
Morphology, Anatomy and Secondary Metabolites Investigations of Premna odorata Blanco and Evaluation of Its Anti-Tuberculosis Activity Using In Vitro and In Silico Studies





Abstract
:1. Introduction
2. Results and Discussion
2.1. Botanical Investigations
2.1.1. Macromorphological Characterization
Leaf
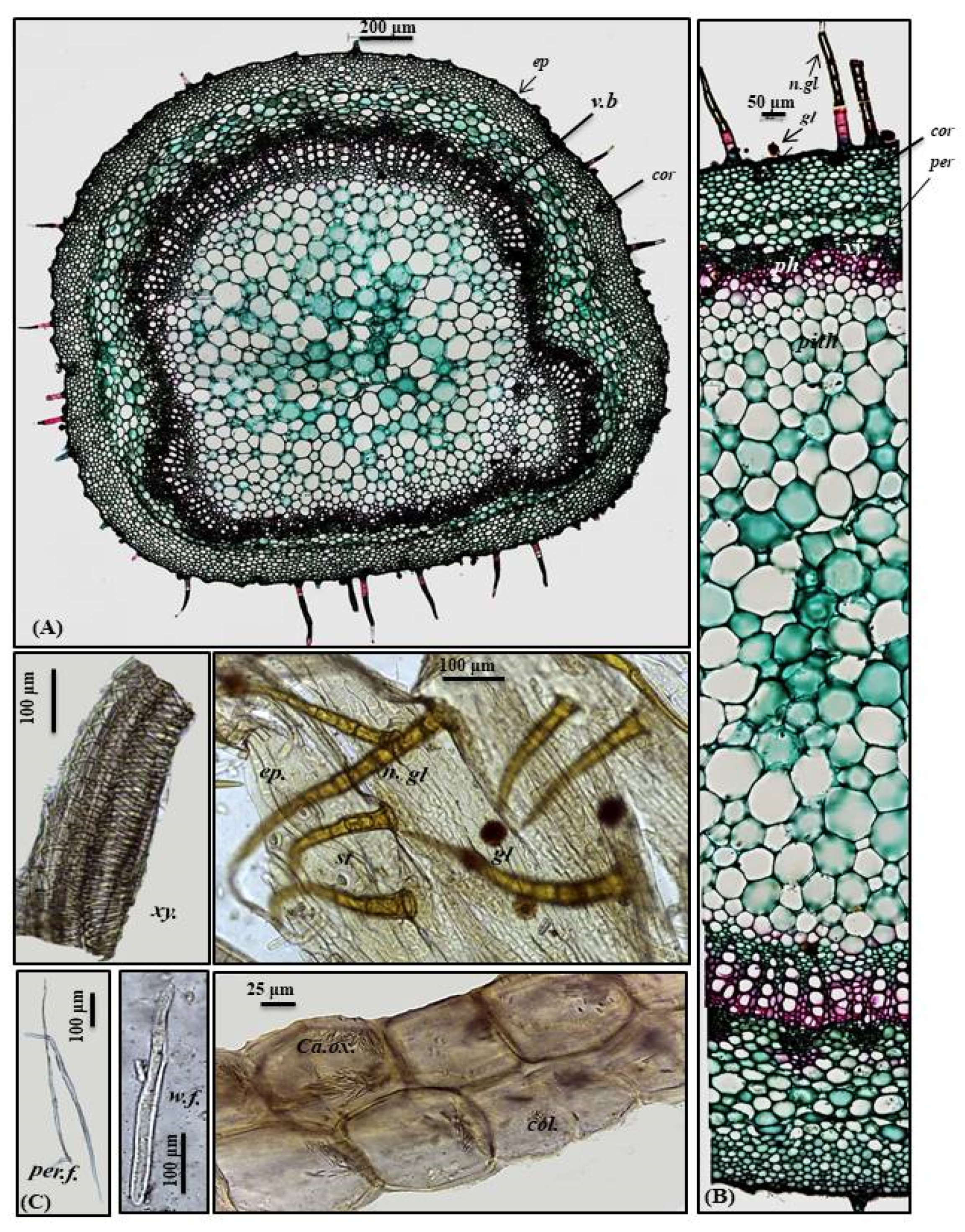
Stem
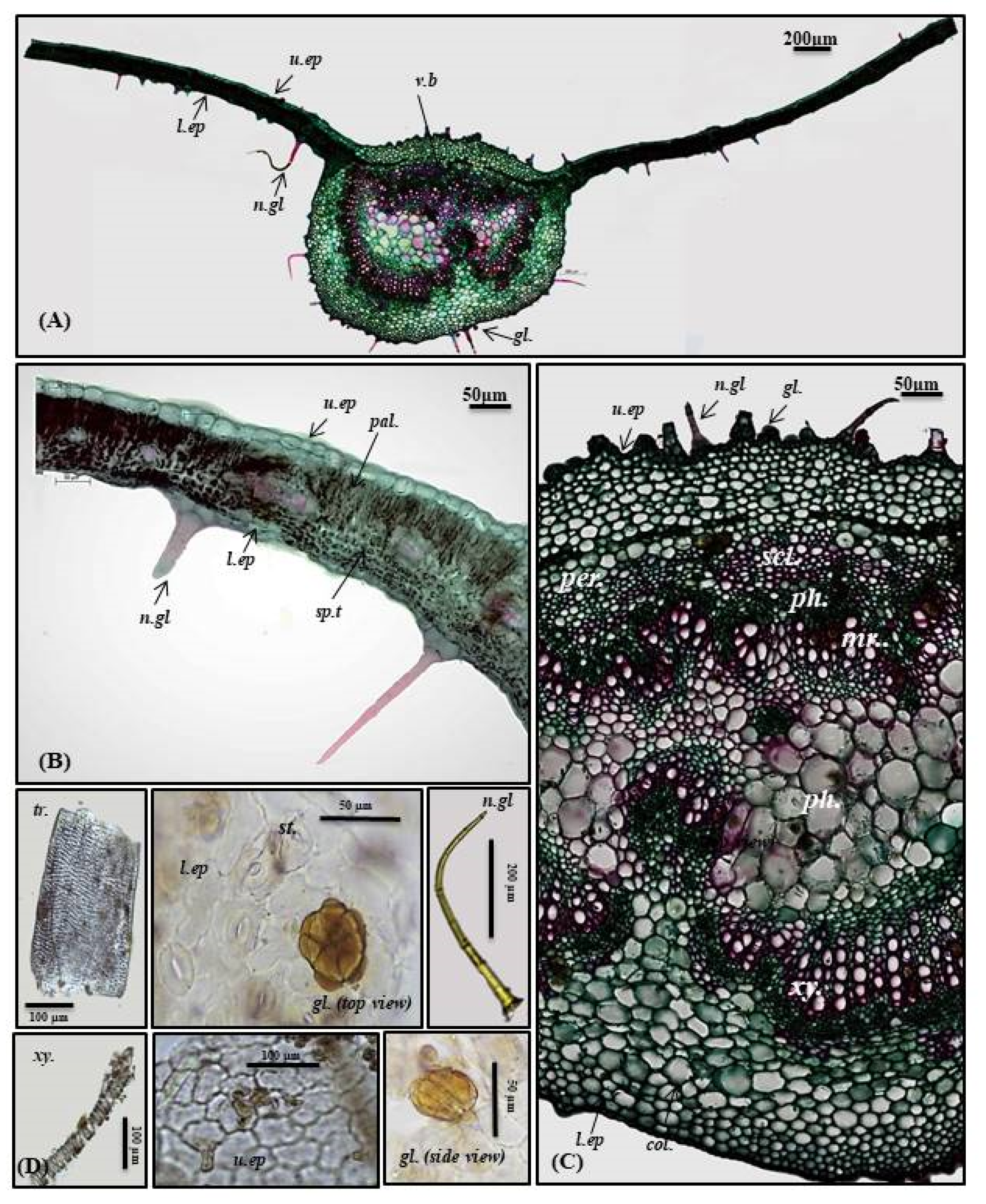
2.1.2. Micromorphological Characterization
Leaf
Petiole
Stem
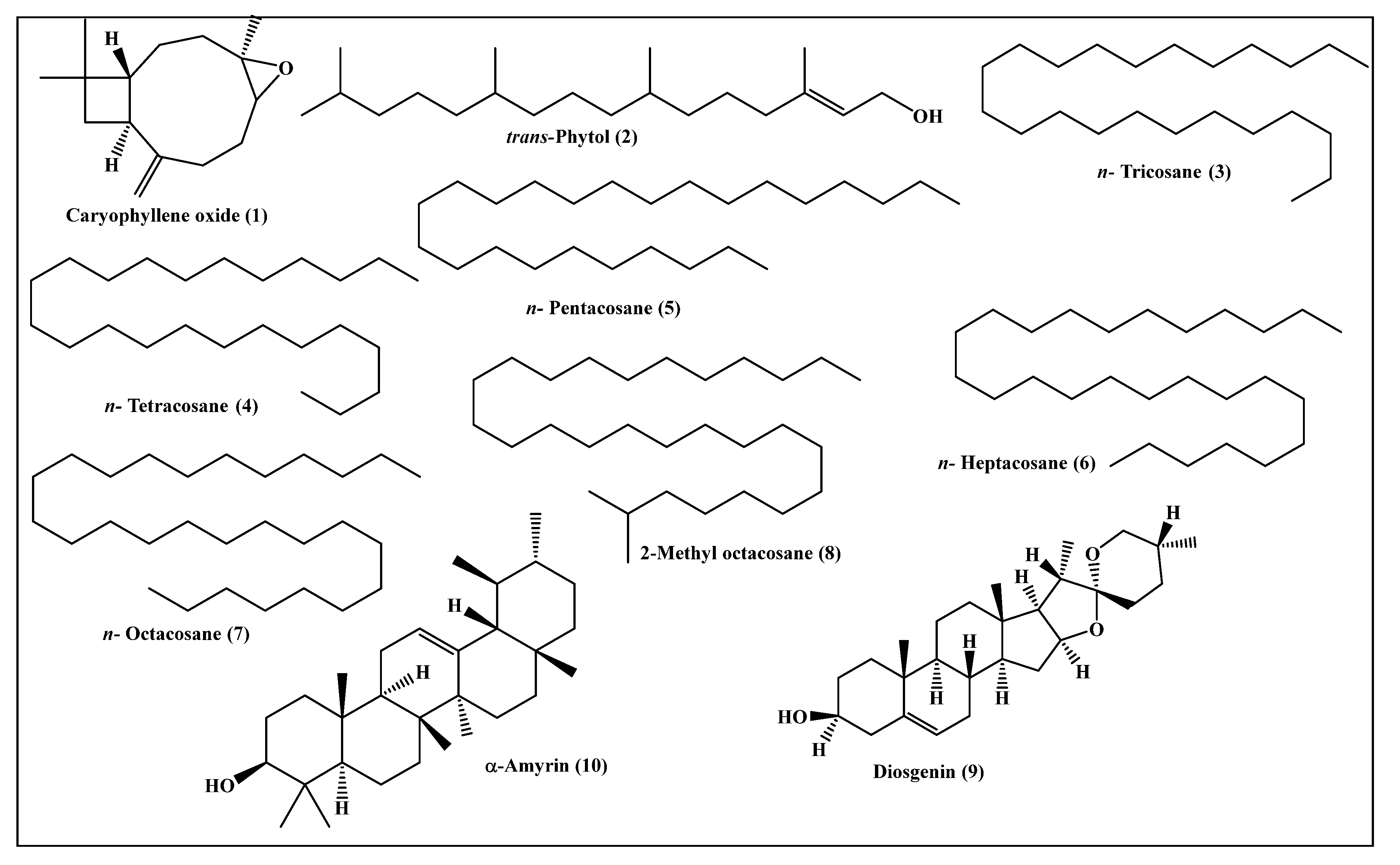
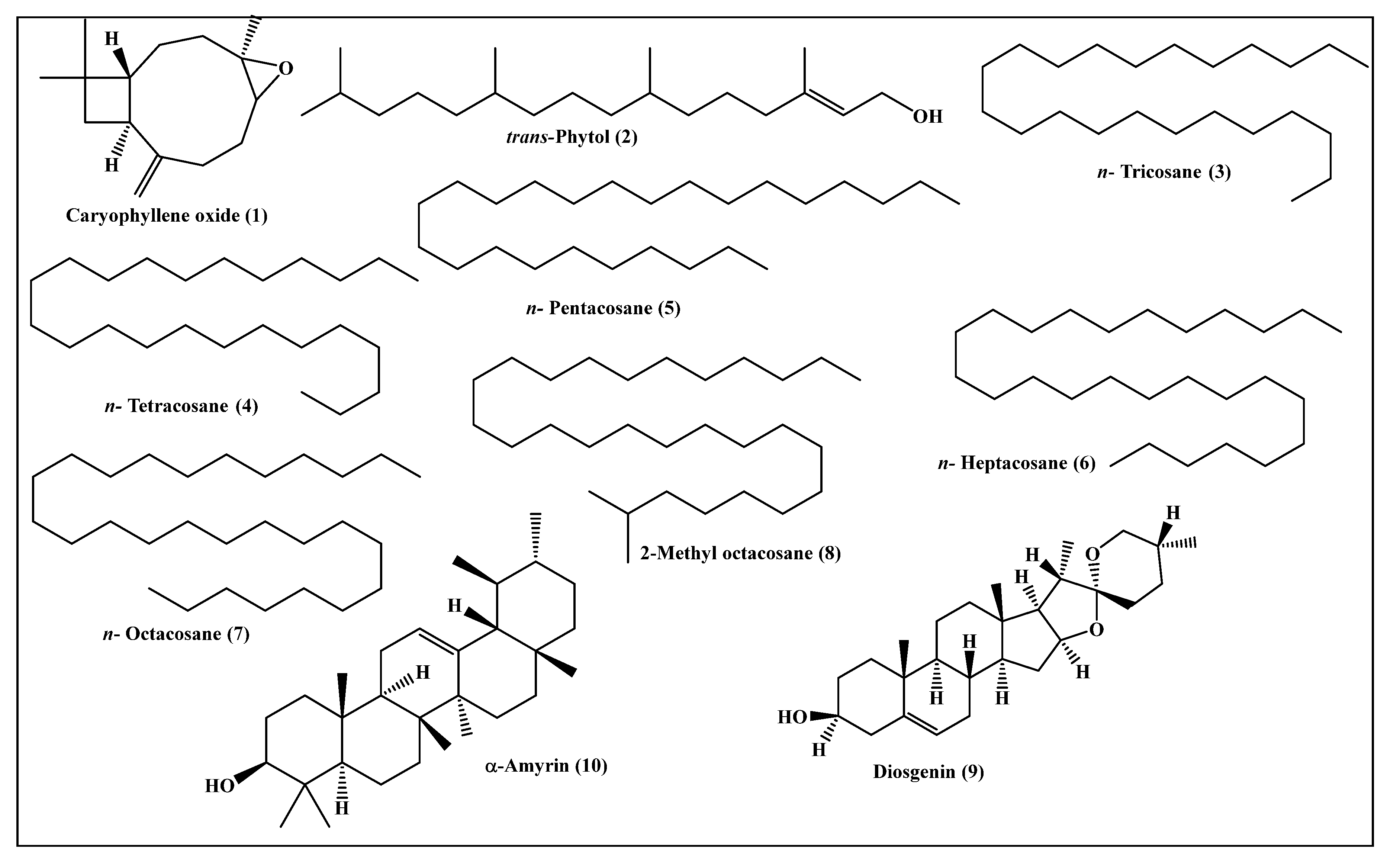
2.2. Metabolic Profiling of the Leaves n-Hexane Fraction Using GC/MS Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heading | Compound | RI | Content [%] | Identification Method | |
| Exp. | Pub. | ||||
| 1 | Caryophyllene oxide | 1576 | 1576 [10] | 7.96 | MS, RI |
| 2 | trans-Phytol | 2096 | 2101 [11] | 24.06 | MS, RI |
| 3 | n-Tricosane | 2280 | 2300 [12] | 3.15 | MS, RI |
| 4 | n-Tetracosane | 2404 | 2400 [12] | 3.20 | MS, RI |
| 5 | n-Pentacosane | 2455 | 2500 [12] | 9.69 | MS, RI |
| 6 | n-Heptacosane | 2669 | 2700 [12] | 4.49 | MS, RI |
| 7 | n-Octacosane | 2798 | 2800 [12] | 15.28 | MS, RI |
| 8 | 2-Methyl octacosane | 2854 | 2857 [13] | 2.53 | MS, RI |
| 9 | Diosgenin | 3276 | 3220 [14] | 9.28 | MS, RI |
| 10 | α-Amyrin | 3384 | 3382 [15] | 13.37 | MS, RI |
| Total identified | 93.01 | ||||
2.3. Evaluation of the Anti-Tuberculous Activity of the Leaves n-Hexane Fraction
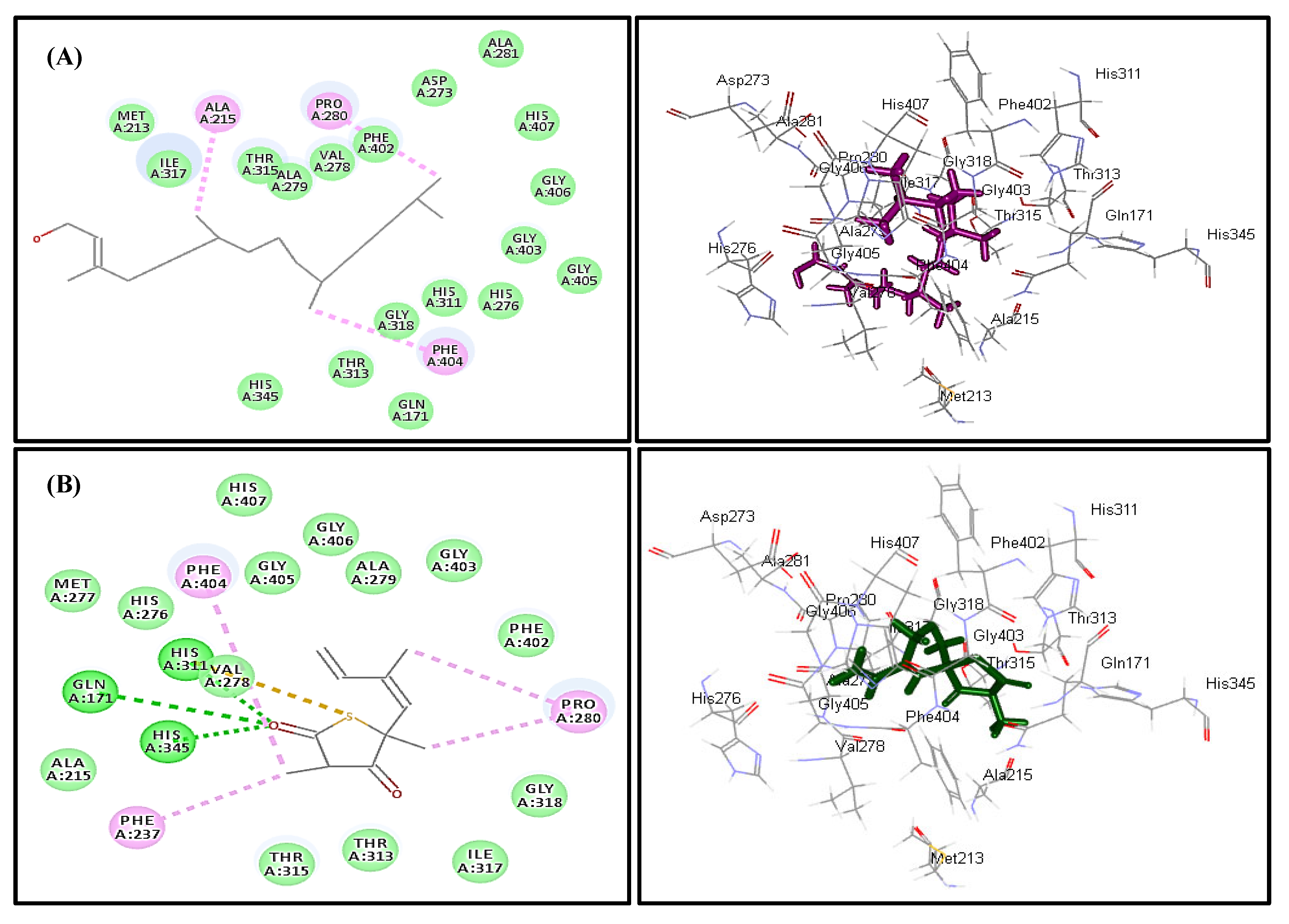
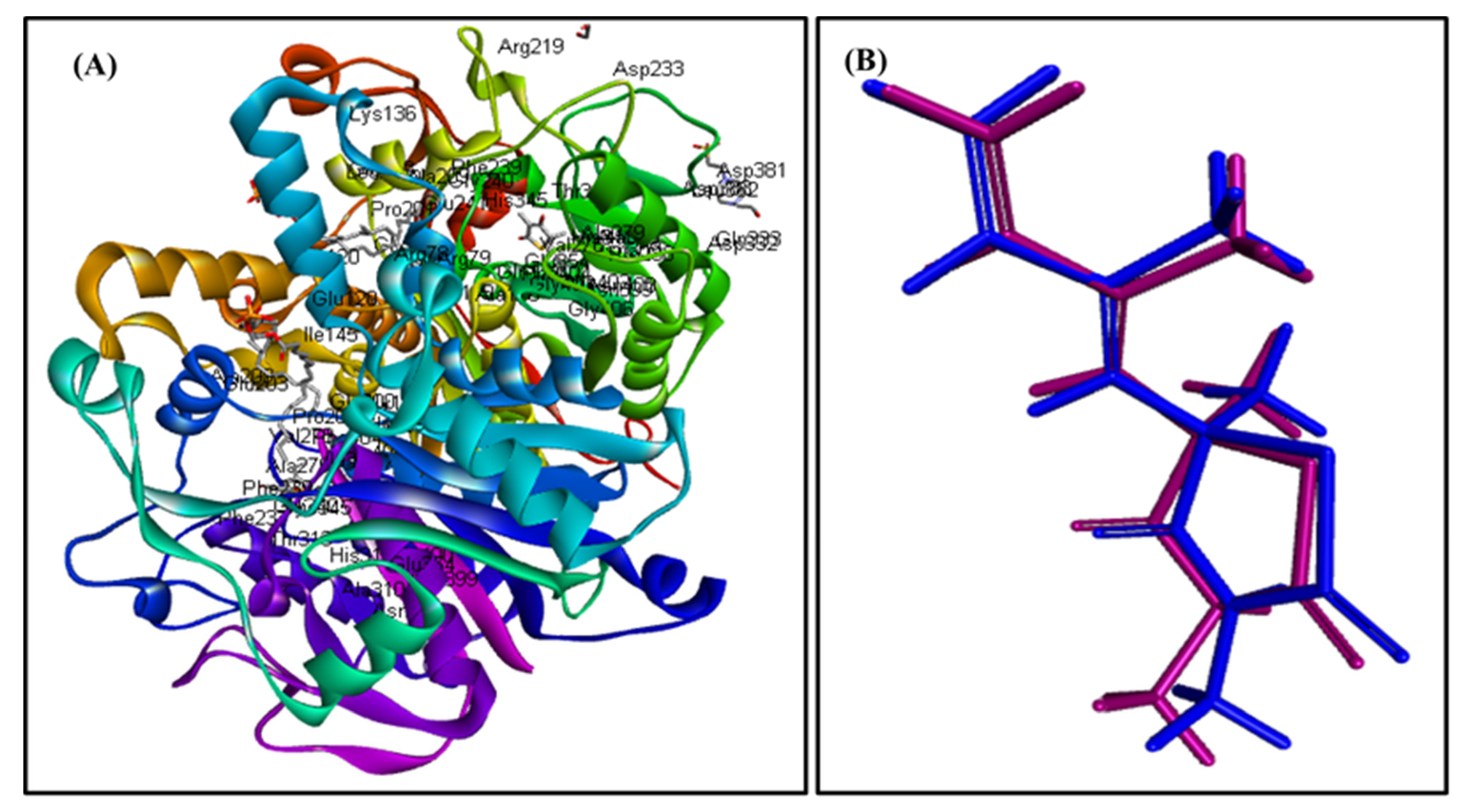
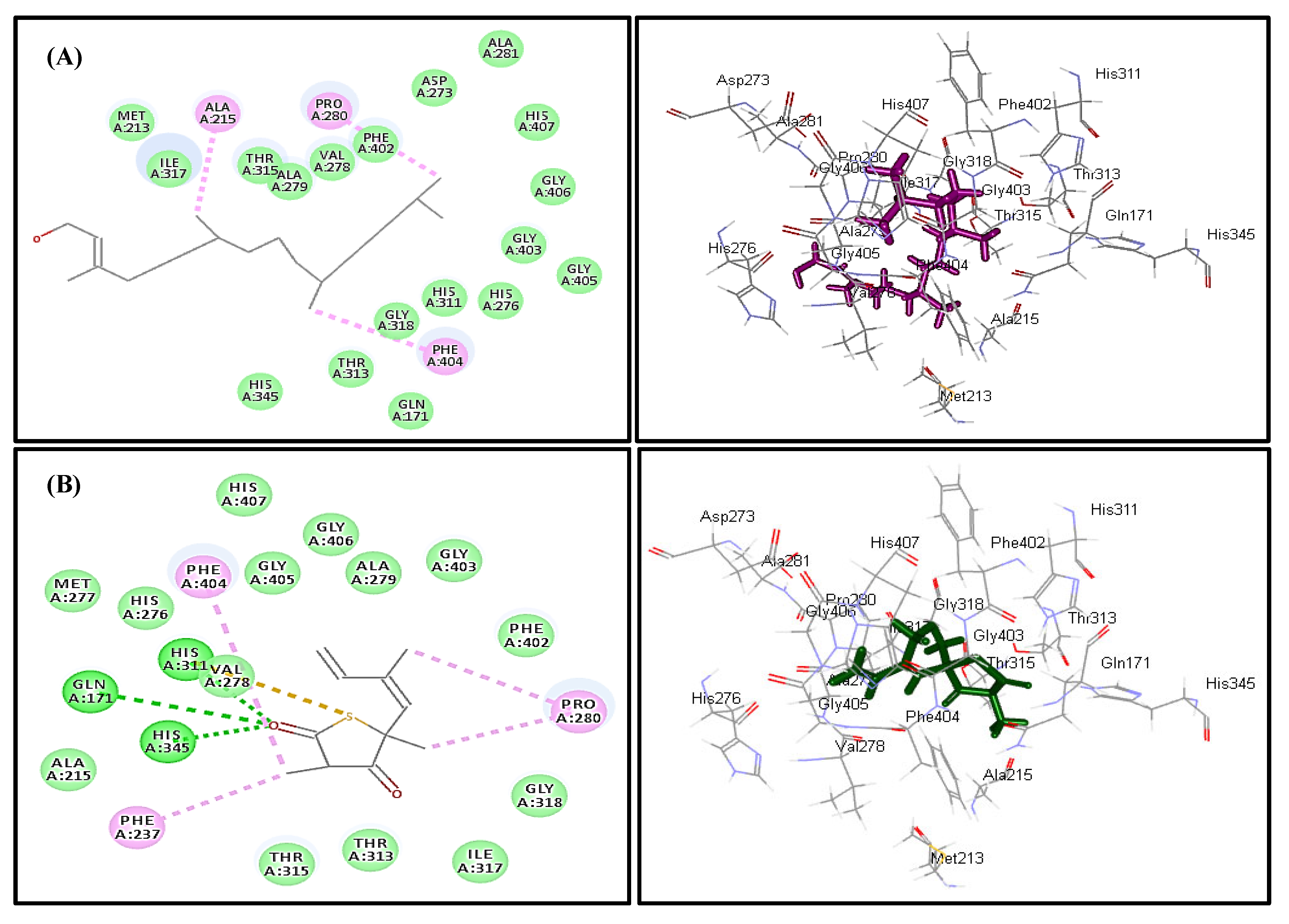
2.4. In Silico Molecular Modeling Study
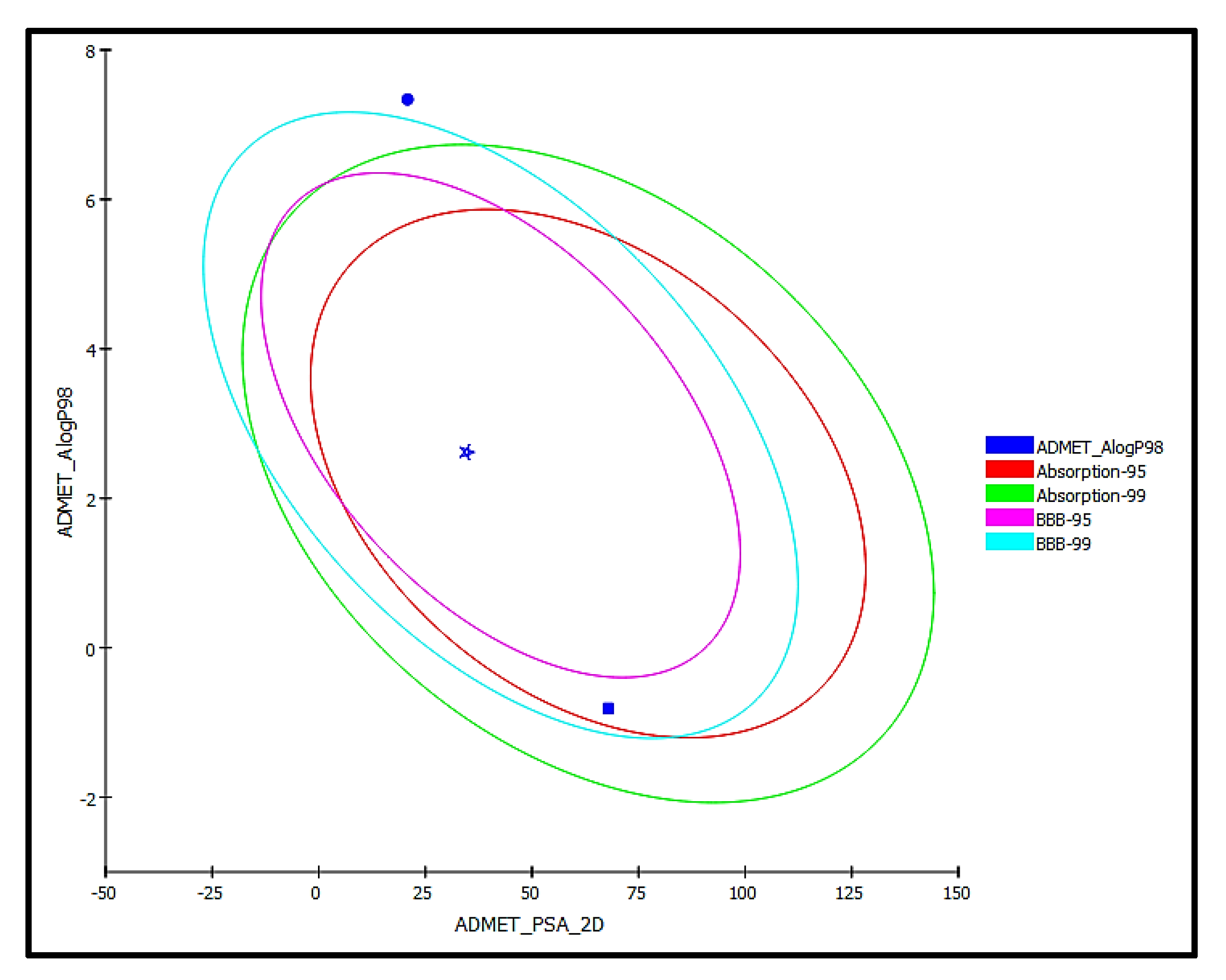
2.5. ADME/TOPKAT Prediction
3. Materials and Methods
3.1. Plant Material
3.2. Morphological and Anatomical Investigations of the Leaves, Petioles and Stems
3.3. Preparation of the Leaves n-Hexane Fraction
3.4. Metabolic Profiling of the Leaves n-Hexane Fraction Using GC/MS Analysis
