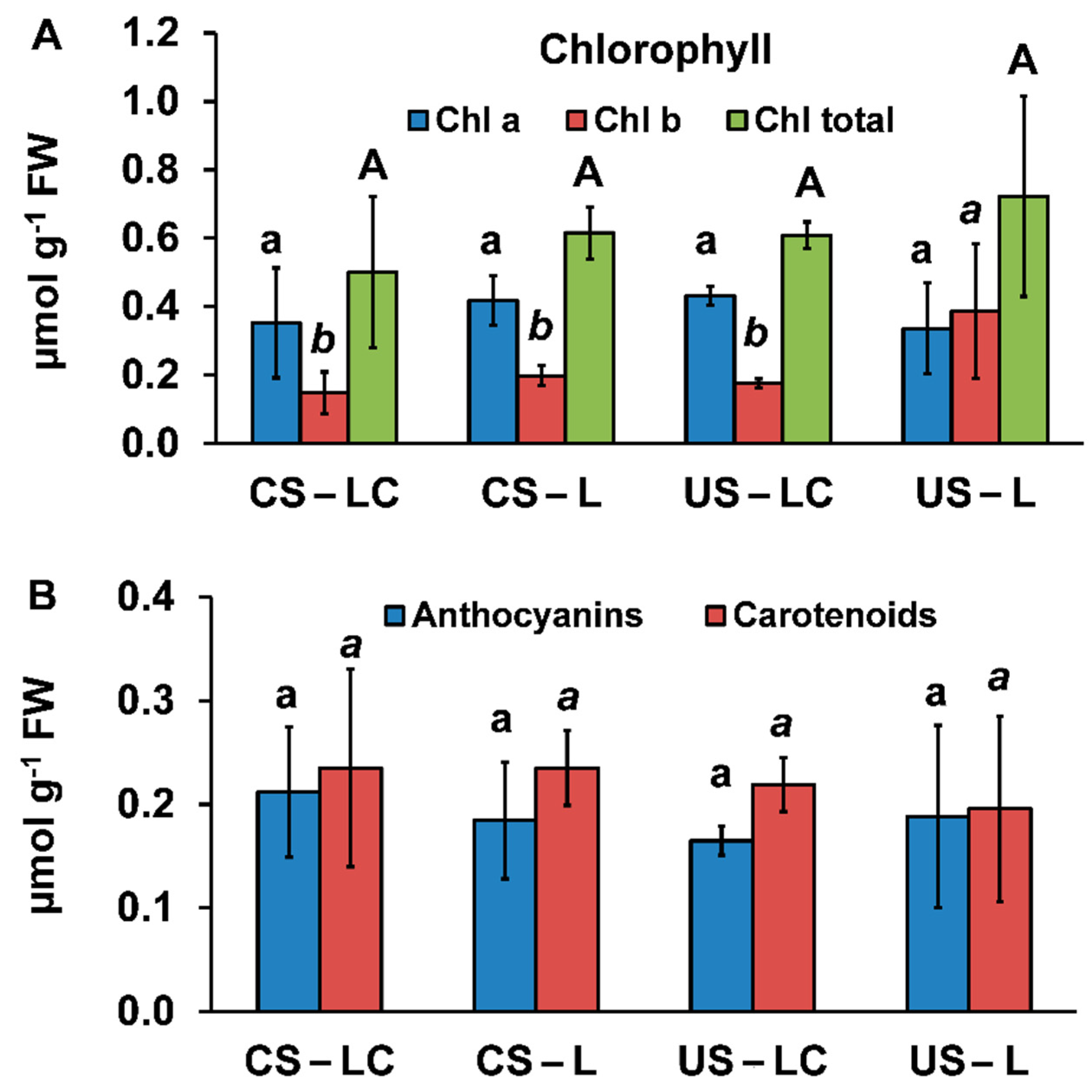
2.4.1. Pigment Concentrations and Visual Symptoms
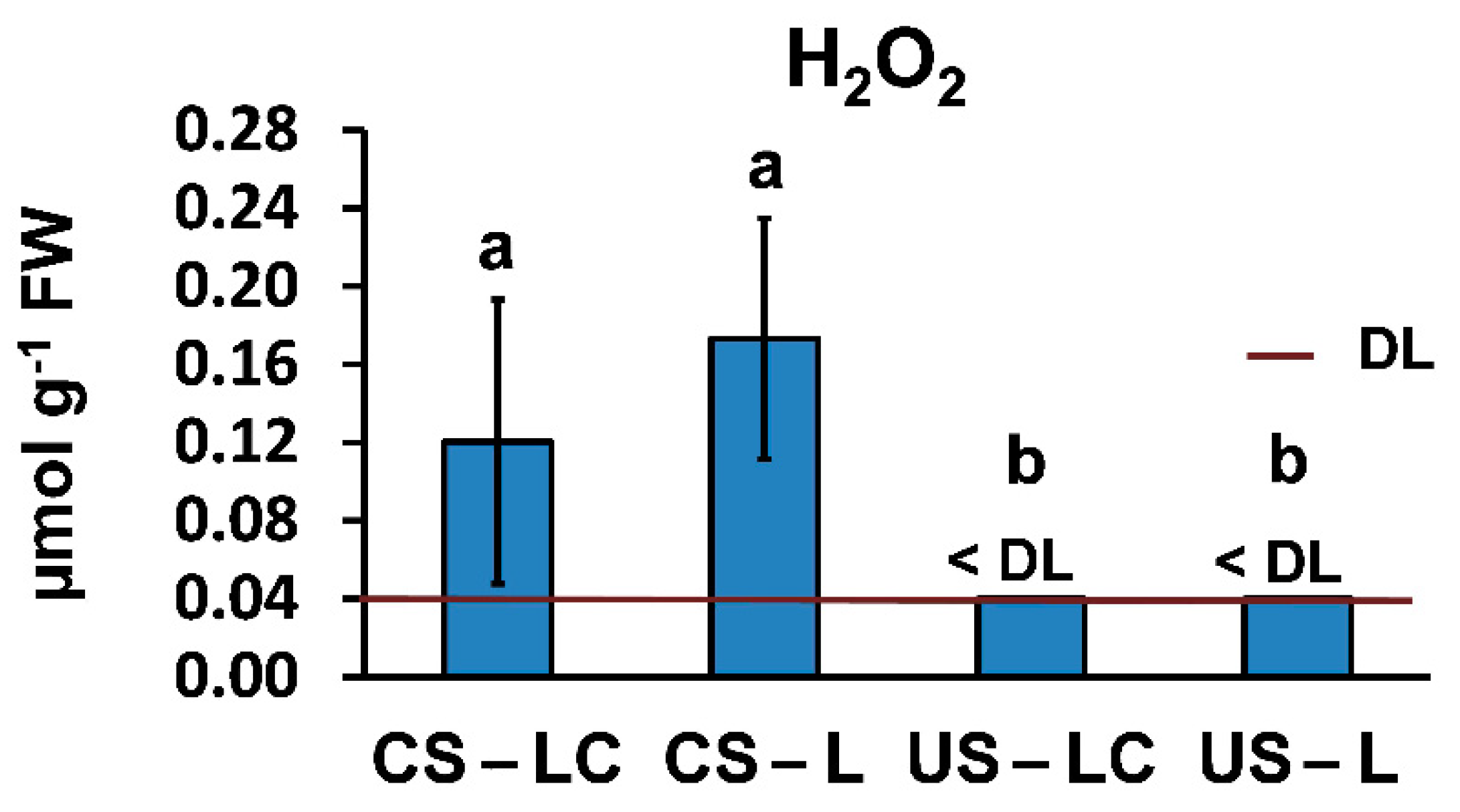
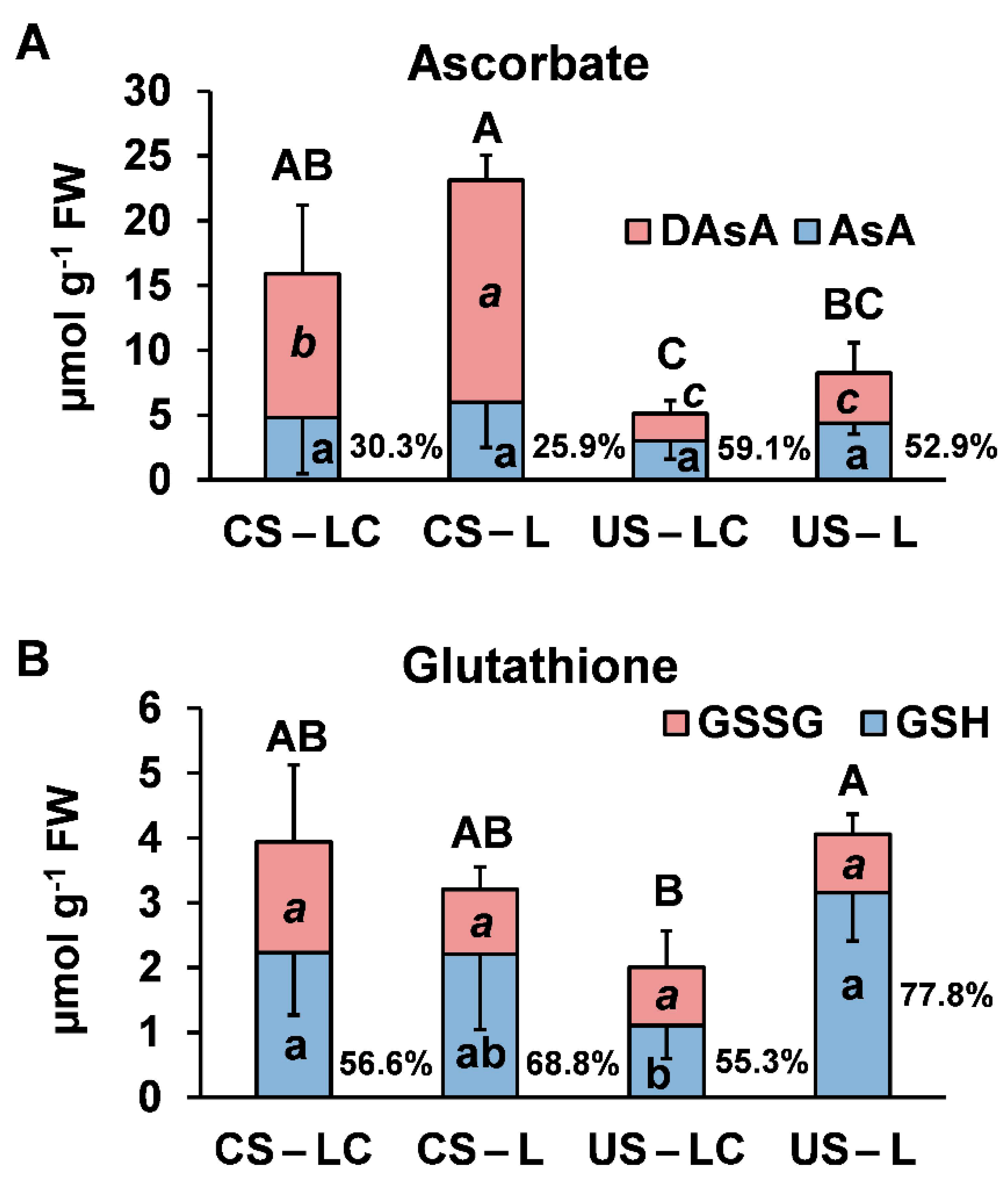
2.4.2. Oxidative Stress and Physiological Response
3. Discussion
4. Material and Methods
4.1. Study Areas and Material Sampling
4.2. Experimental Set-Up and Monitoring
4.2.1. Germination Assay
4.2.2. Plant Development and Ecophysiological Assay
4.3. Analytical Methods
4.3.1. Soil Characterization
4.3.2. Multielemental Concentration in Shoots and Roots of L. pedunculata and Plant Physiological Analysis
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Csavina, J.; Field, J.; Taylor, M.P.; Gao, S.; Landázuri, A.; Betterton, E.A.; Sáez, A.E. A review on the importance of metals and metalloids in atmospheric dust and aerosol from mining operations. Sci. Total Environ. 2012, 433, 58–73. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.S.; Abreu, M.M.; Saraiva, J.A. Mutielemental concentration and physiological responses of Lavandula pedunculata growing in soils developed on different mine wastes. Environ. Pollut. 2016, 213, 43–52. [Google Scholar] [CrossRef]
- Wakeel, A.; Xu, M.; Gan, Y. Chromium-induced reactive oxygen species accumulation by altering the enzymatic antioxidant system and associated cytotoxic, genotoxic, ultrastructural, and photosynthetic changes in plants. Int. J. Mol. Sci. 2020, 21, 728. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; He, T.; Xu, F.; Li, X.; Yuan, L.; Wang, Q.; Liu, H. Analysis of physiological and metabolite response of Celosia argentea to copper stress. Plant Biol. 2021, 23, 391–399. [Google Scholar] [CrossRef]
- Abreu, M.M.; Santos, E.S.; Magalhães, M.C.F.; Fernandes, E. Trace elements tolerance, accumulation and translocation in Cistus populifolius, Cistus salviifolius and their hybrid growing in polymetallic contaminated mine areas. J. Geochem. Explor. 2012, 123, 52–60. [Google Scholar] [CrossRef]
- Arenas-Lago, D.; Santos, E.S.; Carvalho, L.C.; Abreu, M.M.; Andrade, M.L. Cistus monspeliensis L. as a potential species for rehabilitation of soils with multielemental contamination under Mediterranean conditions. Environ. Sci. Pollut. Res. 2018, 25, 6443–6455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, E.S.; Abreu, M.M.; Nabais, C.; Saraiva, J.A. Trace elements and