
Investigation of Chemical Profiles of Different Parts of Morus alba Using a Combination of Molecular Networking Methods with Mass Spectral Data from Two Ionization Modes of LC/MS


Abstract
1. Introduction
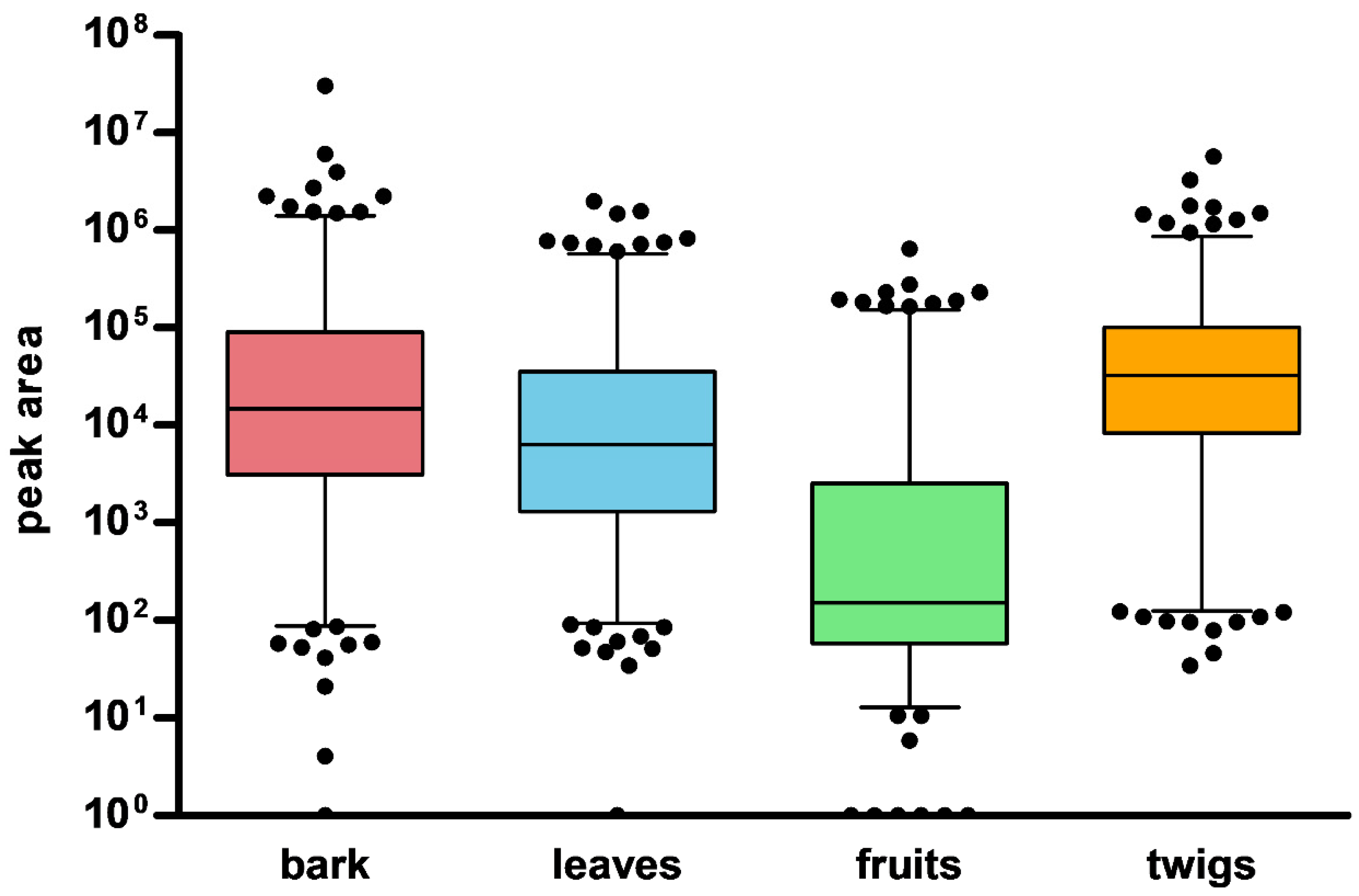
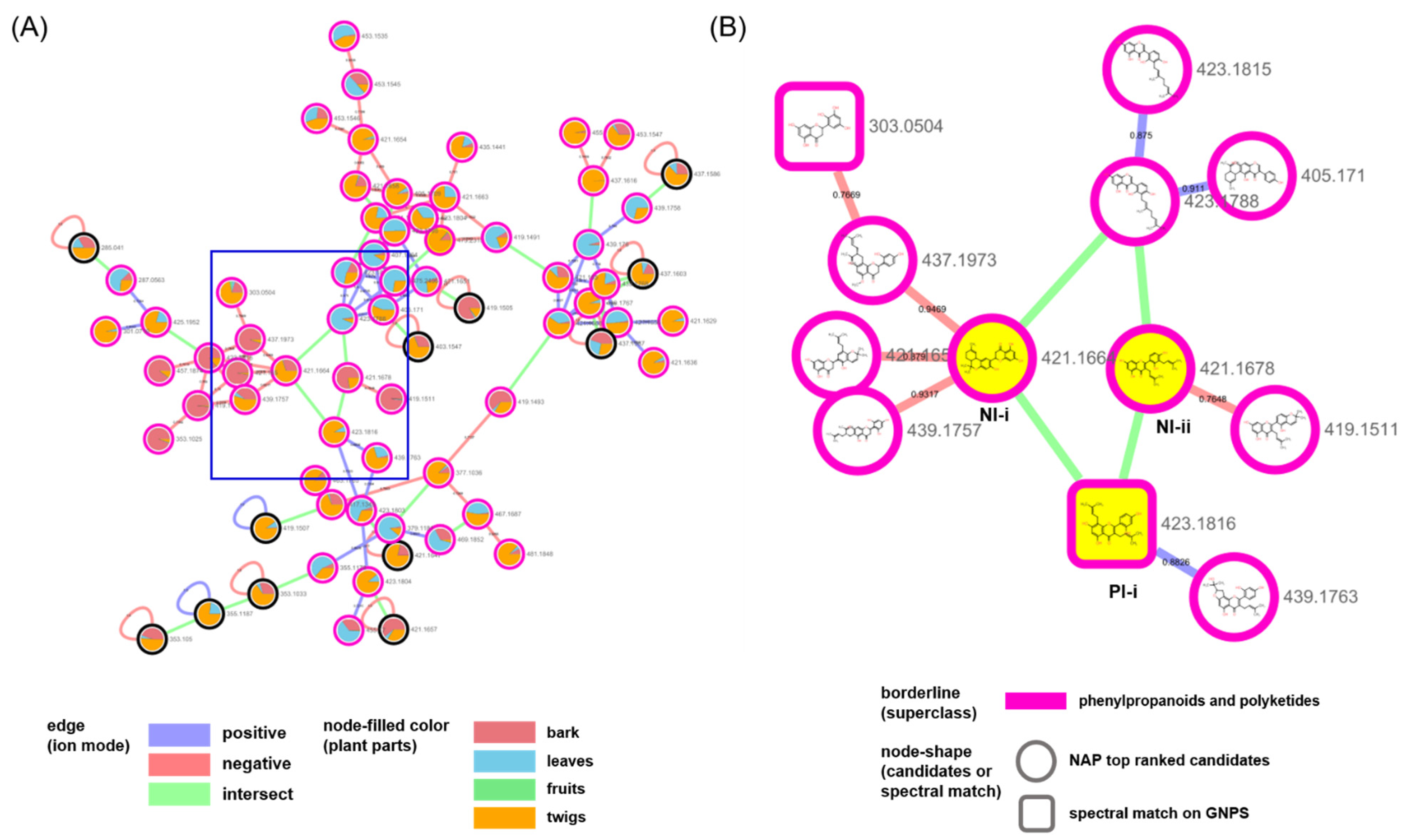
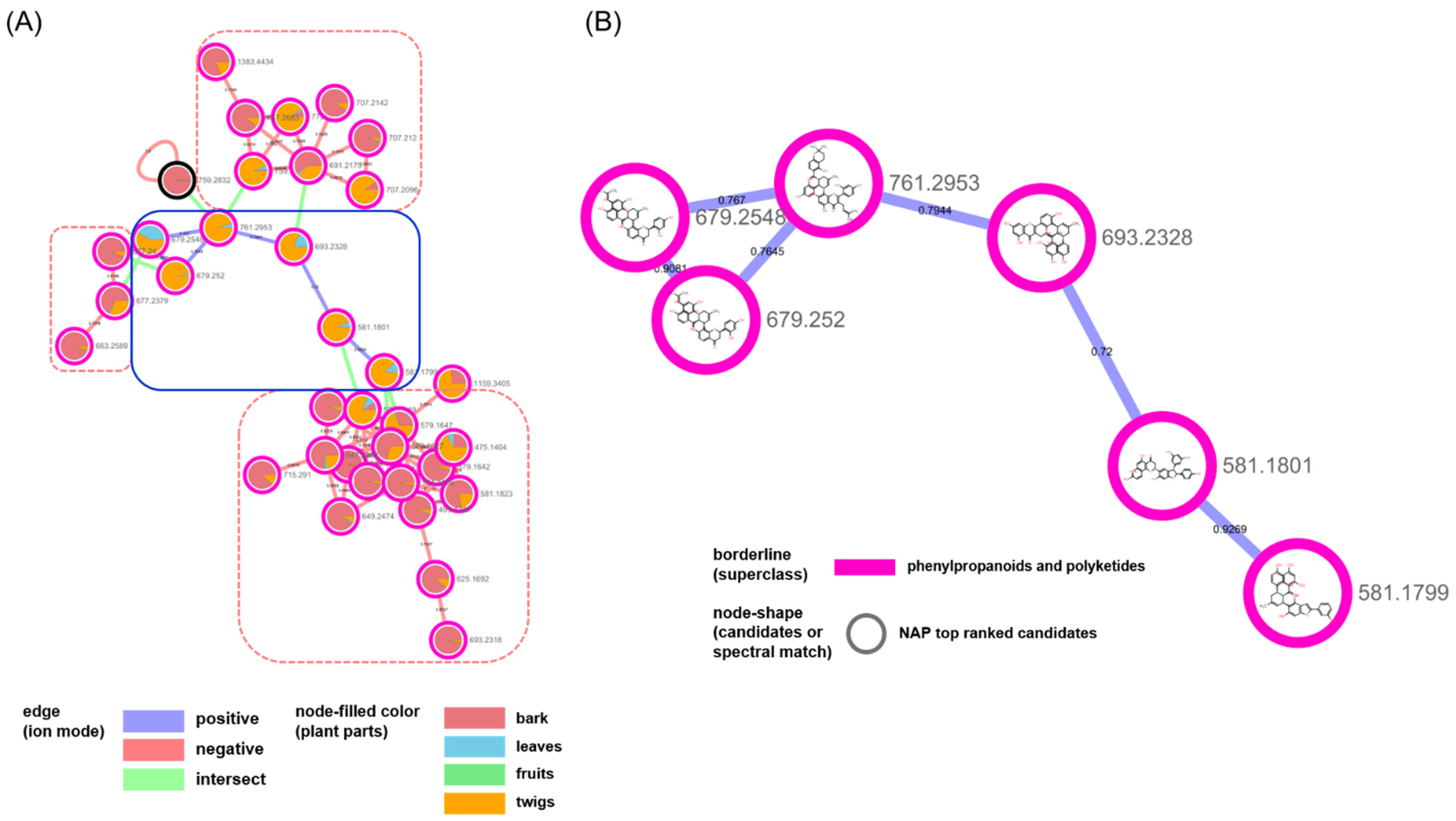
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Sample Preparation
3.3. LC-MS/MS Analysis Condition
3.4. LC-MS/MS Data Analysis
3.4.1. Molecular Networking
3.4.2. In Silico Annotation Tool, Network Annotation Propagation (NAP)
3.4.3. Chemical Class-Level Annotation Tool, MolNetEnhancer
3.4.4. Merge Polarity Networks
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef]
- Richard, N.B.; Roger, M. Wallsgrove Tansley Review No. 72. Secondary Metabolites in Plant Defence Mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar]
- Gutbrodt, B.; Gutbrodt, B.; Dorn, S.; Dorn, S.; Mody, K.; Mody, K. Drought stress affects constitutive but not induced herbivore resistance in apple plants. Arthropod-Plant. Interact. 2012, 6, 171–179. [Google Scholar] [CrossRef]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 2016, 6, 29265. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Saito, K.; Yamazaki, M. Integrated omics analysis of specialized metabolism in medicinal plants. Plant J. 2017, 90, 764–787. [Google Scholar] [CrossRef] [PubMed]
- Wangchuk, P. Therapeutic Applications of Natural Products in Herbal Medicines, Biodiscovery Programs, and Biomedicine. J. Biol. Act. Prod. Nat. 2018, 8, 1–20. [Google Scholar] [CrossRef]