3.5. Evaluation of the Anti-Tuberculous Activity of the Leaves n-Hexane Fraction
3.6. In Silico Molecular Modeling Study
3.7. ADME/TOPKAT Prediction
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munir, A.A. A taxonomic revision of the genus Premna L. (Verbenaceae) in Australia. J. Adel. Bot. Gard. 1984, 7, 1–43. [Google Scholar]
- Harley, R.; Atkins, S.; Budanstev, A.; Cantino, P.; Conn, B.; Grayer, R.; Harley, M.; De Tok, R.; Krestovskaja, T.; Morales, R. Flowering plants, dicotyledons. Fam. Genera Vasc. Plants. 2004, 6, 167–275. [Google Scholar]
- Dianita, R.; Jantan, I. Ethnomedicinal uses, phytochemistry and pharmacological aspects of the genus Premna: A review. Pharm. Biol. 2017, 55, 1715–1739. [Google Scholar] [CrossRef] [Green Version]
- de Kok, R. The genus Premna l. (Lamiaceae) in the Flora Malesiana area. Kew Bull. 2013, 68, 55–84. [Google Scholar] [CrossRef]
- Altyar, A.E.; Ashour, M.L.; Youssef, F.S. Premna odorata: Seasonal metabolic variation in the essential oil composition of its leaf and verification of its anti-ageing potential via in vitro assays and molecular modelling. Biomolecules 2020, 10, 879. [Google Scholar] [CrossRef]
- Pinzon, L.C.; Uy, M.M.; Sze, K.H.; Wang, M.; Chu, I.K. Isolation and characterization of antimicrobial, anti-inflammatory and chemopreventive flavones from Premna odorata Blanco. J. Med. Plants Res. 2011, 5, 2729–2735. [Google Scholar]
- Abdulrahaman, A.; Oladele, F. Stomata, trichomes and epidermal cells as diagnostic features in six species of genus Ocimum L. (Lamiaceae). Nig. J. Bot. 2005, 18, 214–223. [Google Scholar]
- Kowalski, R.; Kowalska, G.; Jankowska, M.; Nawrocka, A.; Kałwa, K.; Pankiewicz, U.; Włodarczyk-Stasiak, M. Secretory structures and essential oil composition of selected industrial species of Lamiaceae. Acta Sci. Pol. Hort.Cul. 2019, 18, 53–69. [Google Scholar] [CrossRef]