activity of antioxidative enzymes in Cistus ladanifer L. growing on an abandoned mine area. Ecotoxicology 2009, 18, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.S.; Abreu, M.M.; Batista, M.J.; Magalhães, M.C.F.; Fernandes, E. Inter-population variation on the accumulation and translocation of potentially harmful chemical elements in Cistus ladanifer L. from Brancanes, Caveira, Chança, Lousal, Neves Corvo and São Domingos mines in the Portuguese Iberian Pyrite Belt. J. Soils Sediments 2014, 14, 758–772. [Google Scholar] [CrossRef]
- Santos, E.S.; Abreu, M.M.; Nabais, C.; Magalhães, M.C.F. Trace elements distribution in soils developed on gossan mine wastes and Cistus ladanifer L. tolerance and bioaccumulation.s. Geochem. Explor. 2012, 123, 45–51. [Google Scholar] [CrossRef]
- Rossini-Oliva, S.; Mingorance, M.D.; Leidi, E.O.; Fernández-Espinosa, A.J. Does the polluted environment modify responses to metal pollution? A case study of two Cistus species and the excess of copper and lead. Catena 2019, 178, 244–255. [Google Scholar] [CrossRef]
- Abreu, M.M.; Batista, M.J.; Magalhães, M.C.F.; Matos, J.X. Acid mine drainage in the Portuguese Iberian Pyrite Belt. In Mine Drainage and Related Problems; Nova Science Publishers Inc.: New York, NY, USA, 2011. [Google Scholar]
- Anawar, H.M.; Freitas, M.C.; Canha, N.; Regina, I.S. Arsenic, antimony, and other trace element contamination in a mine tailings affected area and uptake by tolerant plant species. Environ. Geochem. Health 2011, 33, 353–362. [Google Scholar] [CrossRef]
- Márquez-García, B.; Pérez-López, R.; Ruíz-Chancho, M.J.; López-Sánchez, J.F.; Rubio, R.; Abreu, M.M.; Nieto, J.M.; Córdoba, F. Arsenic speciation in soils and Erica andevalensis Cabezudo & Rivera and Erica australis L. from São Domingos Mine area, Portugal. J. Geochem. Explor. 2012, 119-120, 51–59. [Google Scholar] [CrossRef]
- Delgado, F.; Gonçalves, O.; Amaro-Silva, C.; Silva, L.; Caldeira, R.; Castanheira, I.; Oliveira, R.; Alberto, D.; Jacinto, P.; Sousa, E.; et al. Seed germination and essential oil of Lavandula luisieri from Central Eastern Portugal. Acta Hortic. 2006, 723, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, A.M.; Peco, B. Interference between perennial grassland and Lavandula stoechas subsp. pedunculata seedlings: A case of spatial segregation cause by competition. Acta Oecol. 2004, 26, 39–44. [Google Scholar] [CrossRef]
