
Competitive Ability Effects of Datura stramonium L. and Xanthium strumarium L. on the Development of Maize (Zea mays) Seeds


 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
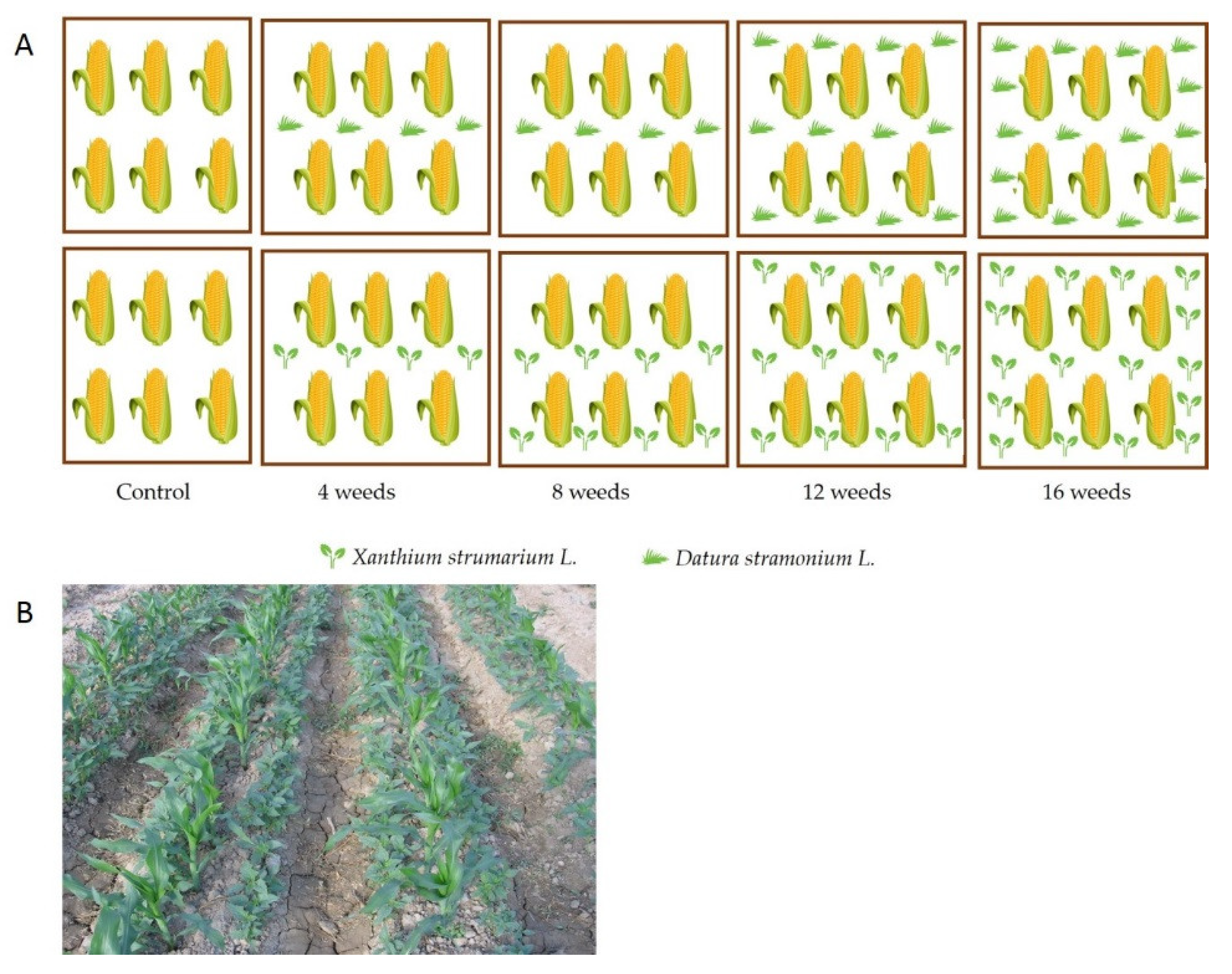
2.1. Experimental Site and Maize Seed Producing Procedures
2.2. Physical Properties Measurement
2.3. Size and Sphericity
2.4. Equivalent Diameter
2.5. Seed Volume and Surface Area
2.6. Aspect Ratio
2.7. Seeds Weight
2.8. Angle of Repose
2.9. Statistical Analysis
3. Results
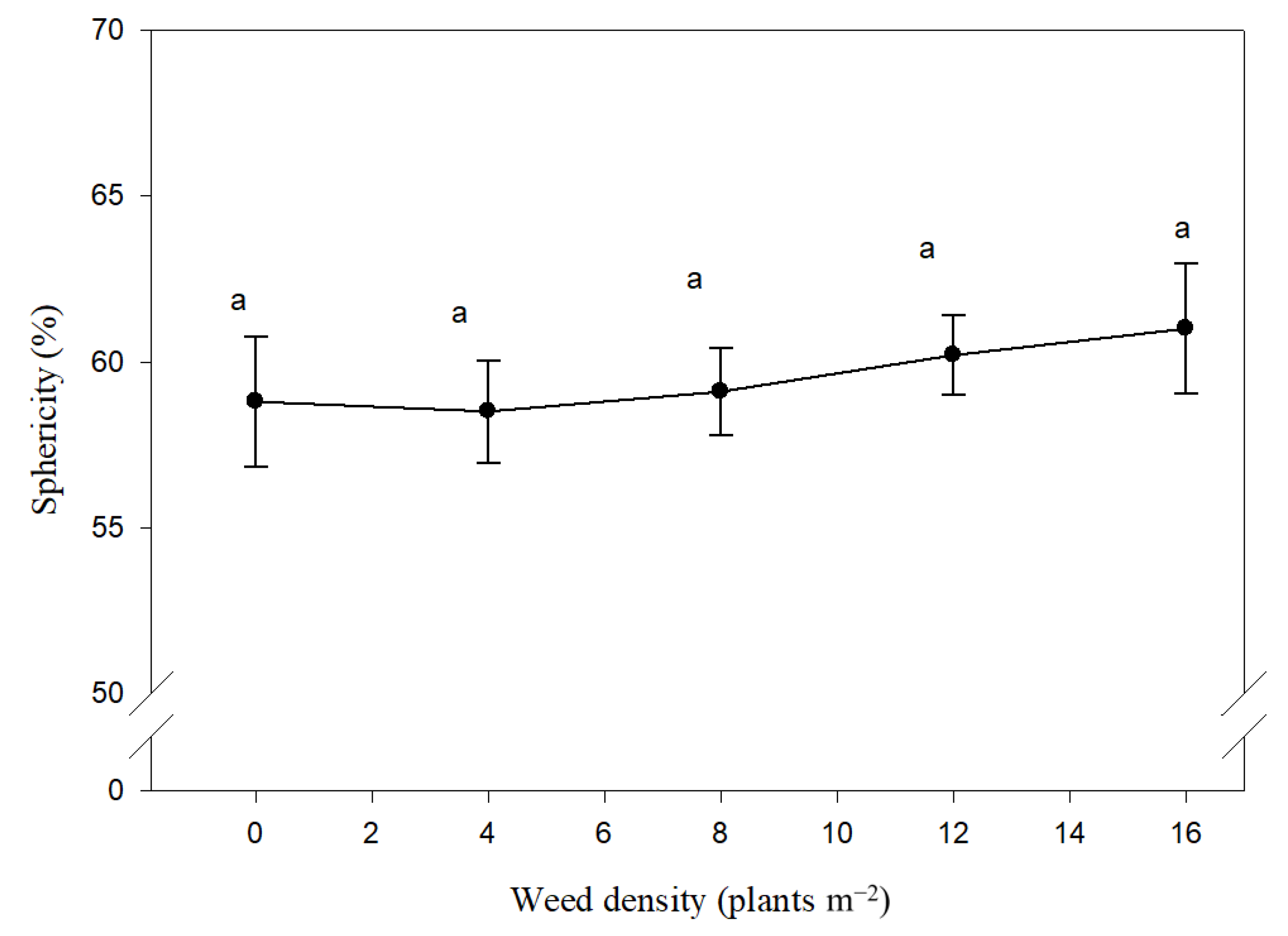
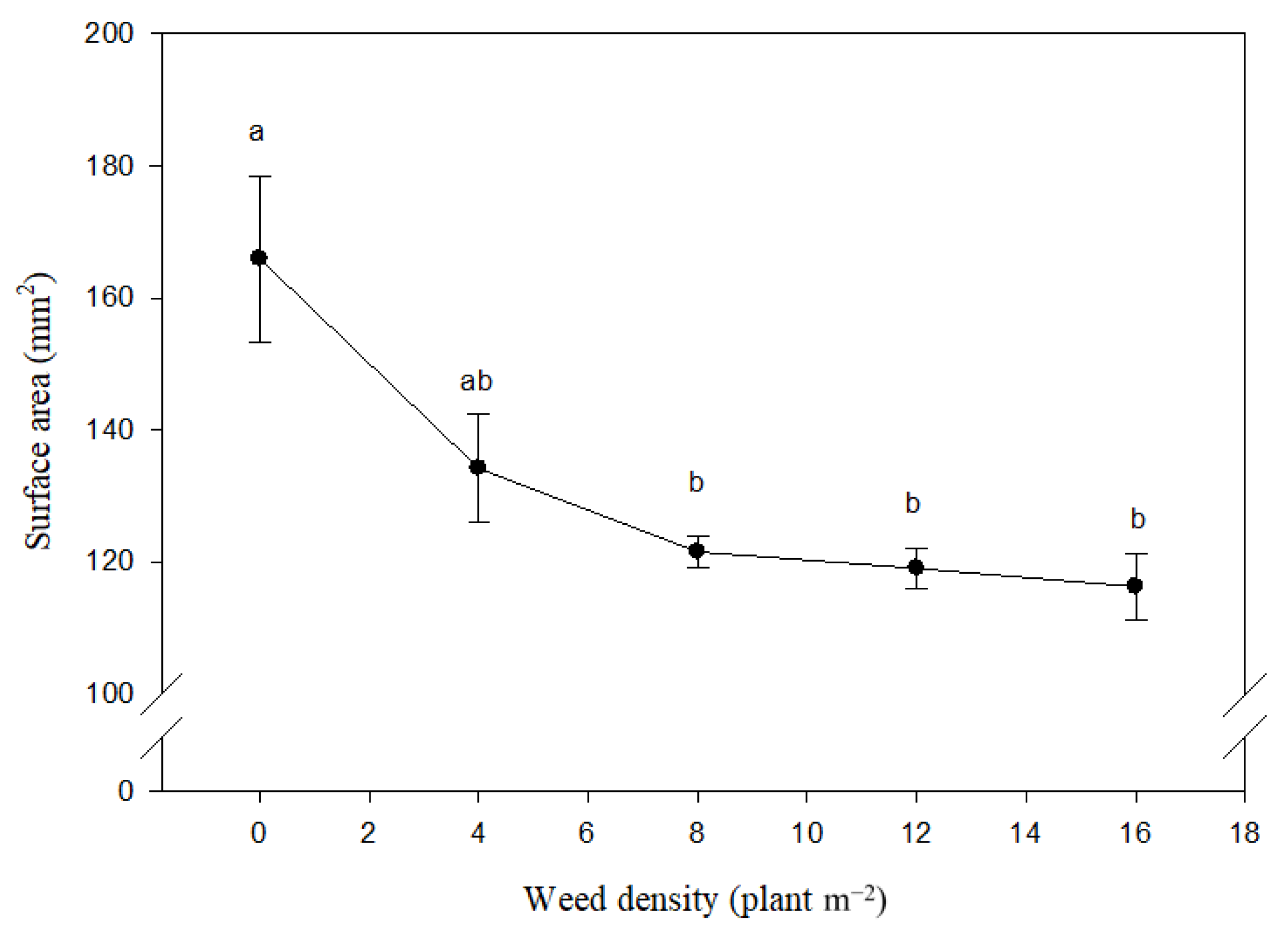
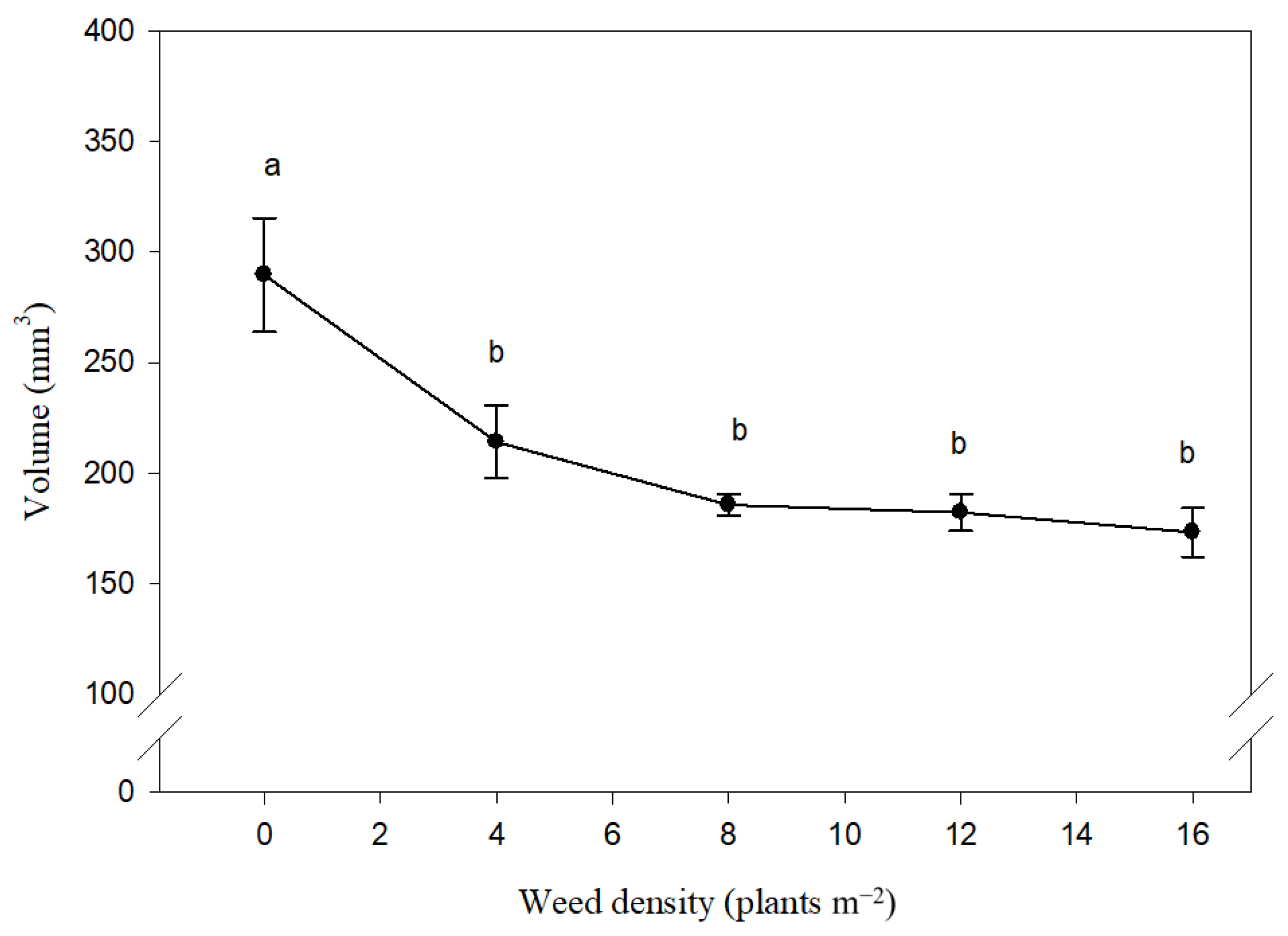
3.1. Basic Geometric Characteristics
3.2. Complex Geometrical Characteristics
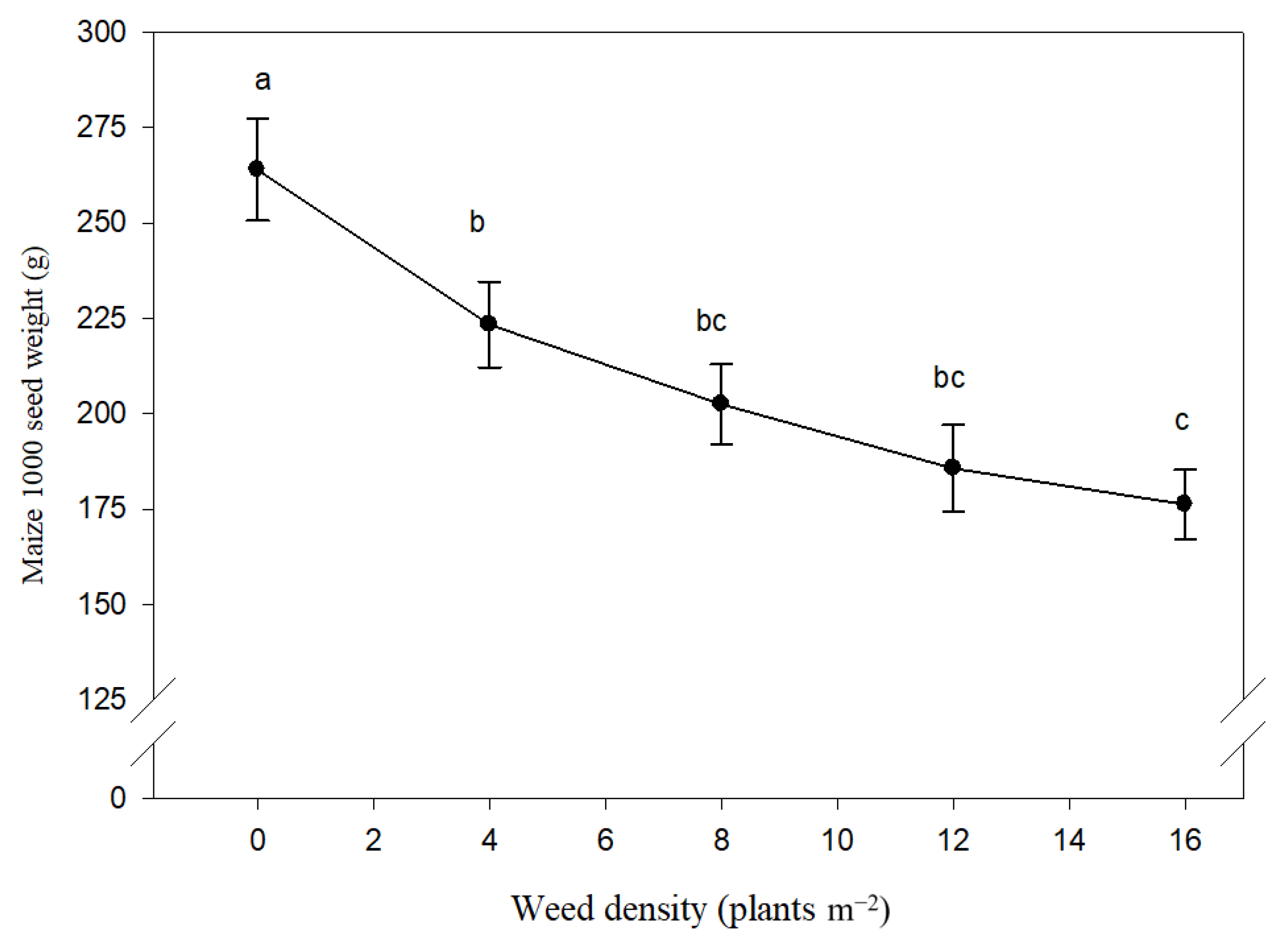
3.3. Seed Weight
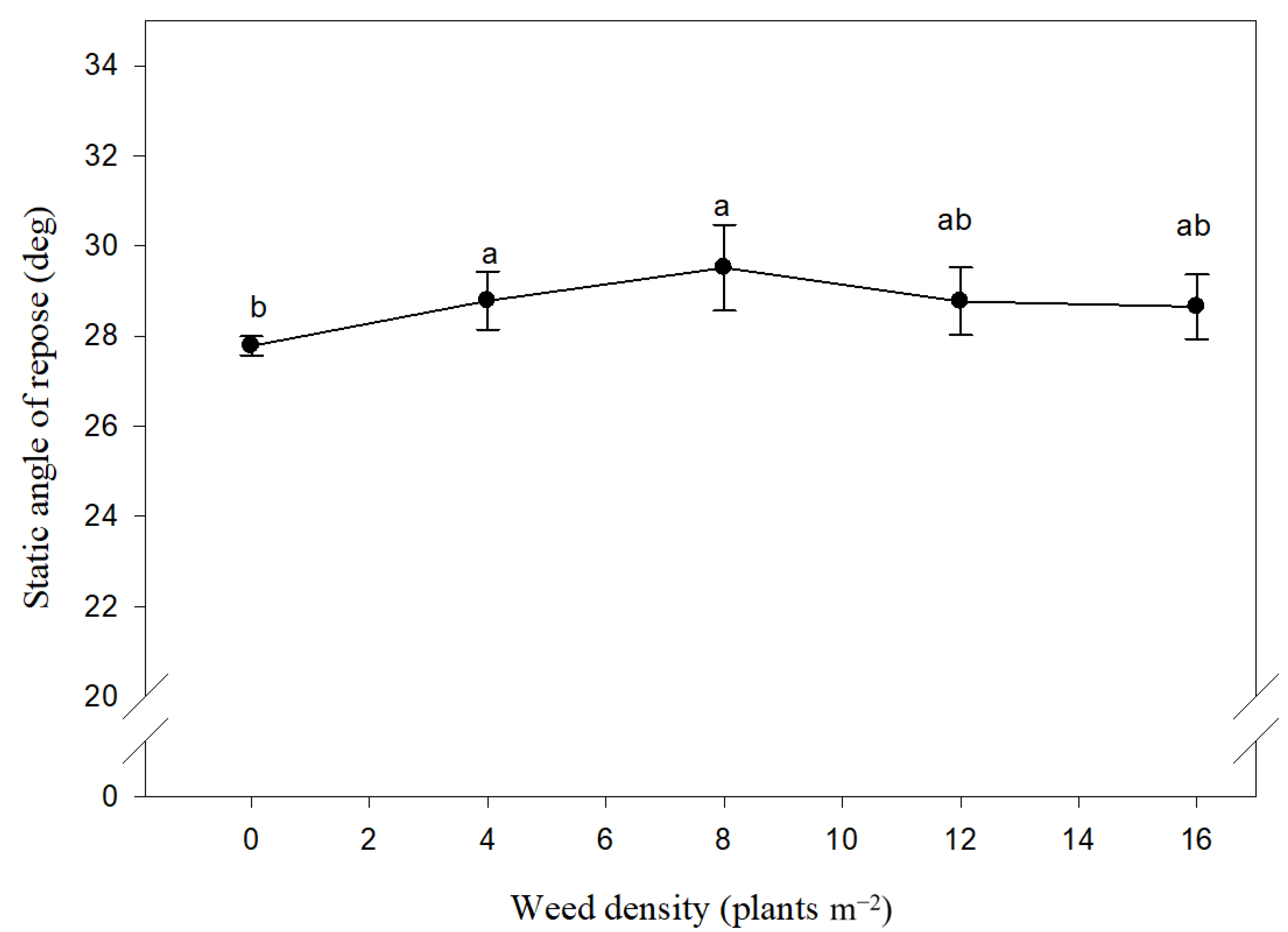
3.4. Angle of Repose
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zimdahl, R.L. Fundamentals of Weed Science, 3rd ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Boston, MA, USA, 2007; 666p. [Google Scholar]
- Kumar, A.; Memo, M.; Mastinu, A. Plant behaviour: An evolutionary response to the environment? Plant Biol. 2020, 22, 961–970. [Google Scholar] [CrossRef]
- Kumar, A.; Premoli, M.; Aria, F.; Bonini, S.A.; Maccarinelli, G.; Gianoncelli, A.; Memo, M.; Mastinu, A. Cannabimimetic plants: Are they new cannabinoidergic modulators? Planta 2019, 249, 1681–1694. [Google Scholar] [CrossRef]
- Abate, G.; Zhang, L.; Pucci, M.; Morbini, G.; Mac Sweeney, E.; Maccarinelli, G.; Ribaudo, G.; Gianoncelli, A.; Uberti, D.; Memo, M.; et al. Phytochemical Analysis and Anti-Inflammatory Activity of Different Ethanolic Phyto-Extracts of Artemisia annua L. Biomolecules 2021, 11, 975. [Google Scholar] [CrossRef]
- Gianoncelli, A.; Bonini, S.A.; Bertuzzi, M.; Guarienti, M.; Vezzoli, S.; Kumar, R.; Delbarba, A.; Mastinu, A.; Sigala, S.; Spano, P.; et al. An Integrated Approach for a Structural and Functional Evaluation of Biosimilars: Implications for Erythropoietin. BioDrugs 2015, 29, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Sanna, D.; Sanna, A.; Mara, L.; Pilichi, S.; Mastinu, A.; Chessa, F.; Pani, L.; Dattena, M. Oct4 