
Transcription Factor Pso9TF Assists Xinjiang Wild Myrobalan Plum (Prunus sogdiana) PsoRPM3 Disease Resistance Protein to Resist Meloidogyne incognita

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
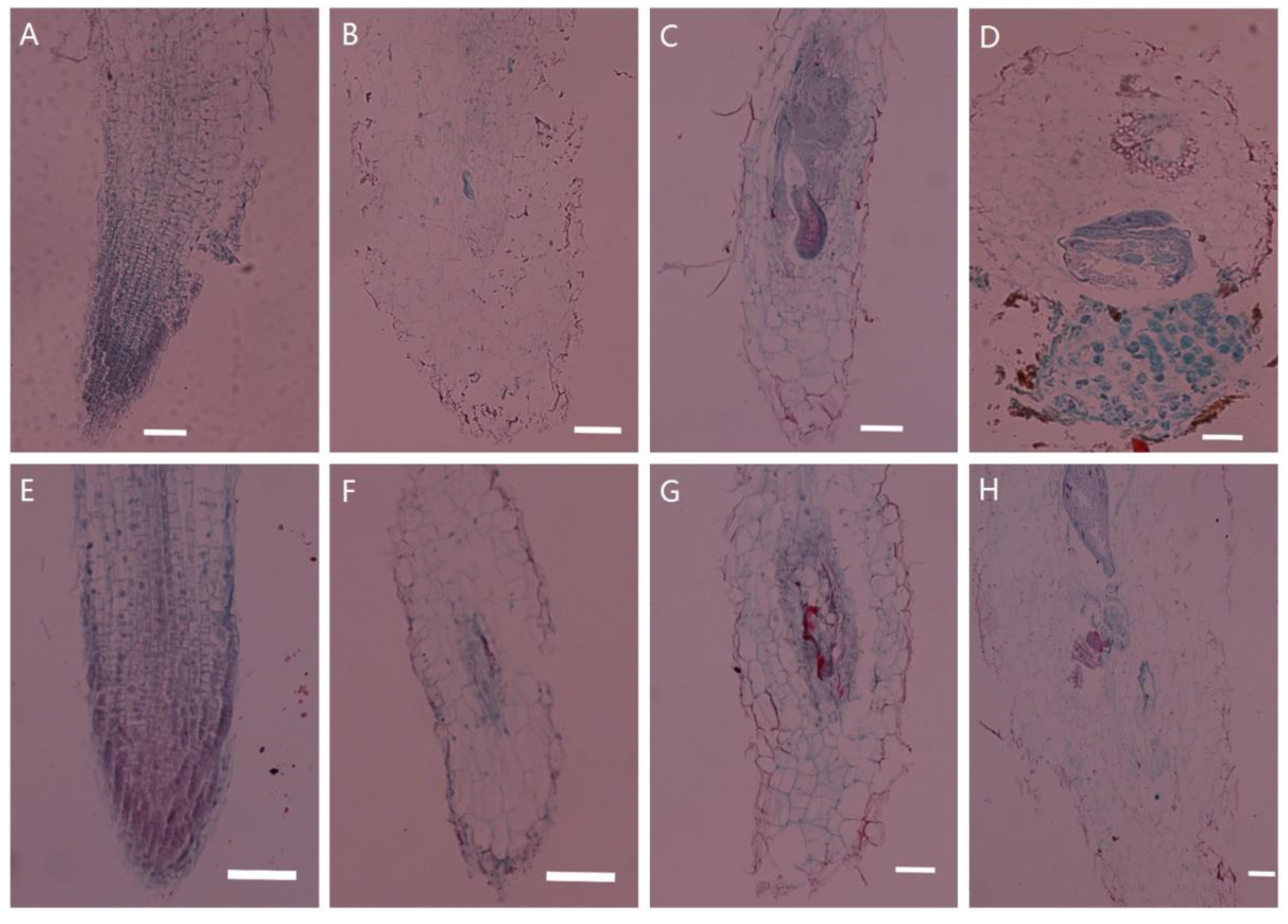
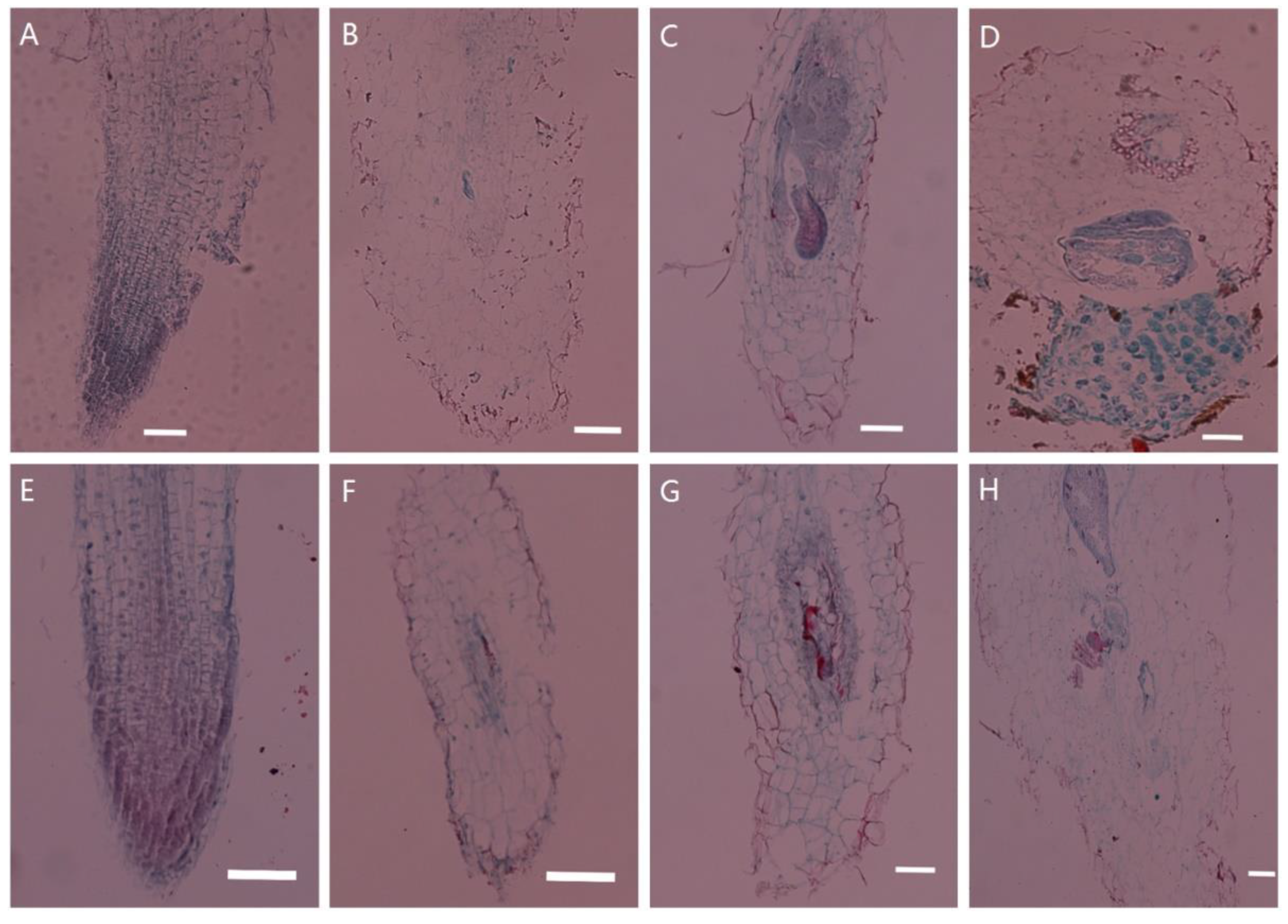
2.1. Histological Observations of Tobacco Transgenic for PsoRPM3
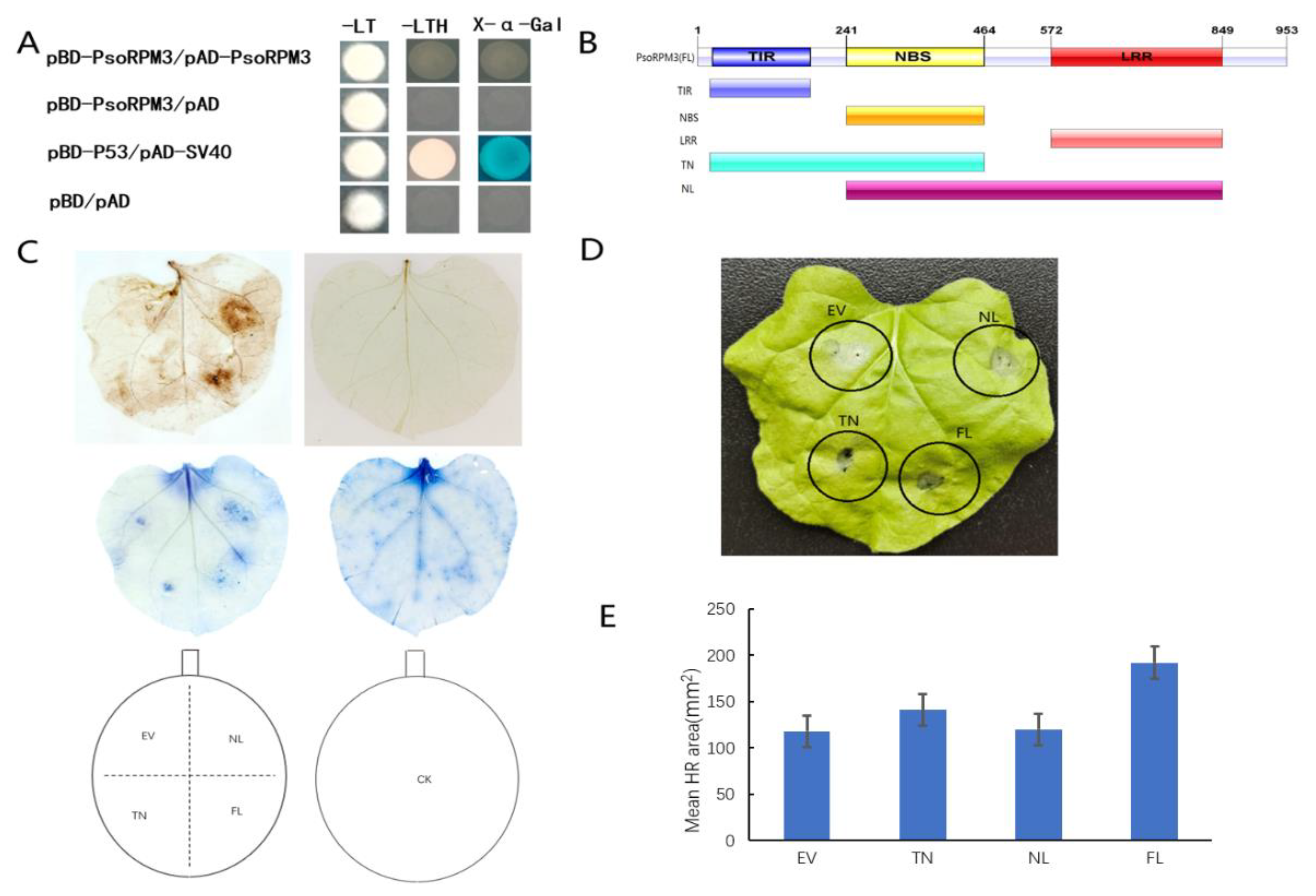
2.2. Disease Resistance Analysis of the PsoRPM3 Protein
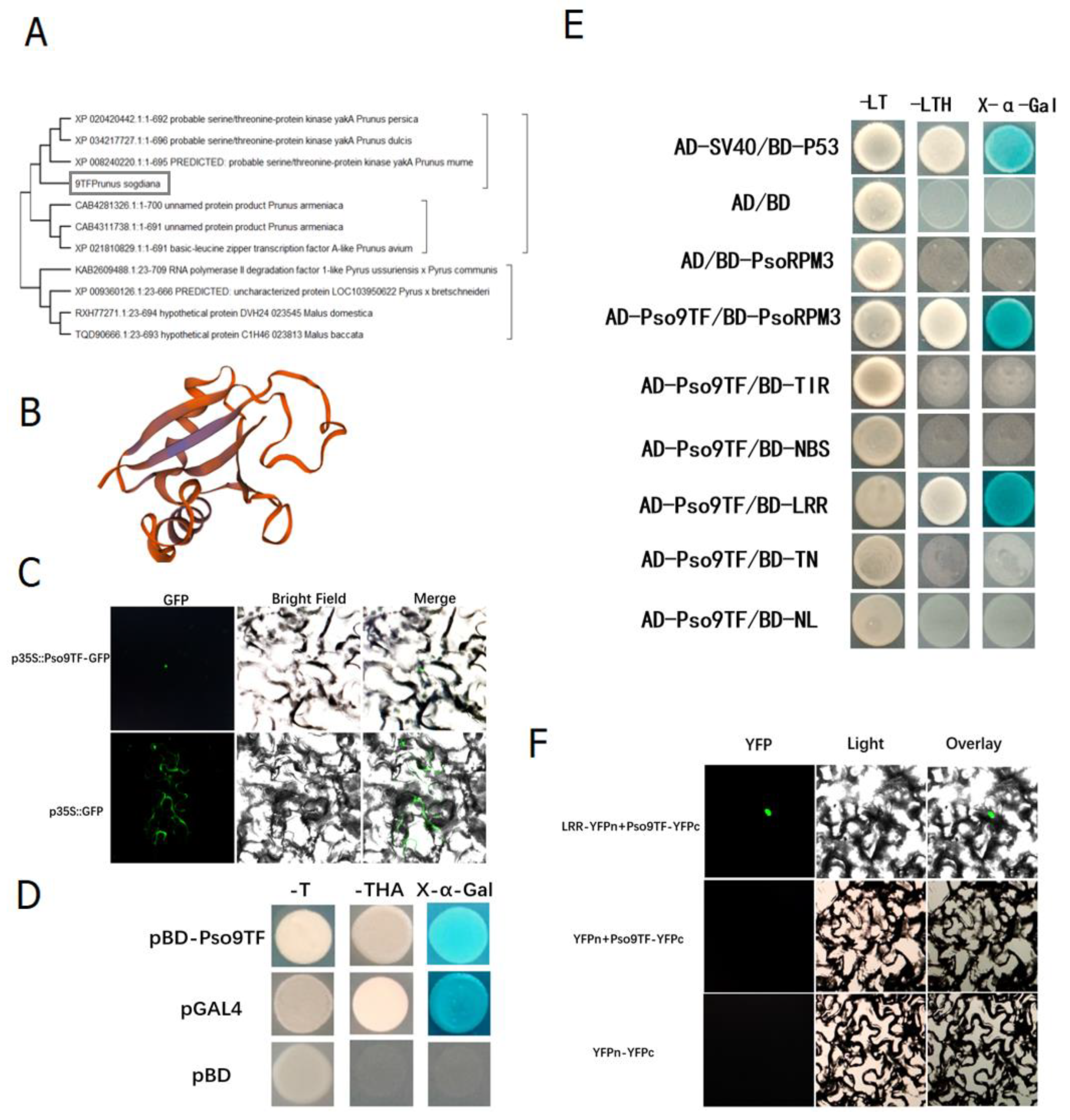
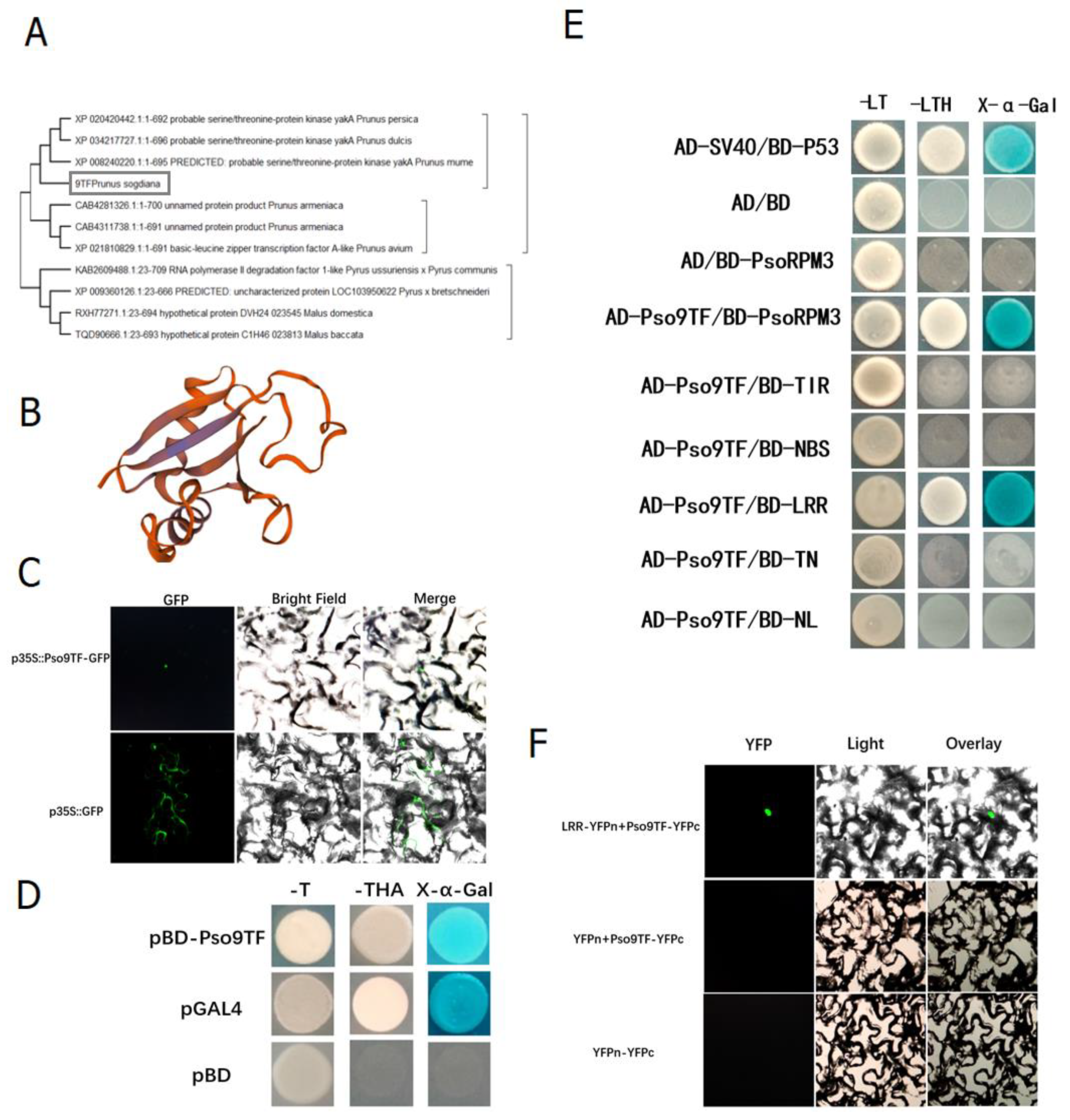
2.3. Interaction of PsoRPM3 Protein with Downstream Transcription Factor Pso9TF
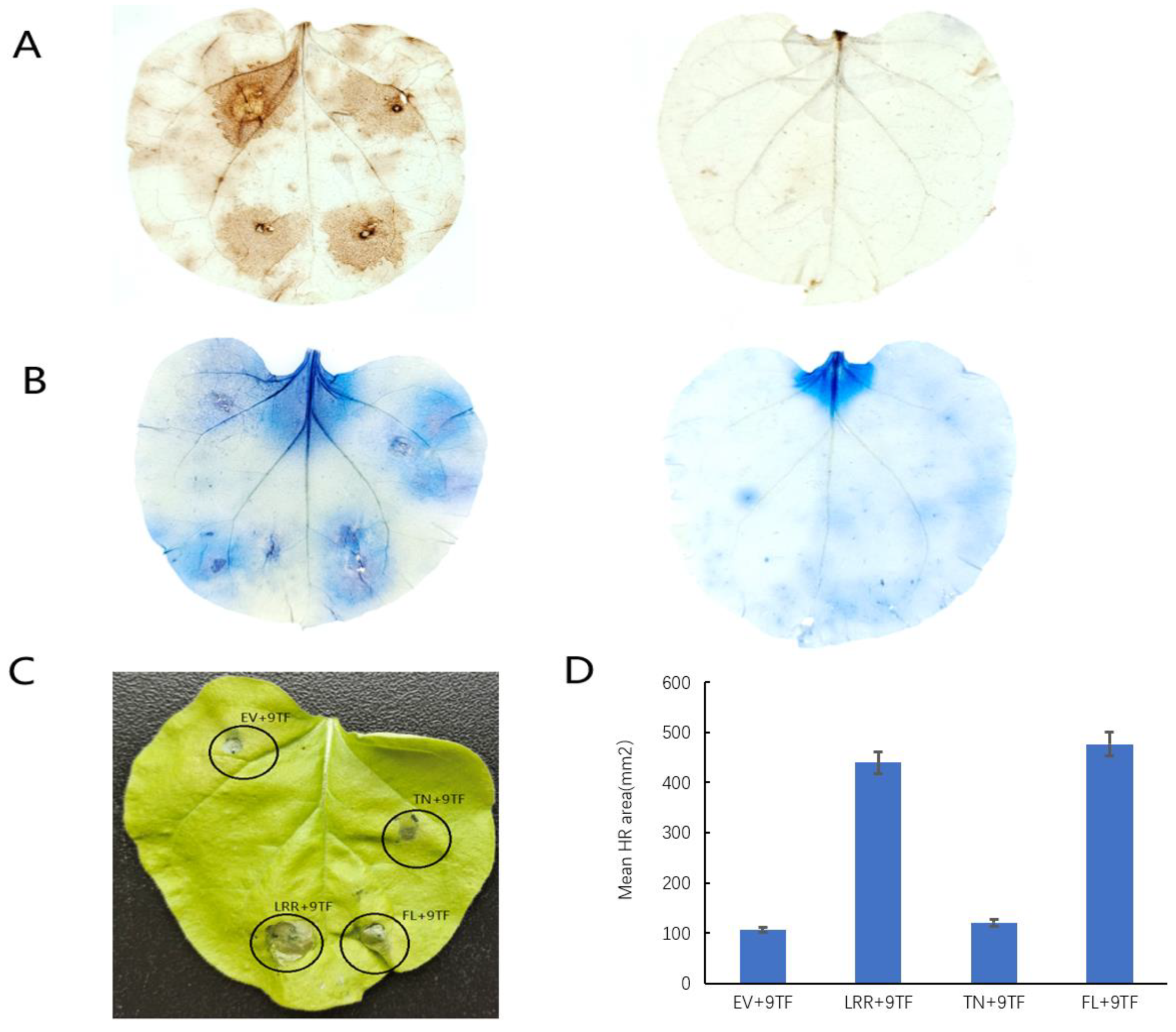
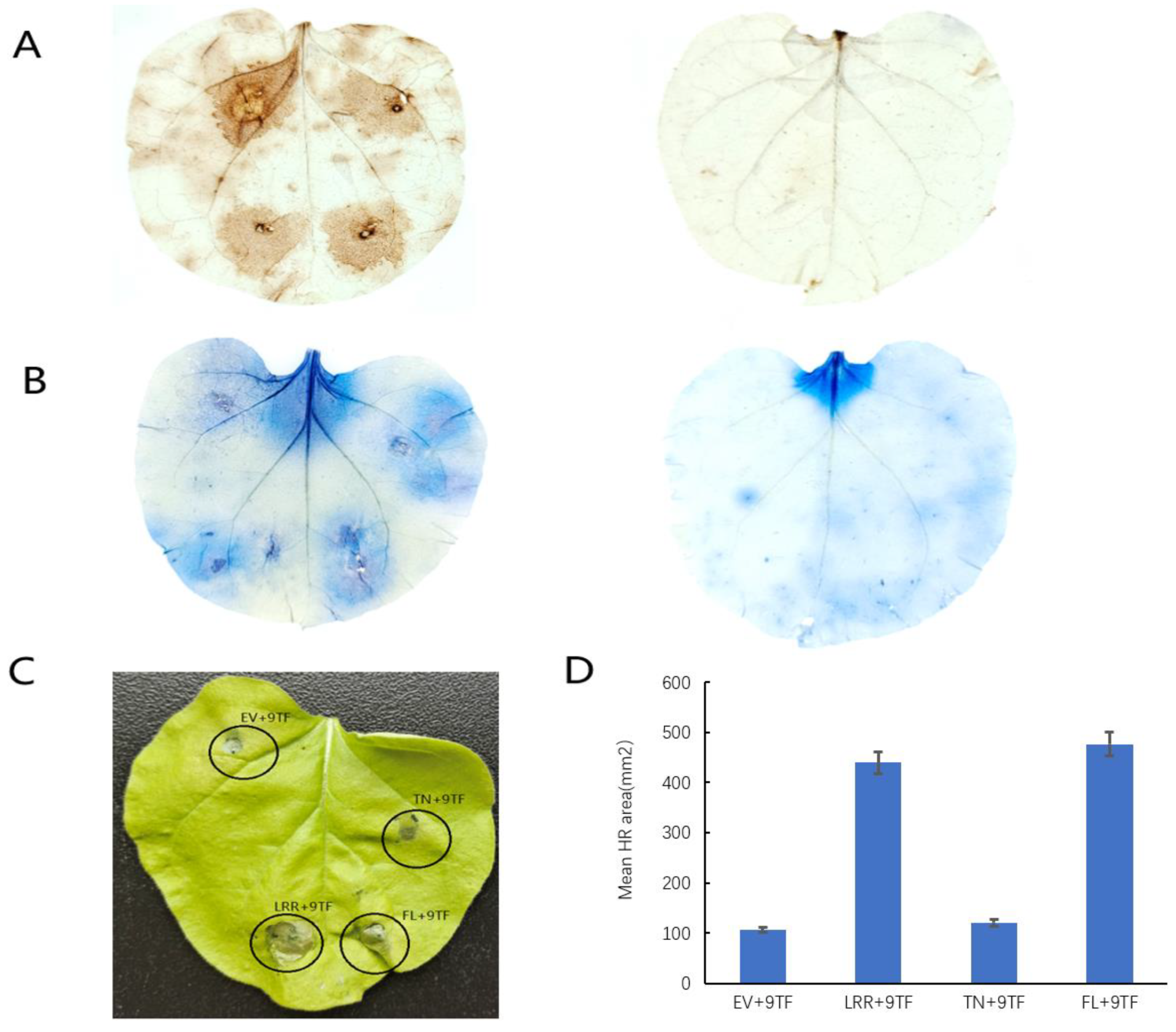
2.4. Pso9TF and PsoRPM3 Proteins together Cause Disease Resistance Responses
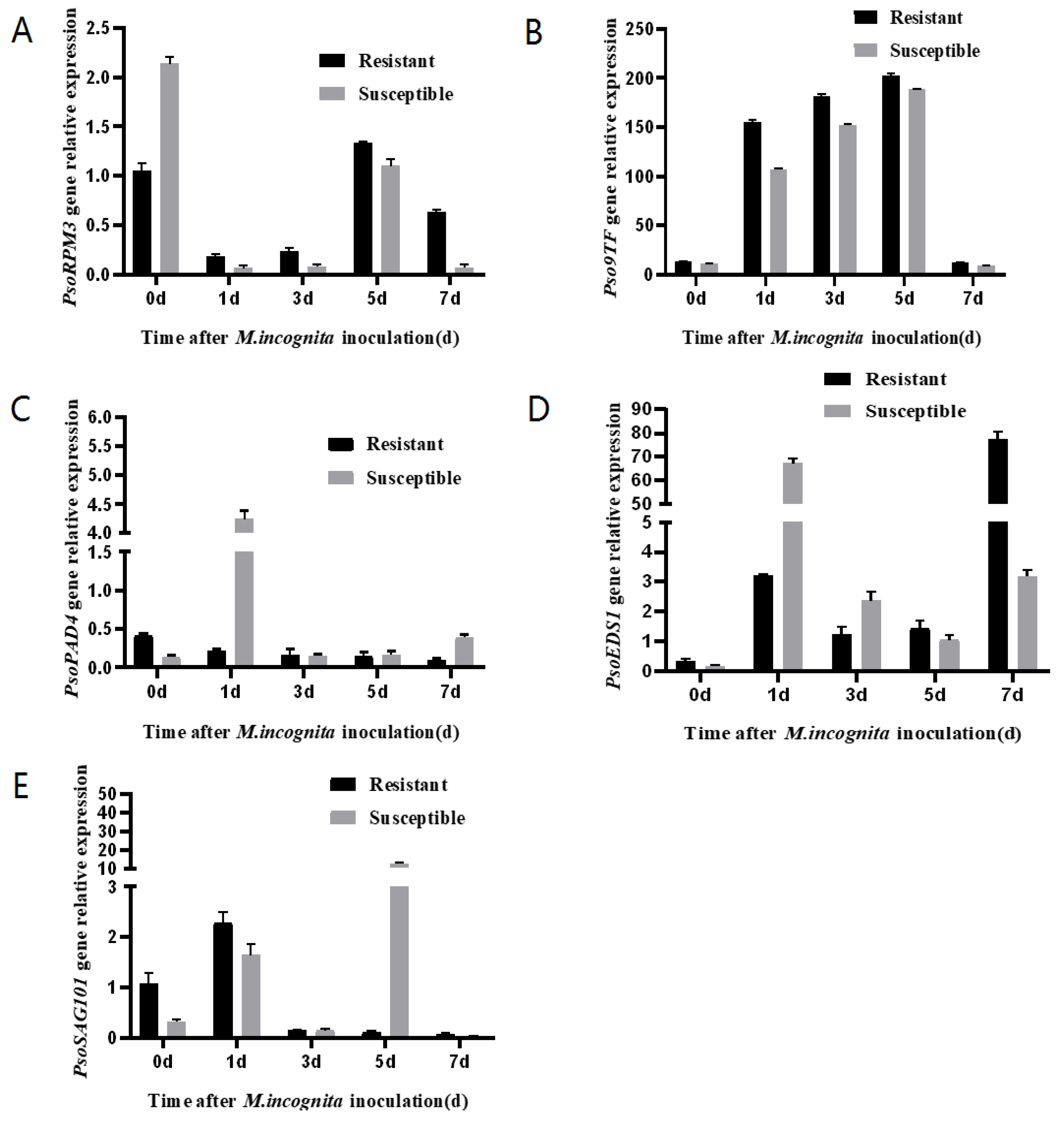
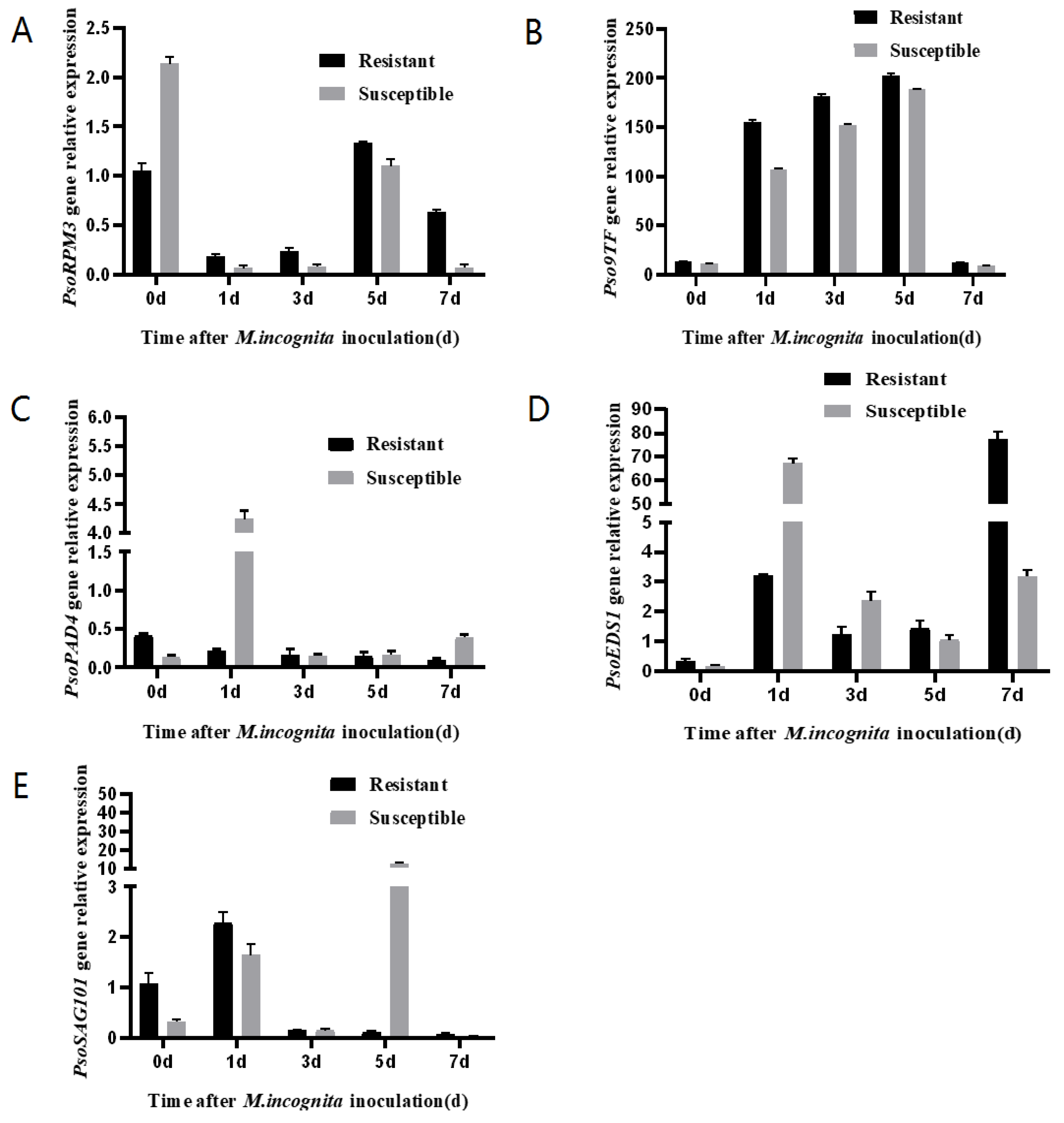
2.5. Expression of Pso9TF and Related Disease Resistance Genes in Xinjiang Wild Myrobalan Plum
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Inoculation with M. incognita
4.2. Gene Cloning and RT-qPCR Assays
4.3. Paraffin Sectioning and Histological Observation
4.4. Protein Location Analysis of Pso9TF
4.5. The Hypersensitive Response in Tobacco Leaves