- Babu, K.; Dharishini, M.P.; Austin, A. Anatomical and thin layer chromatographic identification of root, root-bark, and leaf of Premna serratifolia L. Int. J. Pharmacog. 2018, 5, 302–307. [Google Scholar]
- Yusuf, M.; Begum, J.; Mondello, L.; d’Alcontres, I.S. Studies on the essential oil bearing plants of Bangladesh. Part VI. Composition of the oil of Ocimum gratissimum L. Flavour Fragr. J. 1998, 13, 163–166. [Google Scholar]
- Pino, J.A.; Marbot, R. Volatile flavor constituents of acerola (Malpighia emarginata DC.) fruit. J. Agric. Food Chem. 2001, 49, 5880–5882. [Google Scholar] [CrossRef] [PubMed]
- Korany, D.A.; Ayoub, I.M.; Labib, R.M.; El-Ahmady, S.H.; Singab, A.N.B. The impact of seasonal variation on the volatile profile of leaves and stems of Brownea grandiceps (Jacq.) with evaluation of their anti-mycobacterial and anti-inflammatory activities. South Afr. J. Bot. 2021, 142, 88–95. [Google Scholar] [CrossRef]
- Steinmetz, I.; Schmolz, E.; Ruther, J. Cuticular lipids as trail pheromone in a social wasp. Proc. R. Soc. London. Ser. B Biol. Sci. 2003, 270, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Van Gelder, W.; Jonker, H.; Huizing, H.; Scheffer, J. Capillary gas chromatography of steroidal alkaloids from solanaceae: Retention indices and simultaneous flame ionization/nitrogen-specific detection. J. Chromatogr. A 1988, 442, 133–145. [Google Scholar] [CrossRef]
- Todua, N. Retention Data. NIST Mass Spectrometry Data Center. 2011.
- Lirio, S.B.; Macabeo, A.P.; Paragas, E.M.; Knorn, M.; Kohls, P.; Franzblau, S.G.; Wang, Y.; Aguinaldo, M.A. Antitubercular constituents from Premna odorata Blanco. J. Ethnopharmacol. 2014, 154, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Elmaidomy, A.H.; Hassan, H.M.; Amin, E.; Mohamed, W.; Hetta, M.H. Premna odorata volatile oil as a new Mycobacterium tuberculosis growth inhibitor for the control of tuberculosis disease. Eur. J. Med. Plants 2017, 21, 1–11. [Google Scholar] [CrossRef]