expression in in-vitro-produced sheep blastocysts and embryonic-stem-like cells. Cell Biol. Int. 2009, 34, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Montaldo, C.; Mastinu, A.; Quartuccio, M.; Piras, V.; Denotti, G.; Pisano, E.; Orru, G. Detection and genotyping of human papillomavirus DNA in samples from healthy Sardinian patients: A preliminary study. J. Oral Pathol. Med. 2007, 36, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Aghajanlou, F.; Mirdavoudi, H.; Shojaee, M.; Mac Sweeney, E.; Mastinu, A.; Moradi, P. Rangeland Management and Ecological Adaptation Analysis Model for Astragalus curvirostris Boiss. Horticulturae 2021, 7, 67. [Google Scholar] [CrossRef]
- Moradi, P.; Aghajanloo, F.; Moosavi, A.; Minfared, H.H.; Khalafi, J.; Taghiloo, M.; Khoshzaman, T.; Shojaee, M.; Mastinu, A. Anthropic Effects on the Biodiversity of the Habitats of Ferula gummosa. Sustainability 2021, 13, 7874. [Google Scholar] [CrossRef]
- Rad, S.V.; Valadabadi, S.A.R.; Pouryousef, M.; Saifzadeh, S.; Zakrin, H.R.; Mastinu, A. Quantitative and Qualitative Evaluation of Sorghum bicolor L. under Intercropping with Legumes and Different Weed Control Methods. Horticulturae 2020, 6, 78. [Google Scholar] [CrossRef]
- Varnamkhasti, M.G.; Mobli, H.; Jafari, A.; Keyhani, A.R.; Soltanabadi, M.H.; Rafiee, S.; Kheiralipour, K. Some physical properties of rough rice (Oryza Sativa L.) grain. J. Cereal Sci. 2008, 47, 496–501. [Google Scholar] [CrossRef]
- Irtwange, S.V.; Igbeka, J.C. Some Physical Properties of Two African Yam Bean (Sphenostylis Stenocarpa) Accessions and Their Interrelations with Moisture Content. Appl. Eng. Agric. 2002, 18. [Google Scholar] [CrossRef]
- Thompson, S.A.; Ross, I.J. Compressibility and Frictional Coefficients of Wheat. Trans. ASAE 1983, 26, 1171–1176. [Google Scholar] [CrossRef]
- Gupta, A.K.; Rather, M.A.; Jha, A.K.; Shashank, A.; Singhal, S.; Sharma, M.; Pathak, U.; Sharma, D.; Mastinu, A. Artocarpus lakoocha Roxb. and Artocarpus heterophyllus Lam. Flowers: New Sources of Bioactive Compounds. Plants 2020, 9, 1329. [Google Scholar] [CrossRef]
