
Defence Responses Associated with Elicitor-Induced, Cultivar-Associated Resistance to Latania Scale in Kiwifruit

 ,
,  and
and
Abstract
1. Introduction
- Use phytohormone and gene expression measurements to investigate whether cultivar-specific resistance is determined by responsiveness to different elicitors (including the pest itself, as well as elicitors of the antagonistic SA versus JA pathways).
- Examine the effects of hormonal crosstalk on the host–pest interaction.
2. Results
2.1. MeJA Elicitor Application Did Not Affect Latania Scale Growth
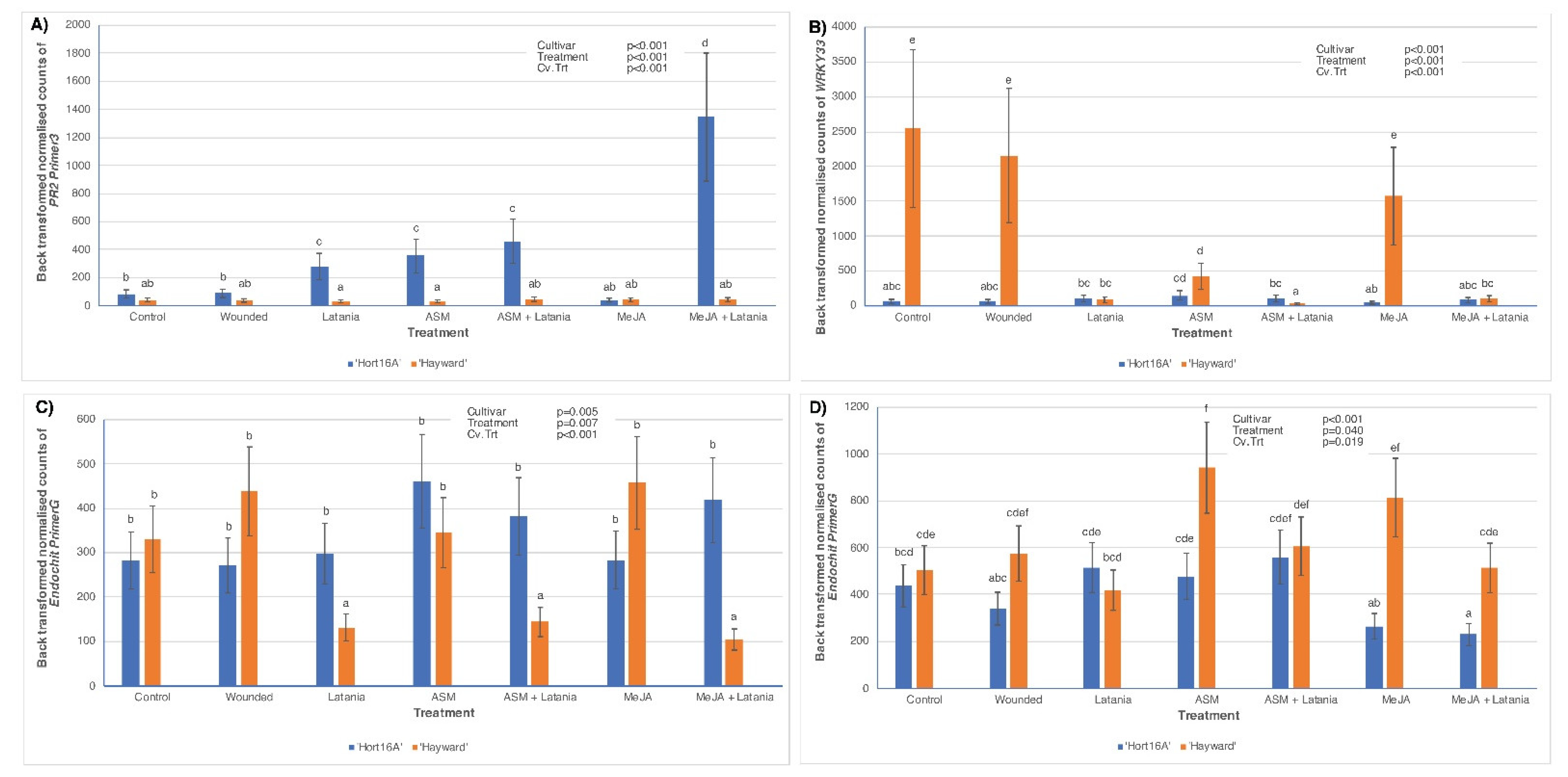
2.2. Changes in Phytohormones and Gene Expression in Elicitor-Induced, Cultivar-Associated Resistance to Latania Scale
2.2.1. Phytohormone Measurements
2.2.2. Gene Expression
3. Discussion
3.1. Cultivar Response to SA Pathway Elicitation
3.2. Cultivar Response to JA Pathway Elicitation
3.3. The Importance of Crosstalk
3.4. Future Directions and Impact
4. Materials and Methods
4.1. Plants and Insects
4.2. Treatment Application and Sampling Procedure
4.3. Measurement of Latania Scale Growth/Survival in the Methyl Jasmonate (MeJA) Treatment
4.4. Phytohormone Measurements
4.5. Gene Expression Measurements by Quantitative Polymerase Chain Reaction (qPCR)
4.6. Gene Expression Measurements by NanoString
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fresh Facts. New Zealand Horticulture. 2019. Available online: https://www.freshfacts.co.nz/files/freshfacts-2019.pdf (accessed on 29 June 2021).