- Anawar, H.M.; Canha, N.; Santa-Regina, I.; Freitas, M.C. Adaptation, tolerance, and evolution of plant species in a pyrite mine in response to contamination level and properties of mine tailings: Sustainable rehabilitation. J. Soils Sediments 2013, 13, 730–741. [Google Scholar] [CrossRef]
- Batista, M.J.; de Oliveira, D.P.S.; Abreu, M.M.; Locutura, J.; Shepherd, T.; Matos, J.; Bel-Lan, A.; Martins, L. Sources, background and enrichment of lead and other elements: Lower Guadiana River. Geoderma 2013, 193–194, 265–274. [Google Scholar] [CrossRef] [Green Version]
- De La Fuente, V.; Rufo, L.; Rodríguez, N.; Amils, R.; Zuluaga, J. Metal accumulation screening of the Río Tinto flora (Huelva, Spain). Biol. Trace Elem. Res. 2010, 134, 318–341. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.S.; Abreu, M.M.; Macías, F. Rehabilitation of mining areas through integrated biotechnological approach: Technosols derived from organic/inorganic wastes and autochthonous plant development. Chemosphere 2019, 224, 765–775. [Google Scholar] [CrossRef]
- Bouazama, S.; Harhar, H.; Costa, J.; Desjobert, J.M.; Talbaoui, A.; Tabyaoui, M. Chemical composition and antibacterial activity of the essential oils of Lavandula pedunculata and Lavandula dentata. J. Mater. Environ. Sci. 2017, 8, 2154–2160. [Google Scholar]
- Costa, P.; Medronho, B.; Gonçalves, S.; Romano, A. Cyclodextrins enhance the antioxidant activity of essential oils from three Lamiaceae species. Ind. Crops Prod. 2015, 70, 341–346. [Google Scholar] [CrossRef]
- Costa, P.; Gonçalves, S.; Valentão, P.; Andrade, P.B.; Almeida, C.; Nogueira, J.M.F.; Romano, A. Metabolic profile and biological activities of Lavandula pedunculata subsp. lusitanica (Chaytor) Franco: Studies on the essential oil and polar extracts. Food Chem. 2013, 141, 2501–2506. [Google Scholar] [CrossRef]
- Lopes, C.L.; Pereira, E.; Soković, M.; Carvalho, A.M.; Barata, A.M.; Lopes, V.; Rocha, F.; Calhelha, R.C.; Barros, L.; Ferreira, I.C.F.R. Phenolic Composition and Bioactivity of Lavandula pedunculata (Mill.) Cav. Samples from Different Geographical Origin. Molecules 2018, 23, 1037. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F.; Baptista, R.; Ladeiras, D.; Madureira, A.M.; Teixeira, G.; Rosado, C.; Fernandes, A.S.; Ascensão, L.; Silva, C.O.; Reis, C.P.; et al. Production and characterization of nanoparticles containing methanol extracts of Portuguese Lavenders. Meas. J. Int. Meas. Confed. 2015, 74, 170–177. [Google Scholar] [CrossRef]