4.6. Yeast Two-Hybrid Analysis
4.7. Bimolecular Fluorescence Complementation (BiFC)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Abd-Elgawad, M.M.; Askary, T.H. Impact of phytonematodes on agriculture economy. In Biocontrol agents of phytonematodes; CAB International: Wallingford, UK, 2015; pp. 3–49. [Google Scholar] [CrossRef]
- McCarter, J. Nematology: Terra incognita no more. Nat. Biotechnol. 2008, 26, 882–884. [Google Scholar] [CrossRef]
- Patel, B.; Patel, H.J.T.R. Effect of physical, cultural and chemical methods of management on population dynamics of phytonematodes in bidi tobacco nursery. Tob. Res. 1999, 25, 51–60. [Google Scholar]
- Brennan, R.J.B.; Glaze-Corcoran, S.; Wick, R.; Hashemi, M. Biofumigation: An alternative strategy for the control of plant parasitic nematodes. J. Integr. Agric. 2020, 19, 1680–1690. [Google Scholar] [CrossRef]
- Stirling, G.R.; Wong, E.; Bhuiyan, S. Pasteuria, a bacterial parasite of plant-parasitic nematodes: Its occurrence in Australian sugarcane soils and its role as a biological control agent in naturally-infested soil. Australas. Plant. Pathol. 2017, 46, 563–569. [Google Scholar] [CrossRef]