- Waleed, A.M.; Samah, S.A.; Mona, F.S.; Abeer, H.E.; Hossam, M.H.; Elham, A.; Mona, H.H. Immunomodulatory effect of Premna odorata volatile oils in Mycobacterium tuberculosis by inhibiting TLR4/NF-κB pathway. J. Herb. Pharmacol. 2019, 8, 1–7. [Google Scholar]
- Somoskovi, A.; Parsons, L.M.; Salfinger, M. The molecular basis of resistance to isoniazid, rifampin, and pyrazinamide in Mycobacterium tuberculosis. Res. Res. 2001, 2, 1–5. [Google Scholar]
- Saludes, J.P.; Garson, M.J.; Franzblau, S.G.; Aguinaldo, A.M. Antitubercular constituents from the hexane fraction of Morinda citrifolia Linn. (Rubiaceae). Phytother. Res. 2002, 16, 683–685. [Google Scholar] [CrossRef]
- Rajab, M.S.; Cantrell, C.L.; Franzblau, S.G.; Fischer, N.H. Antimycobacterial activity of (E)-phytol and derivatives: A preliminary structure-activity study. Planta Med. 1998, 64, 2–4. [Google Scholar] [CrossRef]
- Wang, P.; Pradhan, K.; Zhong, X.-b.; Ma, X. Isoniazid metabolism and hepatotoxicity. Acta Pharm. Sinica B 2016, 6, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Bürgin, H.; Schmid, B.; Zbinden, G. Assessment of DNA damage in germ cells of male rabbits treated with isoniazid and procarbazine. Toxicology 1979, 12, 251–257. [Google Scholar] [CrossRef]
- Röhrborn, G.; Propping, P.; Buselmaier, W. Mutagenic activity of isoniazid and hydrazine in mammalian test systems. Mutat. Res.Fund. Mol. Mech. Mut. 1972, 16, 189–194. [Google Scholar] [CrossRef]
- Milad, R.; Youssef, F.S.; Ashour, M.L.; Singab, A.N. Botanical and genetic characteristics of Eremophila maculata and Eremophila purpurascens (Scrophulariaceae). World J. Pharm. Res. 2013, 2, 2711–2736. [Google Scholar]