- Ferguson, A.R.; Huang, H.W. Genetic resources of kiwifruit: Domestication and breeding. In Horticultural Reviews; John Wiley and Sons: Hoboken, NJ, USA, 2007; Volume 33, pp. 1–121. [Google Scholar]
- Hill, M.G.; Wurms, K.V.; Davy, M.W.; Gould, E.; Allan, A.; Mauchline, N.A.; Luo, Z.; Chee, A.A.; Stannard, K.; Storey, R.D.; et al. Transcriptome analysis of kiwifruit (Actinidia chinensis) bark in response to armoured scale insect (Hemiberlesia lataniae) feeding. PLoS ONE 2015, 10, e0141664. [Google Scholar] [CrossRef]
- Edwards, R.; Carraher, C.; Poulton, J.; Sandanayaka, M.; Todd, J.H.; Dobson, S.; Mauchline, N.; Hill, G.; McKenna, C.; Newcomb, R.D. DNA diagnostics of three armored scale species on kiwifruit in New Zealand. J. Econ. Entomol. 2008, 101, 1944–1949. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.G.; Mauchline, N.A.; Connolly, P.G.; Maher, B.J. Measuring resistance to armoured scale insects in kiwifruit (Actinidia) germplasm. N. Z. J. Crop Hortic. Sci. 2010, 38, 69–85. [Google Scholar] [CrossRef]
- CABI International. Hemiberlesia lataniae (Latania Scale). Available online: https://www.cabi.org/isc/datasheet/26647 (accessed on 29 June 2021).
- Kabashima, J.N.; Dreistadt, S.H. Pest Notes: Scales. Available online: https://www.ipm.ucanr.edu/PMG/PESTNOTES/pn7408.html (accessed on 23 July 2021).
- Erb, M.; Meldau, S.; Howe, G.A. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 2012, 17, 250–259. [Google Scholar] [CrossRef]
- Smith, C.M.; Clement, S.L. Molecular basis of plant resistance to arthropods. Annu. Rev. Entomol. 2012, 57, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.G.; Mauchline, N.A.; Jones, M.K.; Sutherland, P.W. The response of resistant kiwifruit (Actinidia chinensis) to armoured scale insect (Diaspididae) feeding. Arthropod-Plant Interact. 2011, 5, 149–161. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal modulation of plant immunity. Annu. Rev. Cell. Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Damian-Zamacona, S.; Toledo-Ibelles, P.; Ibarra-Abundis, M.Z.; Uribe-Figueroa, L.; Hernandez-Lemus, E.; Macedo-Alcibia, K.P.; Delgado-Coello, B.; Mas-Oliva, J.; Reyes-Grajeda, J.P. Early transcriptomic response to LDL and oxLDL in human vascular smooth muscle cells. PLoS ONE 2016, 11, e0163924. [Google Scholar] [CrossRef]
- Meimoun, P.; Mordret, E.; Langlade, N.B.; Balzergue, S.; Arribat, S.; Bailly, C.; El-Maarouf-Bouteau, H. Is gene transcription involved in seed dry after-ripening? PLoS ONE 2014, 9, e86442. [Google Scholar] [CrossRef]
- Mosqueira, M.; Willmann, G.; Zeiger, U.; Khurana, T.S. Expression profiling reveals novel hypoxic biomarkers in peripheral blood of adult mice exposed to chronic hypoxia. PLoS ONE 2012, 7, e37497. [Google Scholar] [CrossRef][Green Version]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance-salicylic acid and its functional analogs. Curr. Plant Biol. 2019, 17, 48–59. [Google Scholar] [CrossRef]
- Dempsey, D.A.; Klessig, D.F. How does the multifaceted plant hormone salicylic acid combat disease in plants and are similar mechanisms utilized in humans? BMC Biol. 2017, 15, 11. [Google Scholar] [CrossRef]
- Klessig, D.F.; Tian, M.Y.; Choi, H.W. Multiple targets of salicylic acid and its derivatives in plants and animals. Front. Immunol. 2016, 7, 10. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, R365–R373. [Google Scholar] [CrossRef]
- Zheng, X.-y.; Zhou, M.; Yoo, H.; Pruneda-Paz, J.L.; Spivey, N.W.; Kay, S.A.; Dong, X. Spatial and temporal regulation of biosynthesis of the plant immune signal salicylic acid. Proc. Natl. Acad. Sci. USA 2015, 112, 9166–9173. [Google Scholar] [CrossRef] [PubMed]
- Aerts, N.; Mendes, M.P.; Van Wees, S.C.M. Multiple levels of crosstalk in hormone networks regulating plant defense. Plant J. 2021, 105, 489–504. [Google Scholar] [CrossRef]