- Mastinu, A.; Bonini, S.A.; Premoli, M.; Maccarinelli, G.; Mac Sweeney, E.; Zhang, L.L.; Lucini, L.; Memo, M. Protective Effects of Gynostemma pentaphyllum (var. Ginpent) against Lipopolysaccharide-Induced Inflammation and Motor Alteration in Mice. Molecules 2021, 26, 570. [Google Scholar] [CrossRef] [PubMed]
- Sajjan, A.S.; Dhanelappagol, M.S.; Jolli, R.B. Seed quality enhancement through seed priming in pigeonpea [Cajanus cajan (L.) Milisp.]. Legume Res. 2017, 40, 173–177. [Google Scholar] [CrossRef]
- Mhlanga, B.; Cheesman, S.; Chauhan, B.S.; Thierfelder, C. Weed emergence as affected by maize (Zea mays L.)-cover crop rotations in contrasting arable soils of Zimbabwe under conservation agriculture. Crop Prot. 2016, 81, 47–56. [Google Scholar] [CrossRef]
- Khaleghnezhad, V.; Yousefi, A.R.; Tavakoli, A.; Farajmand, B.; Mastinu, A. Concentrations-dependent effect of exogenous abscisic acid on photosynthesis, growth and phenolic content of Dracocephalum moldavica L. under drought stress. Planta 2021, 253, 127. [Google Scholar] [CrossRef] [PubMed]
- Saiedirad, M.H.; Tabatabaeefar, A.; Borghei, A.; Mirsalehi, M.; Badii, F.; Varnamkhasti, M.G. Effects of moisture content, seed size, loading rate and seed orientation on force and energy required for fracturing cumin seed (Cuminum cyminum Linn.) under quasi-static loading. J. Food Eng. 2008, 86, 565–572. [Google Scholar] [CrossRef]
- Markowski, M.; Żuk-Gołaszewska, K.; Kwiatkowski, D. Influence of variety on selected physical and mechanical properties of wheat. Ind. Crops Prod. 2013, 47, 113–117. [Google Scholar] [CrossRef]
- Sandhu, K.S.; Singh, N.; Malhi, N.S. Some properties of corn grains and their flours I: Physicochemical, functional and chapati-making properties of flours. Food Chem. 2007, 101, 938–946. [Google Scholar] [CrossRef]
- Mastinu, A.; Bonini, S.A.; Rungratanawanich, W.; Aria, F.; Marziano, M.; Maccarinelli, G.; Abate, G.; Premoli, M.; Memo, M.; Uberti, D. Gamma-oryzanol Prevents LPS-induced Brain Inflammation and Cognitive Impairment in Adult Mice. Nutrients 2019, 11, 728. [Google Scholar] [CrossRef] [PubMed]