- Youssef, F.S.; Ashour, M.L.; Wink, M. Morphological, anatomical, genetical and high performance thin layer chromatography profiling of Buddleia indica (Scrophulariaceae). Flora 2018, 246, 83–95. [Google Scholar] [CrossRef]
- Thabet, A.A.; Youssef, F.S.; Korinek, M.; Chang, F.-R.; Wu, Y.-C.; Chen, B.-H.; El-Shazly, M.; Singab, A.N.B.; Hwang, T.-L. Study of the anti-allergic and anti-inflammatory activity of Brachychiton rupestris and Brachychiton discolor leaves (Malvaceae) using in vitro models. BMC Complement. Alt. Med. 2018, 18, 299. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, I.M.; Youssef, F.S.; El-Shazly, M.; Ashour, M.L.; Singab, A.N.B.; Wink, M. Volatile constituents of Dietes bicolor (Iridaceae) and their antimicrobial activity. Z. Naturforsch. C 2015, 70, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; Hamoud, R.; Ashour, M.L.; Singab, A.N.; Wink, M. Volatile oils from the aerial parts of Eremophila maculata and their antimicrobial activity. Chem. Biodivers. 2014, 11, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Mamadalieva, N.Z.; Youssef, F.S.; Ashour, M.L.; Sasmakov, S.A.; Tiezzi, A.; Azimova, S.S. Chemical composition, antimicrobial and antioxidant activities of the essential oils of three Uzbek Lamiaceae species. Nat. Prod. Res. 2019, 33, 2394–2397. [Google Scholar] [CrossRef] [PubMed]
- Gamal El-Din, M.I.; Youssef, F.S.; Ashour, M.L.; Eldahshan, O.A.; Singab, A.N.B. Comparative Analysis of Volatile constituents of Pachira aquatica Aubl. and Pachira glabra Pasq., their anti-mycobacterial and anti-helicobacter pylori activities and their metabolic discrimination using chemometrics. J. Ess. Oil Bear. Plants 2018, 21, 1550–1567. [Google Scholar] [CrossRef]
- Youssef, F.S.; Altyar, A.E.; Omar, A.M.; Ashour, M.L. Phytoconstituents, in vitro anti-infective activity of Buddleja indica Lam., and in silico evaluation of its SARS-CoV-2 inhibitory potential. Front. Pharmacol. 2021, 12, 298. [Google Scholar] [CrossRef] [PubMed]