- Spoel, S.H.; van Ooijen, G. Circadian redox signaling in plant immunity and abiotic stress. Antioxid. Redox Signal. 2014, 20, 3024–3039. [Google Scholar] [CrossRef] [PubMed]
- Hickman, R.; Van Verk, M.C.; Van Dijken, A.J.H.; Mendes, M.P.; Vroegop-Vos, I.A.; Caarls, L.; Steenbergen, M.; Van der Nagel, I.; Wesselink, G.J.; Jironkin, A.; et al. Architecture and dynamics of the jasmonic acid gene regulatory network. Plant Cell 2017, 29, 2086–2105. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef]
- Thatcher, L.F.; Anderson, J.P.; Singh, K.B. Plant defence responses: What have we learnt from Arabidopsis? Funct. Plant Biol. 2005, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed]
- Cellini, A.; Fiorentini, L.; Buriani, G.; Yu, J.; Donati, I.; Cornish, D.A.; Novak, B.; Costa, G.; Vanneste, J.L.; Spinelli, F. Elicitors of the salicylic acid pathway reduce incidence of bacterial canker of kiwifruit caused by Pseudomonas syringae pv. actinidiae. Ann. Appl. Biol. 2014, 165, 441–453. [Google Scholar] [CrossRef]
- De Jong, H.; Reglinksi, T.; Elmer, P.A.G.; Wurms, K.; Vanneste, J.L.; Guo, L.F.; Alavi, M. Integrated use of Aureobasidium pullulans strain CG163 and acibenzolar-S-methyl for management of bacterial canker in kiwifruit. Plants 2019, 8, 287. [Google Scholar] [CrossRef] [PubMed]
- Michelotti, V.; Lamontanara, A.; Buriani, G.; Orru, L.; Cellini, A.; Donati, I.; Vanneste, J.L.; Cattivelli, L.; Tacconi, G.; Spinelli, F. Comparative transcriptome analysis of the interaction between Actinidia chinensis var. chinensis and Pseudomonas syringae pv. actinidiae in absence and presence of acibenzolar-S-methyl. BMC Genom. 2018, 19, 585. [Google Scholar] [CrossRef]
- Wurms, K.; Cui, W.; Ah-Chee, A.; Rees-George, J.; Bublin, M.; Breiteneder, H. Down Regulation of putative defence-associated transcripts correlates with ripe rot symptoms on kiwifruit (Actinidia chinensis). J. Phytopathol. 2011, 159, 435–442. [Google Scholar] [CrossRef]
- Wurms, K.; Gould, E.; Ah Chee, A.; Taylor, J.; Curran, B.; Reglinski, T. Elicitor induction of defence genes and reduction of bacterial canker in kiwifruit. N. Z. Plant Prot. 2017, 70, 272–284. [Google Scholar] [CrossRef][Green Version]
- Wurms, K.V.; Hardaker, A.J.; Ah Chee, A.; Bowen, J.; Phipps, J.; Taylor, J.; Jensen, D.; Cooney, J.; Wohlers, M.; Reglinski, T. Phytohormone and putative defense gene expression differentiates the response of ‘Hayward’ kiwifruit to Psa and Pfm infections. Front. Plant Sci. 2017, 8, 1366. [Google Scholar] [CrossRef]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef]
- Gamir, J.; Darwiche, R.; van’t Hof, P.; Choudhary, V.; Stumpe, M.; Schneiter, R.; Mauch, F. The sterol-binding activity of pathogenesis-related protein 1 reveals the mode of action of an antimicrobial protein. Plant J. 2017, 89, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.F.; Behmer, S.T. Insect sterol nutrition: Physiological mechanisms, ecology, and applications. Annu. Rev. Entomol. 2020, 65, 251–271. [Google Scholar] [CrossRef]
- Minic, Z. Physiological roles of plant glycoside hydrolases. Planta 2008, 227, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Morant, A.V.; Jorgensen, K.; Jorgensen, C.; Paquette, S.M.; Sanchez-Perez, R.; Moller, B.L.; Bak, S. Beta-glucosidases as detonators of plant chemical defense. Phytochemistry 2008, 69, 1795–1813. [Google Scholar] [CrossRef] [PubMed]
- Behr, M.; Neutelings, G.; El Jaziri, M.; Baucher, M. You want it sweeter: How glycosylation affects plant response to oxidative stress. Front. Plant Sci. 2020, 11, 1443. [Google Scholar] [CrossRef]