- CCME. Canadian Soil Quality Guidelines for the Protection of Environmental and Human Health. Soil Quality Index 1.0 Technical Report; CCME: Winnipeg, Canada, 2007. [Google Scholar]
- Schützendübel, A.; Polle, A. Plant responses to abiotic stresses: Heavy metal-induced oxidative stress and protection by mycorrhization. J. Exp. Bot. 2002, 53, 1351–1365. [Google Scholar] [CrossRef]
- Parra, A.; Zornoza, R.; Conesa, E.; Gómez-López, M.D.; Faz, A. Seedling emergence, growth and trace elements tolerance and accumulation by Lamiaceae species in a mine soil. Chemosphere 2014, 113, 132–140. [Google Scholar] [CrossRef]
- Santos, E.S.; Abreu, M.M.; de Varennes, A.; Macías, F.; Leitão, S.; Cerejeira, M.J. Evaluation of chemical parameters and ecotoxicity of a soil developed on gossan following application of polyacrylates and growth of Spergularia purpurea. Sci. Total Environ 2013, 461–462, 360–370. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants, 2rd ed.; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar] [CrossRef]
- Mendez, M.O.; Maier, R.M. Phytoremediation of mine tailings in temperate and arid environments. Rev. Environ. Sci. Biotechnol. 2008, 7, 47–59. [Google Scholar] [CrossRef]
- Freitas, H.; Prasad, M.N.V.; Pratas, J. Plant community tolerant to trace elements growing on the degraded soils of São Domingos mine in the south east of Portugal: Environmental implications. Environ. Int. 2004, 30, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Alloway, B.J. Heavy Metals in Soils; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-4469-1. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Carvalho, L.C.; Santos, E.S.; Abreu, M.M. Unraveling the crucial role of the ascorbate-glutathione cycle in the resilience of Cistus monspeliensis L. to withstand high As concentrations. Ecotoxicol. Environ. Saf. 2019, 171, 389–397. [Google Scholar] [CrossRef]
- Manios, T.; Stentiford, E.I.; Millner, P.A. The effect of heavy metals accumulation on the chlorophyll concentration of Typha latifolia plants, growing in a substrate containing sewage sludge compost and watered with metaliferus water. Ecol. Eng. 2003, 37, 1441–1451. [Google Scholar] [CrossRef]