- Thabet, A.A.; Youssef, F.S.; El-Shazly, M.; El-Beshbishy, H.A.; Singab, A.N.B. Validation of the antihyperglycaemic and hepatoprotective activity of the flavonoid rich fraction of Brachychiton rupestris using in vivo experimental models and molecular modelling. Food Chem. Toxicol. 2018, 114, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Talaat, A.N.; Ebada, S.S.; Labib, R.M.; Esmat, A.; Youssef, F.S.; Singab, A.N.B. Verification of the anti-inflammatory activity of the polyphenolic-rich fraction of Araucaria bidwillii Hook. using phytohaemagglutinin-stimulated human peripheral blood mononuclear cells and virtual screening. J. Ethnopharmacol. 2018, 226, 44–47. [Google Scholar] [CrossRef]
- Labib, R.; Youssef, F.; Ashour, M.; Abdel-Daim, M.; Ross, S. Chemical composition of Pinus roxburghii bark volatile oil and validation of its anti-inflammatory activity using molecular modelling and bleomycin-induced inflammation in Albino mice. Molecules 2017, 22, 1384. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Durdagi, S.; Ekhteiari Salmas, R.; Macedonio, G.; Stefanucci, A.; Dimmito, M.P.; Novellino, E. Combinatorial peptide library screening for discovery of diverse α-glucosidase inhibitors using molecular dynamics simulations and binary QSAR models. J. Biomol. Str. Dyn. 2019, 37, 726–740. [Google Scholar] [CrossRef] [Green Version]


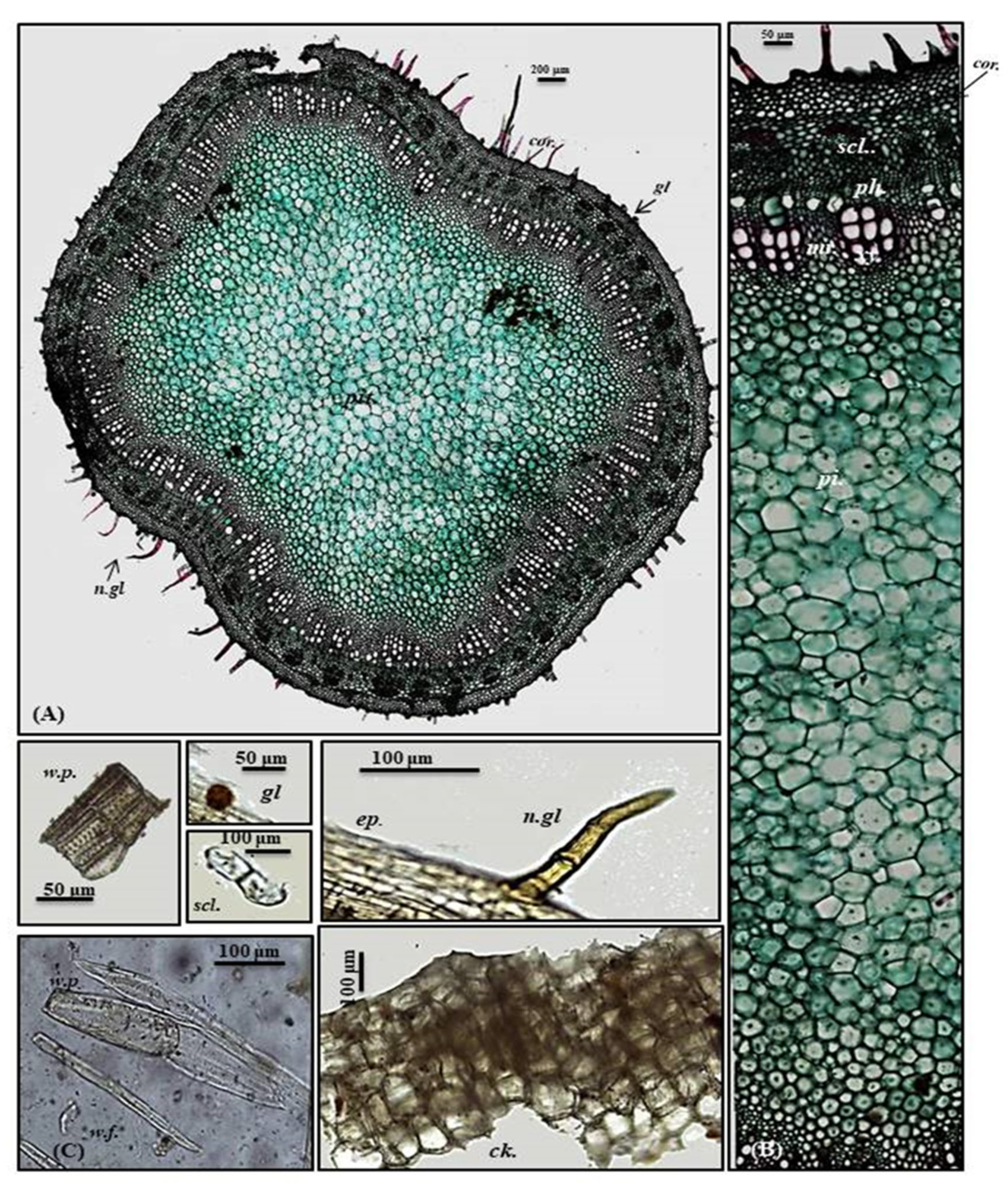

| Item | Length | Width | Height | Diameter |
|---|---|---|---|---|
| Leaf | ||||
| Upper epidermis | 71.50–66.8–62.20 | 8.80–10.20–11.00 | 6.40–10.70–15.00 | |
| Lower epidermis | 30.45–40.18–49.90 | 45.60–10.10–14.60 | 5.20–5.50–6.80 | |
| Palisade cells | 20.30–25.47–30.64 | 5.60–6.80–7.20 | 15.40–16.67–17.80 | |
| Stomata | 21.45–25.50–29.55 | 13.90–16.50–19.10 | ||
| Non-glandular trichome | 250.56–400.34–550.23 | 47.60–50.79–53.98 | ||