- Hussain, M.; Zouhar, M.; Ryšánek, P. Suppression of Meloidogyne incognita by the Entomopathogenic Fungus Lecanicillium muscarium. Plant. Dis. 2018, 102, 977–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimnoi, P.; Pongsilp, N.; Ruanpanun, P. Monitoring the efficiency of Streptomyces galilaeus strain KPS-C004 against root knot disease and the promotion of plant growth in the plant-parasitic nematode infested soils. Biol. Control. 2017, 114, 158–166. [Google Scholar] [CrossRef]
- Li, J.; Zou, C.; Xu, J.; Jianping, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, K.-Q. Molecular Mechanisms of Nematode-Nematophagous Microbe Interactions: Basis for Biological Control of Plant-Parasitic Nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef]
- Dong, L.Q.; Zhang, K.Q. Microbial control of plant-parasitic nematodes: A five-party interaction. Plant. Soil 2006, 288, 31–45. [Google Scholar] [CrossRef]
- Vieira, C.C.; Chen, P.; Usovsky, M.; Vuong, T.; Howland, A.D.; Nguyen, H.T.; Li, Z.; Mitchum, M.G.; Shannon, G. A major quantitative trait locus resistant to southern root-knot nematode sustains soybean yield under nematode pressure. Crop. Sci. 2021, 61, 1773–1782. [Google Scholar] [CrossRef]
- Zhang, X.; Dodds, P.; Bernoux, M. What Do We Know About NOD-Like Receptors in Plant Immunity? Annu. Rev. Phytopathol. 2017, 55, 205–229. [Google Scholar] [CrossRef]