- Gosetti, F.; Chiuminatto, U.; Martinotti, S.; Bolfi, B.; Ranzato, E.; Manfredi, M.; Marengo, E. Characterization of the Volatile and Nonvolatile Fractions of Heartwood Aqueous Extract from Pterocarpus marsupium and Evaluation of Its Cytotoxicity against Cancer Cell Lines. Planta Med. 2016, 82, 1295–1301. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.; Sanchez, L.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Wolfender, J.; Litaudon, M.; Touboul, D.; Queiroz, E.F. Innovative omics-based approaches for prioritisation and targeted isolation of natural products—New strategies for drug discovery. Nat. Prod. Rep. 2019, 36, 855–868. [Google Scholar] [CrossRef]
- Rockwood, A.L.; Kushnir, M.M.; Clarke, N.J. Mass Spectrometry. Princ. Appl. Clin. Mass Spectrom. 2018, 33, 33–65. [Google Scholar]
- Charles, L.; Laure, C.; Lutz, J.; Roy, R.K. Tandem mass spectrometry sequencing in the negative ion mode to read binary information encoded in sequence-defined poly(alkoxyamine amide)s. Rapid Commun. Mass Spectrom. 2016, 30, 22–28. [Google Scholar] [CrossRef]
- Arnould, M.A.; Vargas, R.; Buehner, R.W.; Wesdemiotis, C. Tandem Mass Spectrometry Characteristics of Polyester Anions and Cations Formed by Electrospray Ionization. Eur. J. Mass Spectrom. 2005, 11, 243–256. [Google Scholar] [CrossRef]