| Glandular (Peltate trichome) | 39.60–42.86–46.12 | 30.59–35.70–40.81 | ||
| Xylem vessels | 19.89–22.11–24.33 | |||
| Petiole | ||||
| Epidermis | 25.00–37.50–50.02 | 6.50–10.40–14.32 | 4.90–5.50–6.10 | |
| Stomata | 20.45–25.50–30.55 | 14.90–16.50–18.10 | ||
| Non-glandular trichome | 300.45–375.52–450.60 | 46.60–51.79–56.99 | ||
| Glandular (Peltate trichome) | 48.60–50.86–53.12 | 23.59–25.70–27.81 | ||
| Xylem vessels | 28.89–34.61–40.33 | |||
| Wood fibers | 350.50–375.60–400.70 | 22.59–25.30–28.00 | ||
| Pericyclic fibers | 640.44–684.70–708.95 | 7.73–10.42 -13.11 | ||
| Young and old stem branch | ||||
| Epidermis | 25.76 –30.58–35.4 | 8.14–10.70–13.26 | 4.00–4.50–5.00 | |
| Cork cells | 48.68–49.61–50.54 | 48.68–49.61–50.54 | ||
| Non-glandular trichome | 157.50–167.52–177.54 | 18.40–21.69–24.98 | ||
| Glandular (Peltate trichome) | 47.60–49.86–52.12 | 22.39–25.30–28.21 | ||
| Xylem vessels | 27.89–35.61–41.33 | |||
| Wood fibers | 320.50–375.40–430.30 | 23.59–26.30–29.00 | ||
| Wood parenchyma | 200.00–210.00–220.00 | 22.00–25.50–29.00 |
| Compound | MTB C171Q Receptor KasA Inhibitor (4C6X) | Number of Formed Hydrogen and π-Bonds with the Amino Acid Residues |
|---|---|---|
| Caryophyllene oxide (1) | 8.04 | 5; Phe404, Thr313, Ile317, Ala279, Val278 |
| trans-Phytol (2) | −15.57 | 3; Phe404, Pro280, Ala215 |
| n-Tricosane (3) | FD | - |
| n-Tetracosane (4) | FD | - |
| n-Pentacosane (5) | FD | - |
| n-Heptacosane (6) | FD | - |
| n-Octacosane (7) | FD | - |
| 2-Methyl octacosane (8) | FD | - |
| Diosgenin (9) | 173.96 | 6; Phe237, Met213, Pro280, Ala215, Phe402, His311 |
| α-Amyrin (10) | FD | - |
| Isoniazid | −21.47 | 4; Asp319, Gln322, His311, Pro280 |
| Co-crystalized ligand (Thiolactomycin) | −13.03 | 6; Gln171, His345, His311, Val278, Phe404, Pro208 |
| Compounds | trans-Phytol | Thiolactomycin | Isoniazid |
|---|---|---|---|
| ADMET parameters | |||