- Zengin, F.K.; Munzuroglu, O. Effects of some heavy metals on content of chlorophyll, proline and some antioxidant chemicals in bean (Phaseolus vulgaris L.) seedlings. Acta Biol. Crac. Ser. Bot. 2005, 47, 157–164. [Google Scholar]
- Alonso-Blázquez, N.; García-Gómez, C.; Fernández, M.D. Influence of Zn-contaminated soils in the antioxidative defence system of wheat (Triticum aestivum) and maize (Zea mays) at different exposure times: Potential use as biomarkers. Ecotoxicology 2015, 24, 279–291. [Google Scholar] [CrossRef]
- Rascio, N.; La Rocca, N. Resurrection plants: The puzzle of surviving extreme vegetative desiccation. CRC Crit. Rev. Plant Sci. 2005, 24, 209–225. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione is a key player in metal-induced oxidative stress defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Lago, D.; Carvalho, L.C.; Santos, E.S.; Abreu, M.M. The physiological mechanisms underlying the ability of Cistus monspeliensis L. from São Domingos mine to withstand high Zn concentrations in soils. Ecotoxicol. Environ. Saf. 2016, 129, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Pérez-López, R.; Álvarez-Valero, A.M.; Nieto, J.M.; Sáez, R.; Matos, J.X. Use of sequential extraction procedure for assessing the environmental impact at regional scale of the São Domingos Mine (Iberian Pyrite Belt). Appl. Geochem. 2008, 23, 3452–3463. [Google Scholar] [CrossRef]
- Álvarez-Valero, A.M.; Pérez-López, R.; Matos, J.; Capitán, M.A.; Nieto, J.M.; Sáez, R.; Delgado, J.; Caraballo, M. Potential environmental impact at São Domingos mining district (Iberian Pyrite Belt, SW Iberian Peninsula): Evidence from a chemical and mineralogical characterization. Environ. Geol. 2008, 55, 1797–1809. [Google Scholar] [CrossRef]
- Quental, L.; Bourguignon, A.; Sousa, A.J.; Batista, M.J.; Brito, M.G.; Tavares, T.; Abreu, M.M.; Vairinho, M.M.; Cottard, F. MINEO Project—Assessing and monitoring the environmental impact of mining in Europe using Advanced Earth Observation Techniques. In MINEO Southern Europe Environment Test Site: Contamination Impact Mapping and Modelling: Final Report; 2002. Available online: https://repositorio.lneg.pt/handle/10400.9/3268 (accessed on 30 November 2021).