- Wang, R.; Xu, S.; Wang, N.; Xia, B.; Jiang, Y.; Wang, R. Transcriptome Analysis of Secondary Metabolism Pathway, Transcription Factors, and Transporters in Response to Methyl Jasmonate in Lycoris aurea. Front. Plant Sci. 2016, 7, 1971. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.; Bin Kang, K.; Caraballo-Rodríguez, A.M.; Nothias, L.-F.; Wandy, J.; Chen, C.; Wang, M.; Rogers, S.; Medema, M.H.; Dorrestein, P.C.; et al. MolNetEnhancer: Enhanced Molecular Networks by Integrating Metabolome Mining and Annotation Tools. Metabolites 2019, 9, 144. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Ricardo, R.; Wang, M.; Nothias, L.F.; van der Hooft, J.J.; Caraballo-Roíguez, A.M.; Fox, E.; Balunas, M.J.; Klassen, J.L.; Lopes, N.P.; et al. Propagating annotations of molecular networks using in silico fragmentation. PLoS Comput. Biol. 2018, 14, e1006089. [Google Scholar] [CrossRef]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Tang, W.; Eisenbrand, G. Chinese Drugs of Plant. Origin; Springer: Berlin/Heidelberg, Germany, 1992; pp. 669–696. [Google Scholar]
- Doi, K.; Kojima, T.; Makino, M.; Kimura, Y.; Fujimoto, Y. Studies on the Constituents of the Leaves of Morus alba L. Chem. Pharm. Bull. 2001, 49, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.; Tomar, R.; Prasad, G.; Jain, S.; Yadav, H. Complementary Hypoglycemic and Anti-Hyperglycemic Activity of Various Extracts of Fenugreek Seeds in Rats. Asian J. Biochem. 2008, 3, 182–187. [Google Scholar] [CrossRef][Green Version]
- Song, W.; Wang, H.; Bucheli, P.; Zhang, P.; Wei, D.; Lu, Y. Phytochemical Profiles of Different Mulberry (Morus sp.) Species from China. J. Agric. Food Chem. 2009, 57, 9133–9140. [Google Scholar] [CrossRef] [PubMed]