- Cesari, S.; Moore, J.; Chen, C.; Webb, D.; Periyannan, S.; Mago, R.; Bernoux, M.; Lagudah, E.; Dodds, P.N. Cytosolic activation of cell death and stem rust resistance by cereal MLA-family CC–NLR proteins. Proc. Natl. Acad. Sci. USA 2016, 113, 10204–10209. [Google Scholar] [CrossRef] [Green Version]
- Maekawa, T.; Cheng, W.; Spiridon, L.; Töller, A.; Lukasik, E.; Saijo, Y.; Liu, P.; Shen, Q.-H.; Micluta, M.A.; Somssich, I.; et al. Coiled-Coil Domain-Dependent Homodimerization of Intracellular Barley Immune Receptors Defines a Minimal Functional Module for Triggering Cell Death. Cell Host Microbe 2011, 9, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Bernoux, M.; Bentham, A.; Newman, T.; Ve, T.; Casey, L.; Raaymakers, T.M.; Hu, J.; Croll, T.; Schreiber, K.J.; et al. Multiple functional self-association interfaces in plant TIR domains. Proc. Natl. Acad. Sci. USA 2017, 114, E2046–E2052. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.-F.; Balint-Kurti, P. Cytoplasmic and Nuclear Localizations Are Important for the Hypersensitive Response Conferred by Maize Autoactive Rp1-D21 Protein. Mol. Plant. Microbe Interact. 