- Le Roy, J.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation is a major regulator of phenylpropanoid availability and biological activity in plants. Front. Plant Sci. 2016, 7, 735. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Umashankar, S.; Rai, M.; Kiat, L.B.; Bing, J.A.S.; Swarup, S. Coordinate regulation of metabolite glycosylation and stress hormone biosynthesis by TT8 in Arabidopsis. Plant Physiol. 2016, 171, 2499–2515. [Google Scholar] [CrossRef]
- Wurms, K.V.; George, M.P.; Lauren, D.R. Involvement of phenolic compounds in host resistance against Botrytis cinerea in leaves of the two commercially important kiwifruit (Actinidia chinensis and A. deliciosa) cultivars. N. Z. J. Crop Hortic. Sci. 2003, 31, 221–233. [Google Scholar] [CrossRef]
- Birkenbihl, R.P.; Kracher, B.; Somssich, I.E. Induced genome-wide binding of three Arabidopsis WRKY transcription factors during early MAMP-triggered immunity. Plant Cell 2017, 29, 20–38. [Google Scholar] [CrossRef]
- Li, S.J.; Fu, Q.T.; Chen, L.G.; Huang, W.D.; Yu, D.Q. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 coordinate induction of plant thermotolerance. Planta 2011, 233, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.Y.; Abu Qamar, S.; Chen, Z.X.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef]
- Alba, J.M.; Schimmel, B.C.J.; Glas, J.J.; Ataide, L.M.S.; Pappas, M.L.; Villarroel, C.A.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Spider mites suppress tomato defenses downstream of jasmonate and salicylate independently of hormonal crosstalk. New Phytol. 2015, 205, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Kaloshian, I.; Walling, L.L. Hemipterans as plant pathogens. Annu. Rev. Phytopathol. 2005, 43, 491–521. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Piotrowska, A. Conjugates of auxin and cytokinin. Phytochemistry 2009, 70, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Westfall, C.S.; Muehler, A.M.; Jez, J.M. Enzyme action in the regulation of plant hormone responses. J. Biol. Chem. 2013, 288, 19304–19311. [Google Scholar] [CrossRef]
- Cheng, C.; Cui, W.; Gea, L.; Hill, M.G.; Hoyte, S.M.; Mauchline, N.A.; McKenna, C.E.; Popowski, E.; Reglinski, T.; Rikkerink, E.H.A.; et al. Improving kiwifruit resistance to pests and diseases. In Utilising Plant Defence for Pest Control; Wurms, K.V., Popay, A.J., Bell, N.L., Jaspers, M.V., Eds.; New Zealand Plant Protection Society: Auckland, New Zealand, 2013; pp. 25–38. [Google Scholar]
- Mur, L.A.J.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol. 2006, 140, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef] [PubMed]
- Van Wees, S.C.M.; de Swart, E.A.M.; van Pelt, J.A.; van Loon, L.C.; Pieterse, C.M.J. Enhancement of induced disease resistance by simultaneous activation of salicylate- and jasmonate-dependent defense pathways in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2000, 97, 8711–8716. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef]
- Eitle, M.W.; Griesser, M.; Vankova, R.; Dobrev, P.; Aberer, S.; Forneck, A. Grape phylloxera (D. vitifoliae) manipulates SA/JA concentrations and signalling pathways in root galls of Vitis spp. Plant Physiol. Biochem. 2019, 144, 85–91. [Google Scholar] [CrossRef]
- Zhang, P.J.; Huang, F.; Zhang, J.M.; Wei, J.N.; Lu, Y.B. The mealybug Phenacoccus solenopsis suppresses plant defense responses by manipulating JA-SA crosstalk. Sci. Rep. 2015, 5, 7. [Google Scholar] [CrossRef][Green Version]