- Mena, P.; Sánchez-Salcedo, E.M.; Tassotti, M.; Martínez, J.J.; Hernández, F.; Del Rio, D. Phytochemical evaluation of eight white (Morus alba L.) and black (Morus nigra L.) mulberry clones grown in Spain based on UHPLC-ESI-MSn metabolomic profiles. Food Res. Int. 2016, 89, 1116–1122. [Google Scholar] [CrossRef]
- Kim, J.Y.; Chung, H.I.; Jung, K.; Wee, J.; Kwon, O. Chemical profiles and hypoglycemic activities of mulberry leaf extracts vary with ethanol concentration. Food Sci. Biotechnol. 2013, 22, 1–5. [Google Scholar] [CrossRef]
- Park, C.H.; Park, Y.E.; Yeo, H.J.; Yoon, J.S.; Park, S.; Kim, J.K.; Park, S.U. Comparative Analysis of Secondary Metabolites and Metabolic Profiling between Diploid and Tetraploid Morus alba L. J. Agric. Food Chem. 2021, 69, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Fukai, T.; Katayanagi, M. Kuwanon A, B, C and Oxydihydromorusin, Four New Flavones from the Root Bark of the Cultivated Mulberry Tree (Morus alba L.). Chem. Pharm. Bull. 1977, 25, 529–532. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhang, Q.; Chen, R.; Yu, D. Four new flavonoids from Morus australis. J. Asian Nat. Prod. Res. 2012, 14, 263–269. [Google Scholar] [CrossRef]
- Jung, J.; Ko, W.; Park, J.; Seo, K.; Oh, E.; Lee, D.; Lee, D.; Kim, Y.; Lim, D.; Han, D.; et al. Isoprenylated flavonoids from the root bark of Morus alba and their hepatoprotective and neuroprotective activities. Arch. Pharm. Res. 2015, 38, 2066–2075. [Google Scholar] [CrossRef]
- Huang, Q.; Lei, C.; Wang, P.; Li, J.; Li, J.; Hou, A. Isoprenylated phenolic compounds with PTP1B inhibition from Morus alba. Fitoterapia 2017, 122, 138–143. [Google Scholar] [CrossRef]