2015, 28, 1023–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Jeong, R.-D.; Lim, G.-H.; Yu, K.; Wang, C.; Chandra-Shekara, A.; Navarre, D.; Klessig, D.F.; Kachroo, A.; Kachroo, P. Double-Stranded RNA-Binding Protein 4 Is Required for Resistance Signaling against Viral and Bacterial Pathogens. Cell Rep. 2013, 4, 1168–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, T.; Han, M.; Qian, L.; Li, J.; Wu, M.; Han, T.; Cao, J.; Nagalakshmi, U.; Rathjen, J.P.; et al. Plant NLR immune receptor Tm-22 activation requires NB-ARC domain-mediated self-association of CC domain. PLoS Pathog. 2020, 16, e1008475. [Google Scholar] [CrossRef] [PubMed]
- El Kasmi, F.; Chung, E.-H.; Anderson, R.G.; Li, J.; Wan, L.; Eitas, T.K.; Gao, Z.; Dangl, J.L. Signaling from the plasma-membrane localized plant immune receptor RPM1 requires self-association of the full-length protein. Proc. Natl. Acad. Sci. USA 2017, 114, E7385–E7394. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.; Sornaraj, P.; de Courcy-Ireland, E.; Menz, R.I.; Kobe, B.; Ellis, J.G.; Dodds, P.; Anderson, P. An Autoactive Mutant of the M Flax Rust Resistance Protein Has a Preference for Binding ATP, Whereas Wild-Type M Protein Binds ADP. Mol. Plant. Microbe Interact. 