- Rungratanawanich, W.; Cenini, G.; Mastinu, A.; Sylvester, M.; Wilkening, A.; Abate, G.; Bonini, S.A.; Aria, F.; Marziano, M.; Maccarinelli, G.; et al. γ-Oryzanol Improves Cognitive Function and Modulates Hippocampal Proteome in Mice. Nutrients 2019, 11, 753. [Google Scholar] [CrossRef] [PubMed]
- Kabas, O.; Yilmaz, E.; Ozmerzi, A.; Akinci, İ. Some physical and nutritional properties of cowpea seed (Vigna sinensis L.). J. Food Eng. 2007, 79, 1405–1409. [Google Scholar] [CrossRef]
- Amin, M.N.; Hossain, M.A.; Roy, K.C. Effects of moisture content on some physical properties of lentil seeds. J. Food Eng. 2004, 65, 83–87. [Google Scholar] [CrossRef]
- Baümler, E.; Cuniberti, A.; Nolasco, S.M.; Riccobene, I.C. Moisture dependent physical and compression properties of safflower seed. J. Food Eng. 2006, 72, 134–140. [Google Scholar] [CrossRef]
- Goyal, R.K.; Patil, R.T.; Kingsly, A.R.P.; Walia, H.; Kumar, P. Status of Post harvest Technology of Aonla in India-A Review. Am. J. Food Technol. 2007, 3, 13–23. [Google Scholar] [CrossRef][Green Version]
- Omobuwajo, T.O.; Omobuwajo, O.R.; Sanni, L.A. Physical properties of calabash nutmeg (Monodora myristica) seeds. J. Food Eng. 2003, 57, 375–381. [Google Scholar] [CrossRef]
- Konak, M.; Çarman, K.; Aydin, C. PH—Postharvest Technology. Biosyst. Eng. 2002, 82, 73–78. [Google Scholar] [CrossRef]
- Mahdavi, A.; Moradi, P.; Mastinu, A. Variation in Terpene Profiles of Thymus vulgaris in Water Deficit Stress Response. Molecules 2020, 25, 1091. [Google Scholar] [CrossRef]
- Naservafaei, S.; Sohrabi, Y.; Moradi, P.; Mac Sweeney, E.; Mastinu, A. Biological Response of Lallemantia iberica to Brassinolide Treatment under Different Watering Conditions. Plants 2021, 10, 496. [Google Scholar] [CrossRef]
- Yousefi, A.R.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and Seedling Growth Responses of Zygophyllum fabago, Salsola kali L. and Atriplex canescens to PEG-Induced Drought Stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Zangani, E.; Afsahi, K.; Shekari, F.; Mac Sweeney, E.; Mastinu, A. Nitrogen and Phosphorus Addition to Soil Improves Seed Yield, Foliar Stomatal Conductance, and the Photosynthetic Response of Rapeseed (Brassica napus L.). Agriculture 2021, 11, 483. [Google Scholar] [CrossRef]