- IUSS Working Group WRB. International soil classification system for naming soils and creating legends for soil maps. In World Reference Base for Soil Resources 2014; World Soil Resources Reports No. 106; FAO Publications: Rome, Italy, 2014. [Google Scholar] [CrossRef]
- Santos, E.S.; Balseiro-Romero, M.; Abreu, M.M.; Macías, F. Bioextracts of Cistus ladanifer L. growing in São Domingos mine as source of valuable compounds. J. Geochem. Explor. 2017, 174, 84–90. [Google Scholar] [CrossRef]
- IPMA—Instituto Português do Mar e da Atmosfera. 2021. Available online: https://www.ipma.pt/pt/oclima/series.longas/ (accessed on 15 September 2021).
- Acácio, V.; Holmgren, M.; Moreira, F.; Mohren, G.M.J. Oak persistence in Mediterranean landscapes: The combined role of management, topography, and wildfires. Ecol. Soc. 2010, 15, 40. [Google Scholar] [CrossRef]
- Nakamura, S.; Nitta, Y.; Goto, Y. Leaf characteristics and shape of sago palm (Metroxylon sagu Rottb.) for developing a method of estimating leaf area. Plant Prod. Sci. 2004, 7, 198–203. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Feng, M.H.; Shan, X.Q.; Zhang, S.Z.; Wen, B. Comparison of a rhizosphere-based method with other one-step extraction methods for assessing the bioavailability of soil metals to wheat. Chemosphere 2005, 59, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Bell, P.F.; Xie, B.; Higby, J.R.; Amínha, N. Digestion of NIST peach leaves using sealed vessels and inexpensive microwave ovens. Commun. Soil Sci. Plant Anal. 2000, 31, 1897–1903. [Google Scholar] [CrossRef]
- Lago-Vila, M.; Arenas-Lago, D.; Andrade, L.; Vega, F.A. Phytoavailable content of metals in soils from copper mine tailings (Touro mine, Galicia, Spain). J. Geochem. Explor. 2014, 147, 159–166. [Google Scholar] [CrossRef]
- Richardson, A.D.; Duigan, S.P.; Berlyn, G.P. An evaluation of noninvasive methods to estimate foliar chlorophyll content. New Phytol. 2002, 153, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Creissen, G.; Firmin, J.; Fryer, M.; Kular, B.; Leyland, N.; Reynolds, H.; Pastori, G.; Wellburn, F.; Baker, N.; Wellburn, A.; et al. Elevated glutathione biosynthetic capacity in the chloroplasts of transgenic tobacco plants paradoxically causes increased oxidative stress. Plant Cell 1999, 11, 1277–1291. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Amâncio, S. Antioxidant defence system in plantlets transferred from in vitro to ex vitro: Effects of increasing light intensity and CO2 concentration. Plant Sci. 2002, 162, 33–40. [Google Scholar] [CrossRef]
- Okamura, M. An improved method for determination of L-ascorbic acid and L-dehydroascorbic acid in blood plasma. Clin. Chim. Acta 1980, 103, 259–268. [Google Scholar] [PubMed]
- Anderson, J.V.; Chevone, B.I.; Hess, J.L. Seasonal variation in the antioxidant system of eastern white pine needles: Evidence for thermal dependence. Plant Physiol. 1992, 98, 501–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.W.; Cunningham, S.D. Lead phytoextraction: Species variation in lead uptake and translocation. New Phytol. 1996, 134, 75–84. [Google Scholar] [CrossRef]
- Abreu, M.M.; Tavares, M.T.; Batista, M.J. Potential use of Erica andevalensis and Erica australis in phytoremediation of sulphide mine environments: São Domingos, Portugal. J. Geochem. Explor. 2008, 96, 210–222. [Google Scholar] [CrossRef] [Green Version]