- Ha, M.T.; Seong, S.H.; Nguyen, T.D.; Cho, W.; Ah, K.J.; Ma, J.Y.; Woo, M.H.; Choi, J.S.; Min, B.S. Chalcone derivatives from the root bark of Morus alba L. act as inhibitors of PTP1B and α-glucosidase. Phytochemistry 2018, 155, 114–125. [Google Scholar] [CrossRef]
- Waters Corporation. A New Data Dependent Acquisition Alogrithm (FastDDA) for the Rapid Characterization of Complex Mixtures Waters Corporation. Appl. Note 2011, 24, 720003961. [Google Scholar]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| positive ion mode | bark (%) | leaves (%) | fruits (%) | twigs (%) |
| fatty acyls | 55.02 | 36.17 | 77.64 | 14.00 |
| flavonoids | 9.70 | 48.58 | 1.96 | 77.06 |
| isoflavonoids | 1.27 | 11.62 | 0.29 | 7.54 |
| organonitrogen compounds | 2.33 | 2.48 | 11.01 | 0.95 |
| peptidomimetics | 0.88 | 0.00 | 2.44 | 0.24 |
| prenol lipids | 5.69 | 0.72 | 3.85 | 0.20 |
| tetrapyrroles and derivatives | 25.11 | 0.43 | 2.82 | 0.00 |
| sum (%) | 100.00 | 100.00 | 100.00 | 100.00 |
| negative ion mode | bark (%) | leaves (%) | fruits (%) | twigs (%) |
| 2-arylbenzofuran flavonoids | 9.43 | 0.82 | 1.13 | 5.26 |
| anthracenes | 1.27 | 0.01 | 0.02 | 4.55 |
| benzopyrans | 48.71 | 1.36 | 0.06 | 4.20 |
| carboxylic acids and derivatives | 0.26 | 0.02 | 0.01 | 0.18 |
| coumarins and derivatives | 1.20 | 1.80 | 5.58 | 3.81 |
| diarylheptanoids | 8.91 | 2.08 | 0.12 | 13.78 |
| fatty acyls | 1.99 | 15.48 | 23.77 | 5.40 |
| flavonoids | 19.01 | 44.46 | 29.37 | 42.78 |