- Liu, Z.; Yan, J.P.; Li, D.K.; Luo, Q.; Yan, Q.; Liu, Z.B.; Ye, L.M.; Wang, J.M.; Li, X.F.; Yang, Y. UDP-glucosyltransferase71C5, a major glucosyltransferase, mediates abscisic acid homeostasis in Arabidopsis. Plant Physiol. 2015, 167, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- Nunes da Silva, M.; Vasconcelos, M.W.; Pinto, V.; Balestra, G.M.; Mazzaglia, A.; Gomez-Cadenas, A.; Carvalho, S.M.P. Role of methyl jasmonate and salicylic acid in kiwifruit plants further subjected to Psa infection: Biochemical and genetic responses. Plant Physiol. Biochem. 2021, 162, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Bulley, S.M.; Cooney, J.M.; Laing, W. Elevating ascorbate in Arabidopsis stimulates the production of abscisic acid, phseic acid, and to a lesser extent auxin (IAA) and jasmonares, resulting in increased expression of DHAR1 and multiple transcription factors associated with abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 6743. [Google Scholar] [CrossRef]
- Petriccione, M.; Mastrobuoni, F.; Zampella, L.; Scortichini, M. Reference gene selection for normalization of RT-qPCR gene expression data from Actinidia deliciosa leaves infected with Pseudomonas syringae pv. actinidiae. Sci. Rep. 2015, 5, 16961. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, R50. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Poelman, E.H.; Van Wees, S.C.M.; Dicke, M. Induced plant responses to microbes and insects. Front. Plant Sci. 2013, 4, 475. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vallet, A.; Lopez, G.; Ramos, B.; Delgado-Cerezo, M.; Riviere, M.P.; Llorente, F.; Fernandez, P.V.; Miedes, E.; Estevez, J.M.; Grant, M.; et al. Disruption of abscisic acid signaling constitutively activates Arabidopsis resistance to the necrotrophic fungus Plectosphaerella cucumerina. Plant Physiol. 2012, 160, 2109–2124. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Nardozza, S.; Boldingh, H.L.; Osorio, S.; Hoehne, M.; Wohlers, M.; Gleave, A.P.; MacRae, E.A.; Richardson, A.C.; Atkinson, R.G.; Sulpice, R.; et al. Metabolic analysis of kiwifruit (Actinidia deliciosa) berries from extreme genotypes reveals hallmarks for fruit starch metabolism. J. Exp. Bot. 2013, 64, 5049–5063. [Google Scholar] [CrossRef]
- Nanostring Technologies. PlexSet Reagents User Manual, 10040-04 ed.; NanoString Technologies: Seattle, WA, USA, 2018; p. 40. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Code | Treatment Details |
|---|---|
| Control | Control—plants left untreated. |
| Wounding | Sample leaf mechanically wounded one week after the start of the trial. |
| Latania | Water + 0.025% v/v DuWett® sprayed on day 1, 200+ latania scale crawlers applied to the sample leaf, one week later. |
| ASM | Actigard™ (ASM, 0.2 g/L) applied to all leaves on day 1. |
| ASM + Latania | Same treatment as above, plus latania scale application one week later. |
| MeJA | Methyl jasmonate (0.05% v/v) + 0.025% (v/v) DuWett® applied to all leaves on day 1. |
| MeJA + Latania | Same treatment as above, plus latania scale application one week later. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wurms, K.; Ah Chee, A.; Stannard, K.; Anderson, R.; Jensen, D.; Cooney, J.; Hedderley, D. Defence Responses Associated with Elicitor-Induced, Cultivar-Associated Resistance to Latania Scale in Kiwifruit. Plants 2022, 11, 10. https://doi.org/10.3390/plants11010010
Wurms K, Ah Chee A, Stannard K, Anderson R, Jensen D, Cooney J, Hedderley D. Defence Responses Associated with Elicitor-Induced, Cultivar-Associated Resistance to Latania Scale in Kiwifruit. Plants. 2022; 11(1):10. https://doi.org/10.3390/plants11010010
Chicago/Turabian StyleWurms, Kirstin, Annette Ah Chee, Kate Stannard, Rachelle Anderson, Dwayne Jensen, Janine Cooney, and Duncan Hedderley. 2022. "Defence Responses Associated with Elicitor-Induced, Cultivar-Associated Resistance to Latania Scale in Kiwifruit" Plants 11, no. 1: 10. https://doi.org/10.3390/plants11010010
APA StyleWurms, K., Ah Chee, A., Stannard, K., Anderson, R., Jensen, D., Cooney, J., & Hedderley, D. (2022). Defence Responses Associated with Elicitor-Induced, Cultivar-Associated Resistance to Latania Scale in Kiwifruit. Plants, 11(1), 10. https://doi.org/10.3390/plants11010010