- Mastinu, A.; Ascrizzi, R.; Ribaudo, G.; Bonini, S.A.; Premoli, M.; Aria, F.; Maccarinelli, G.; Gianoncelli, A.; Flamini, G.; Pistelli, L.; et al. Prosocial Effects of Nonpsychotropic Cannabis sativa in Mice. Cannabis Cannabinoid Res. 2021. [Google Scholar] [CrossRef]
- Zaragoza, C.; Pardo, S. Competition between maize andDatura stramoniumin an irrigated field under semi-arid conditions. Weed Res. 2002, 39, 225–240. [Google Scholar] [CrossRef]
- Deng, F. Effects of glyphosate, chlorsulfuron, and methyl jasmonate on growth and alkaloid biosynthesis of jimsonweed (Datura stramonium L.). Pestic. Biochem. Physiol. 2005, 82, 16–26. [Google Scholar] [CrossRef]
- Wassom, J.J.; Knepp, A.W.; Tranel, P.J.; Wax, L.M. Variability in photosynthetic rates and accumulated biomass among greenhouse-grown common cocklebur (Xanthium strumarium) accessions. Weed Technol. 2003, 17, 84–88. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Oliveira, M.J. Light and Temperature Requirements for Common Cocklebur (Xanthium strumarium) Germination During After-Ripening under Field Conditions. Weed Sci. 2007, 55, 227–234. [Google Scholar] [CrossRef]
- Webster, T.M.; Davis, R.F. Southern Root-Knot Nematode (Meloidogyne incognita) Affects Common Cocklebur (Xanthium strumarium) Interference with Cotton. Weed Sci. 2017, 55, 143–146. [Google Scholar] [CrossRef]
- Westbrook, A.S.; Han, R.; Zhu, J.; Cordeau, S.; DiTommaso, A. Drought and Competition With Ivyleaf Morningglory (Ipomoea hederacea) Inhibit Corn and Soybean Growth. Front. Agron. 2021, 3. [Google Scholar] [CrossRef]
- Yousefi, A.R.; Gonzalez-Andujar, J.L.; Alizadeh, H.; Baghestani, M.A.; Rahimian Mashhadi, H.; Karimmojeni, H. Interactions between reduced rate of imazethapyr and multiple weed species-soyabean interference in a semi-arid environment. Weed Res. 2012, 52, 242–251. [Google Scholar] [CrossRef]
- Travlos, I.; Gazoulis, I.; Kanatas, P.; Tsekoura, A.; Zannopoulos, S.; Papastylianou, P. Key Factors Affecting Weed Seeds’ Germination, Weed Emergence, and Their Possible Role for the Efficacy of False Seedbed Technique as Weed Management Practice. Front. Agron. 2020, 2. [Google Scholar] [CrossRef]