2011, 24, 897–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tameling, W.I.; Vossen, J.H.; Albrecht, M.; Lengauer, T.; Berden, J.A.; Haring, M.A.; Cornelissen, B.J.; Takken, F.L. Mutations in the NB-ARC Domain of I-2 That Impair ATP Hydrolysis Cause Autoactivation. Plant. Physiol. 2006, 140, 1233–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedl, S.J.; Li, W.; Chao, Y.; Schwarzenbacher, R.; Shi, Y. Structure of the apoptotic protease-activating factor 1 bound to ADP. Nat. Cell Biol. 2005, 434, 926–933. [Google Scholar] [CrossRef]
- Lindner, S.; Keller, B.; Singh, S.P.; Hasenkamp, Z.; Jung, E.; Müller, M.C.; Bourras, S.; Keller, B. Single residues in the LRR domain of the wheat PM3A immune receptor can control the strength and the spectrum of the immune response. Plant. J. 2020, 104. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yan, C.; Liu, P.; Huang, Z.; Ma, R.; Zhang, C.; Wang, R.; Zhang, Y.; Martinon, F.; Miao, D.; et al. Crystal Structure of NLRC4 Reveals Its Autoinhibition Mechanism. Science 2013, 341, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Tameling, W.I.; Nooijen, C.; Ludwig, N.; Boter, M.; Slootweg, E.J.; Goverse, A.; Shirasu, K.; Joosten, M.H. RanGAP2 Mediates