| Absorption Level | 3 | 0 | 0 |
| Solubility Level | 2 | 3 | 4 |
| BBB Level | 4 | 3 | 1 |
| PPB Level | True | True | False |
| CPY2D6 | NI | NI | NI |
| Hepatotoxic | Non-toxic | Non-toxic | Toxic |
| PSA-2D | 20.82 | 34.60 | 67.91 |
| Alog p98 | 7.3 | 2.62 | −0.81 |
| TOPKAT parameters | |||
| Ames prediction | Non-mutagen | Non-mutagen | Mutagen |
| Rat oral LD50 (g/kg·bw) | 9.43 | 0.20 | 0.48 |
| Rat female FDA | Non-carcinogen | Non-carcinogen | Carcinogen |
| Rat Male FDA | Non-carcinogen | Carcinogen | Non-carcinogen |
| Skin irritancy | Moderate | Moderate | None |
| Ocular irritancy | None | Mild | Mild |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, F.S.; Ovidi, E.; Musayeib, N.M.A.; Ashour, M.L. Morphology, Anatomy and Secondary Metabolites Investigations of Premna odorata Blanco and Evaluation of Its Anti-Tuberculosis Activity Using In Vitro and In Silico Studies. Plants 2021, 10, 1953. https://doi.org/10.3390/plants10091953
Youssef FS, Ovidi E, Musayeib NMA, Ashour ML. Morphology, Anatomy and Secondary Metabolites Investigations of Premna odorata Blanco and Evaluation of Its Anti-Tuberculosis Activity Using In Vitro and In Silico Studies. Plants. 2021; 10(9):1953. https://doi.org/10.3390/plants10091953
Chicago/Turabian StyleYoussef, Fadia S., Elisa Ovidi, Nawal M. Al Musayeib, and Mohamed L. Ashour. 2021. "Morphology, Anatomy and Secondary Metabolites Investigations of Premna odorata Blanco and Evaluation of Its Anti-Tuberculosis Activity Using In Vitro and In Silico Studies" Plants 10, no. 9: 1953. https://doi.org/10.3390/plants10091953
APA StyleYoussef, F. S., Ovidi, E., Musayeib, N. M. A., & Ashour, M. L. (2021). Morphology, Anatomy and Secondary Metabolites Investigations of Premna odorata Blanco and Evaluation of Its Anti-Tuberculosis Activity Using In Vitro and In Silico Studies. Plants, 10(9), 1953. https://doi.org/10.3390/plants10091953