- Schut, M.; Rodenburg, J.; Klerkx, L.; Kayeke, J.; van Ast, A.; Bastiaans, L. RAAIS: Rapid Appraisal of Agricultural Innovation Systems (Part II). Integrated analysis of parasitic weed problems in rice in Tanzania. Agric. Syst. 2015, 132, 12–24. [Google Scholar] [CrossRef]
- Mohsenin, N.N. Physical Properties of Plant and Animal Materials: Structure, Physical Characteristics, and Mechanical Properties, 2nd ed.; Gordon and Breach: New York, NY, USA, 1986. [Google Scholar]
- Bande, Y.M.; Adam, N.M.; Azmi, Y.; Jamarei, O. Moisture-dependent Physical and Compression Properties of Bitter Melon (Citrullus colocynthis lanatus) Seeds. Int. J. Agric. Res. 2012, 7, 243–254. [Google Scholar] [CrossRef][Green Version]
- Cerrudo, D.; Page, E.R.; Tollenaar, M.; Stewart, G.; Swanton, C.J. Mechanisms of Yield Loss in Maize Caused by Weed Competition. Weed Sci. 2017, 60, 225–232. [Google Scholar] [CrossRef]
- Steinmaus, S.J.; Norris, R.F. Growth analysis and canopy architecture of velvetleaf grown under light conditions representative of irrigated Mediterranean-type agroecosystems. Weed Sci. 2002, 50, 42–53. [Google Scholar] [CrossRef]
- Gou, F.; van Ittersum, M.K.; Simon, E.; Leffelaar, P.A.; van der Putten, P.E.L.; Zhang, L.; van der Werf, W. Intercropping wheat and maize increases total radiation interception and wheat RUE but lowers maize RUE. Eur. J. Agron. 2017, 84, 125–139. [Google Scholar] [CrossRef]
- Su, Y.; Cui, T.; Zhang, D.; Xia, G.; Gao, X.; He, X.; Xu, Y. Effects of shape feature on compression characteristics and crack rules of maize kernel. J. Food Process. Preserv. 2019, 44. [Google Scholar] [CrossRef]
- Wandscheer, A.C.D.; Rizzardi, M.A. Interference of soybean and corn with Chloris distichophylla. Ciênc. Agrotecnologia 2013, 37, 306–312. [Google Scholar] [CrossRef][Green Version]
- Karimmojeni, H.; Rahimian Mashhadi, H.; Alizadeh, H.M.; Cousens, R.D.; Beheshtian Mesgaran, M. Interference between maize and Xanthium strumarium or Datura stramonium. Weed Res. 2010, 50, 253–261. [Google Scholar] [CrossRef]