| furanoid lignans | 6.04 | 0.38 | 0.04 | 6.55 |
| isoflavonoids | 2.83 | 3.47 | 0.24 | 8.22 |
| organooxygen compounds | 0.29 | 25.69 | 30.76 | 4.21 |
| prenol lipids | 0.04 | 0.64 | 0.41 | 1.02 |
| steroids and steroid derivatives | 0.00 | 3.81 | 8.50 | 0.05 |
| sum (%) | 100.00 | 100.00 | 100.00 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.Y.; Park, J.; Kim, J.; Lee, J.; Yang, H. Investigation of Chemical Profiles of Different Parts of Morus alba Using a Combination of Molecular Networking Methods with Mass Spectral Data from Two Ionization Modes of LC/MS. Plants 2021, 10, 1711. https://doi.org/10.3390/plants10081711
Choi SY, Park J, Kim J, Lee J, Yang H. Investigation of Chemical Profiles of Different Parts of Morus alba Using a Combination of Molecular Networking Methods with Mass Spectral Data from Two Ionization Modes of LC/MS. Plants. 2021; 10(8):1711. https://doi.org/10.3390/plants10081711
Chicago/Turabian StyleChoi, Seong Yeon, Jinyoung Park, Juyeol Kim, Jiho Lee, and Heejung Yang. 2021. "Investigation of Chemical Profiles of Different Parts of Morus alba Using a Combination of Molecular Networking Methods with Mass Spectral Data from Two Ionization Modes of LC/MS" Plants 10, no. 8: 1711. https://doi.org/10.3390/plants10081711
APA StyleChoi, S. Y., Park, J., Kim, J., Lee, J., & Yang, H. (2021). Investigation of Chemical Profiles of Different Parts of Morus alba Using a Combination of Molecular Networking Methods with Mass Spectral Data from Two Ionization Modes of LC/MS. Plants, 10(8), 1711. https://doi.org/10.3390/plants10081711