- Brooks, R.R. Phytoremediation by volatilisation. In Plants that Hyper-Accumulate Heavy Metals: Their Role in Phytoremediation, Microbiology, Archaeology, Mineral Exploration and Phytomining; CAB International: New York, NY, USA, 1998. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Units | CS | US | |
|---|---|---|---|
| pHH2O | - | 3.51 ± 0.06 a | 5.54 ± 0.16 b |
| Organic C | g kg−1 | 5.03 ± 0.25 a | 4.05 ± 0.24 b |
| Total N(Kjeldahl) | g kg−1 | 0.16 ± 0.01 a | 0.15 ± 0.01 a |
| C/N | - | 31.44 ± 0.26 a | 27.00 ± 0.25 b |
| Extractable P | mg kg−1 | 17.4 ± 0.03 a | 5.28 ± 1.48 b |
| Extractable K | mg kg−1 | 459 ± 210 a | 153 ± 13 b |
| Total Concentrations of Elements | |||
| As | mg kg−1 | 9993 * ± 1190 a | 18.7 * ± 0.9 b |
| Ca | g kg−1 | <0.1 b | 1.0 ± 0.0 a |
| Cd | mg kg−1 | 0.9 ± 0.4 a | 0.9 ± 0.2 a |
| Cr | mg kg−1 | 104 * ± 10 a | <2 b |
| Cu | mg kg−1 | 311 * ± 33 a | 61.3 ± 22.9 b |
| Fe | g kg−1 | 236 ± 23 a | 33.7 ± 2.8 b |
| K | g kg−1 | 3.3 ± 0.1 b | 18.9 ± 3.1 a |
| Mg | g kg−1 | <0.1 b | 3.9 ± 0.9 a |
| Mn | mg kg−1 | 50.3 ± 7.0 b | 256 ± 175 a |
| Mo | mg kg−1 | 11.0 ± 1.0 a | <2 b |
| Na | g kg−1 | 0.8 ± 0.1 b | 3.9 ± 0.2 a |
| Ni | mg kg−1 | 16.0 ± 2.0 b | 42.7 ± 15.3 a |
| Pb | g kg−1 | 477 * ± 168 a | 33.4 ± 1.6 b |
| Sb | mg kg−1 | 2123 * ± 40 a | 1.6 ± 0.1 b |
| Zn | mg kg−1 | 111 ± 10 a | 69.3 ± 25.1 b |
| Elements’ Concentrations in the Soil Available Fraction | |||
| As | mg kg−1 | 2.41 ± 0.25 a | 0.34 ± 0.19 b |
| Ca | 573 ± 14 a | 297 ± 17 b | |
| Cd | 0.08 ± 0.03 a | <dl | |
| Cr | 0.01 ± 0.01 b | 0.12 ± 0.02 a | |
| Cu | 0.05 ± 0.04 a | <dl | |
| Fe | 31.2 ± 3.1 b | 80.0 ± 5.1 a | |
| K | 8.86 ± 2.21 b | 52.8 ± 7.5 a | |
| Mg | 75.3 ± 4.3 b | 107 ± 5 a | |
| Mn | 9.35 ± 0.70 a | 6.4 ± 0.4 b | |
| Mo | 0.13 ± 0.09 a | <dl | |
| Na | 69.0 ± 6.7 a | 36.7 ± 2.4 b | |
| Ni | <dl | 0.04 ± 0.01 a | |
| Pb | 2.28 ± 0.65 a | <0.01 b | |
| Sb | 0.01 ± 0.06 a | <dl | |
| Zn | 3.58 ± 1.08 a | 0.78 ± 0.48 b | |
| Element | Units | CS-LC | CS-L | US-LC | US-L |
|---|---|---|---|---|---|
| Roots | |||||
| As | mg kg−1 | 416 ± 104 a | 292 ± 110 a | 23.6 ± 11.9 b | 100 ± 70 b |
| Ca | g kg−1 | 7.87 ± 4.51 a | 8.35 ± 1.16 a | 5.64 ± 1.78 b | 6.30 ± 2.31 ab |
| Cd | mg kg−1 | 32.3 ± 10.0 a | 28.3 ± 12.8 a | <0.3 b | <0.3 b |
| Cr | mg kg−1 | 10.1 ± 4.8 a | 7.66 ± 5.7 ab | 6.05 ± 2.56 b | <2 c |
| Cu | mg kg−1 | 60.1 ± 12.3 a | 45.4 ± 13.2 ab | 29.5 ± 12.5 b | 32.2 ± 21.6 b |
| Fe | g kg−1 | 5.64 ± 1.31 a | 3.64 ± 1.38 b | 0.98 ± 0.46 c | 1.21 ± 0.17 c |
| K | g kg−1 | 5.22 ± 1.12 c | 4.56 ± 0.67 c | 9.86 ± 3.06 b | 15.26 ± 6.10 a |
| Mg | g kg−1 | 6.31 ± 1.47 a | 5.37 ± 1.20 a | 2.52 ± 0.83 b | 3.16 ± 1.14 b |
| Mn | mg kg−1 | 106 ± 19 a | 110 ± 23 a | 60.8 ± 30.0 b | 87.5 ± 33.7 ab |
| Mo | mg kg−1 | 4.82 ± 2.67 a | 6.07 ± 2.25 a | <1 b | <1 b |
| Na | g kg−1 | 12.1 ± 2.0 a | 10.3 ± 3.2 a | 2.13 ± 0.41 b | 3.28 ± 1.48 b |
| Ni | mg kg−1 | <1 c | <1 c | 38.6 ± 28.7 b | 152 ± 65 a |
| Pb | mg kg−1 | 907 ± 215 a | 603 ± 213 a | <3 b | <3 b |
| Sb | mg kg−1 | <0.1 a | <0.1 a | <0.1 a | <0.1 a |
| Zn | mg kg−1 | 330 ± 51 a | 485 ± 164 a | 175 ± 92 b | 268 ± 105 ab |