- Massinga, R.A.; Currie, R.S.; Horak, M.J.; Boyer, J. Interference of Palmer amaranth in corn. Weed Sci. 2001, 49, 202–208. [Google Scholar] [CrossRef]
- Deng, J.; Zuo, W.; Wang, Z.; Fan, Z.; Ji, M.; Wang, G.; Ran, J.; Zhao, C.; Liu, J.; Niklas, K.J.; et al. Insights into plant size-density relationships from models and agricultural crops. Proc. Natl. Acad. Sci. USA 2012, 109, 8600–8605. [Google Scholar] [CrossRef]
- Dhima, K.V.; Eleftherohorinos, I.G.; Vasilakoglou, I.B. Interference between Avena sterilis, Phalaris minor and five barley cultivars. Weed Res. 2000, 40, 549–559. [Google Scholar] [CrossRef]
- IŞIk, E.; ÜNal, H. Moisture-dependent physical properties of white speckled red kidney bean grains. J. Food Eng. 2007, 82, 209–216. [Google Scholar] [CrossRef]
- Soltani, N.; Dille, J.A.; Burke, I.C.; Everman, W.J.; VanGessel, M.J.; Davis, V.M.; Sikkema, P.H. Potential Corn Yield Losses from Weeds in North America. Weed Technol. 2017, 30, 979–984. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Oliveira, M.J. Comparison of the critical period for weed control in wide- and narrow-row corn. Weed Sci. 2004, 52, 802–807. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Cavigelli, M.A. Subplots facilitate assessment of corn yield losses from weed competition in a long-term systems experiment. Agron. Sustain. Dev. 2010, 30, 445–453. [Google Scholar] [CrossRef]
- Nikolić, N.; Rizzo, D.; Marraccini, E.; Ayerdi Gotor, A.; Mattivi, P.; Saulet, P.; Persichetti, A.; Masin, R. Site and time-specific early weed control is able to reduce herbicide use in maize—A case study. Ital. J. Agron. 2021. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.O.V | df | Length | Width | Thickness | Aspect Ratio | Surface Area | Volume | Sphericity | Equivalent Diameter | Angle of Repose | Seed Weight |
|---|---|---|---|---|---|---|---|---|---|---|---|
| p-Value | |||||||||||
| Year (Y) | 1 | 0.0001 | 0.32 | 0.19 | 0.01 | 0.15 | 0.12 | 0.53 | 0.11 | 0.71 | 0.06 |
| Block [Y] | 4 | 0.60 | 0.08 | 0.12 | 0.91 | 0.01 | 0.00 | 0.13 | 0.00 | 0.40 | 0.27 |
| Density (D) | 4 | 0.00 | 0.00 | 0.12 | 0.02 | 0.01 | 0.00 | 0.42 | 0.00 | 0.01 | 0.00 |