| Element | Units | Shoots | |||
| As | mg kg−1 | 41.1 ± 21.6 a | 50.8 ± 18.4 a | 9.71 ± 5.66 b | 13.3 ± 9.2 b |
| Ca | g kg−1 | 6.40 ± 1.53 a | 8.14 ± 2.48 a | 6.25 ± 2.17 a | 7.68 ± 0.58 a |
| Cd | mg kg−1 | <0.3 a | <0.3 a | <0.3 a | <0.3 a |
| Cr | mg kg−1 | <2 a | <2 a | <2 a | <2 a |
| Cu | mg kg−1 | 4.12 ± 2.15 b | 4.10 ± 2.10 b | 11.6 ± 1.5 a | 10.7 ± 4.1 a |
| Fe | g kg−1 | 0.47 ± 0.23 c | 0.71 ± 0.30 bc | 0.78 ± 0.27 b | 1.05 ± 0.14 a |
| K | g kg−1 | 11.1 ± 3.8 b | 12.3 ± 4.6 b | 39.3 ± 12.6 a | 42.1 ± 3.7 a |
| Mg | g kg−1 | 5.58 ± 1.31 a | 6.15 ± 2.50 a | 3.37 ± 1.33 a | 5.32 ± 1.44 a |
| Mn | mg kg−1 | 564 ± 164 a | 522 ± 197 a | 478 ± 161 a | 525 ± 169 a |
| Mo | mg kg−1 | <1 b | 6.01 ± 3.11 a | <1 b | <1 b |
| Na | g kg−1 | 4.68 ± 0.93 a | 6.43 ± 2.93 a | 2.61 ± 0.95 b | 2.85 ± 0.43 b |
| Ni | mg kg−1 | 6.15 ± 3.42 ab | 31.8 ± 19.5 a | 3.48 ± 1.82 b | 5.12 ± 1.62 ab |
| Pb | mg kg−1 | 53.2 ± 32.45 a | 60.6 ± 20.0 a | <3 b | <3 b |
| Sb | mg kg−1 | <0.1 a | <0.1 a | <0.1 a | <0.1 a |
| Zn | mg kg−1 | 179 ± 40 b | 196 ± 68 b | 228 ± 76 b | 337 ± 18 a |
| Element | CS-LC | CS-L | US-LC | US-L |
|---|---|---|---|---|
| TranslC | ||||
| As | 0.10 | 0.17 | 0.41 | 0.13 |
| Ca | 0.81 | 0.97 | 1.11 | 1.22 |
| Cd | * | * | * | * |
| Cr | * | * | * | * |
| Cu | 0.07 | 0.09 | 0.39 | 0.33 |
| Fe | 0.08 | 0.20 | 0.80 | 0.87 |
| K | 2.13 | 2.70 | 3.99 | 2.76 |
| Mg | 0.88 | 1.15 | 1.34 | 1.68 |
| Mn | 5.32 | 4.75 | 7.86 | 6.00 |
| Mo | * | 0.99 | * | * |
| Na | 0.39 | 0.62 | 1.23 | 0.87 |
| Ni | * | * | 0.09 | 0.03 |
| Pb | 0.06 | 0.10 | * | * |
| Sb | * | * | * | |
| Zn | 0.54 | 0.40 | 1.30 | 1.26 |
| TransferC | ||||
| As | <0.01 | 0.01 | 0.52 | 0.71 |
| Ca | 64.00 | 81.40 | 6.25 | 7.68 |
| Cd | * | * | * | * |
| Cr | * | * | * | * |
| Cu | 0.01 | 0.01 | 0.19 | 0.17 |
| Fe | <0.01 | <0.01 | 0.02 | 0.03 |
| K | 3.36 | 3.73 | 2.08 | 2.23 |
| Mg | 55.80 | 61.50 | 0.86 | 1.36 |
| Mn | 11.21 | 10.38 | 1.87 | 2.05 |
| Mo | * | 0.55 | * | * |
| Na | 5.85 | 8.04 | 0.67 | 0.73 |
| Ni | 0.38 | 1.99 | 0.08 | 0.12 |
| Pb | 1.59 | 1.81 | * | * |
| Sb | * | * | * | * |
| Zn | 1.61 | 1.77 | 3.29 | 4.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arenas-Lago, D.; Carvalho, L.C.; Santos, E.S.; Abreu, M.M. Influence of Seed Source and Soil Contamination on Ecophysiological Responses of Lavandula pedunculata in Rehabilitation of Mining Areas. Plants 2022, 11, 105. https://doi.org/10.3390/plants11010105
Arenas-Lago D, Carvalho LC, Santos ES, Abreu MM. Influence of Seed Source and Soil Contamination on Ecophysiological Responses of Lavandula pedunculata in Rehabilitation of Mining Areas. Plants. 2022; 11(1):105. https://doi.org/10.3390/plants11010105
Chicago/Turabian StyleArenas-Lago, Daniel, Luisa C. Carvalho, Erika S. Santos, and Maria Manuela Abreu. 2022. "Influence of Seed Source and Soil Contamination on Ecophysiological Responses of Lavandula pedunculata in Rehabilitation of Mining Areas" Plants 11, no. 1: 105. https://doi.org/10.3390/plants11010105
APA StyleArenas-Lago, D., Carvalho, L. C., Santos, E. S., & Abreu, M. M. (2022). Influence of Seed Source and Soil Contamination on Ecophysiological Responses of Lavandula pedunculata in Rehabilitation of Mining Areas. Plants, 11(1), 105. https://doi.org/10.3390/plants11010105