| Species (S) | 1 | 0.49 | 0.35 | 0.44 | 0.40 | 0.87 | 0.81 | 0.24 | 0.85 | 0.13 | 0.32 |
| D × S | 4 | 0.22 | 0.52 | 0.80 | 0.16 | 0.92 | 0.89 | 0.14 | 0.91 | 0.22 | 0.44 |
| Y × D | 4 | 0.58 | 0.10 | 0.13 | 0.58 | 0.15 | 0.10 | 0.21 | 0.17 | 0.92 | 0.38 |
| Y × S | 1 | 0.18 | 0.35 | 0.46 | 0.12 | 0.33 | 0.34 | 0.37 | 0.33 | 0.83 | 0.09 |
| S × D × Y | 4 | 0.17 | 0.89 | 0.36 | 0.51 | 0.43 | 0.48 | 0.68 | 0.35 | 0.45 | 0.24 |
| Error | 36 | - | - | - | - | - | - | - | - | - | - |
| Density | Length (mm) | Width (mm) | Thickness (mm) | Aspect Ratio | Equivalent Diameter (mm) |
|---|---|---|---|---|---|
| 0 | 13.44 a | 8.33 a | 4.45 a | 61.83 b | 8.19 a |
| 4 | 12.16 b | 7.73 b | 3.84 a | 63.66 ab | 7.41 b |
| 8 | 11.47 bc | 7.50 b | 3.62 a | 65.50 ab | 7.07 b |
| 12 | 11.14 c | 7.59 b | 3.62 a | 68.33 a | 7.03 b |
| 16 | 10.8 c | 7.39 b | 3.57 a | 68.16 a | 6.9 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karimmojeni, H.; Rahimian, H.; Alizadeh, H.; Yousefi, A.R.; Gonzalez-Andujar, J.L.; Sweeney, E.M.; Mastinu, A. Competitive Ability Effects of Datura stramonium L. and Xanthium strumarium L. on the Development of Maize (Zea mays) Seeds. Plants 2021, 10, 1922. https://doi.org/10.3390/plants10091922
Karimmojeni H, Rahimian H, Alizadeh H, Yousefi AR, Gonzalez-Andujar JL, Sweeney EM, Mastinu A. Competitive Ability Effects of Datura stramonium L. and Xanthium strumarium L. on the Development of Maize (Zea mays) Seeds. Plants. 2021; 10(9):1922. https://doi.org/10.3390/plants10091922
Chicago/Turabian StyleKarimmojeni, Hassan, Hamid Rahimian, Hassan Alizadeh, Ali Reza Yousefi, Jose L. Gonzalez-Andujar, Eileen Mac Sweeney, and Andrea Mastinu. 2021. "Competitive Ability Effects of Datura stramonium L. and Xanthium strumarium L. on the Development of Maize (Zea mays) Seeds" Plants 10, no. 9: 1922. https://doi.org/10.3390/plants10091922
APA StyleKarimmojeni, H., Rahimian, H., Alizadeh, H., Yousefi, A. R., Gonzalez-Andujar, J. L., Sweeney, E. M., & Mastinu, A. (2021). Competitive Ability Effects of Datura stramonium L. and Xanthium strumarium L. on the Development of Maize (Zea mays) Seeds. Plants, 10(9), 1922. https://doi.org/10.3390/plants10091922