Nucleocytoplasmic Partitioning of the NB-LRR Immune Receptor Rx in the Solanaceae, Thereby Dictating Rx Function. Plant. Cell 2010, 22, 4176–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ade, J.; DeYoung, B.J.; Golstein, C.; Innes, R.W. Indirect activation of a plant nucleotide binding site-leucine-rich repeat protein by a bacterial protease. Proc. Natl. Acad. Sci. USA 2007, 104, 2531–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milligan, S.B.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V.M.J.T.P.C. The root knot nematode resistance gene Mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich repeat family of plant genes. Plant. Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonska, B.; Ammiraju, J.S.; Bhattarai, K.K.; Mantelin, S.; de Ilarduya, O.M.; Roberts, P.A.; Kaloshian, I. The Mi-9 Gene from Solanum arcanum Conferring Heat-Stable Resistance to Root-Knot Nematodes Is a Homolog of Mi-1. Plant. Physiol. 2006, 143, 1044–1054. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Li, H.; Zhang, L.; Zhang, J.; Xiao, J.; Ye, Z. CaMi, a root-knot nematode resistance gene from hot pepper (Capsium annuum L.) confers nematode resistance in tomato. Plant. Cell Rep. 2007, 26, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant. Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Ernst, K.; Kumar, A.; Kriseleit, D.; Kloos, D.-U.; Phillips, M.S.; Ganal, M.W. The broad-spectrum potato cyst nematode resistance gene (Hero) from tomato is the only member of a large gene family of NBS-LRR genes with an unusual amino acid repeat in the LRR region. Plant. J. 2002, 31, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Van Der Vossen, E.A.G.; Van Der Voort, J.N.A.M.R.; Kanyuka, K.; Bendahmane, A.; Sandbrink, H.; Baulcombe, D.C.; Bakker, J.; Stiekema, W.J.; Klein-Lankhorst, R.M. Homologues of a single resistance-gene cluster in potato confer resistance to distinct pathogens: A virus and a nematode. Plant. J. 2000, 23, 567–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paal, J.; Henselewski, H.; Muth, J.; Meksem, K.; Menéndez, C.M.; Salamini, F.; Ballvora, A.; Gebhardt, C. Molecular cloning of the potato Gro1-4 gene conferring resistance to pathotype Ro1 of the root cyst nematode Globodera rostochiensis, based on a candidate gene approach. Plant. J. 2004, 38, 285–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claverie, M.; Dirlewanger, E.; Bosselut, N.; Van Ghelder, C.; Voisin, R.; Kleinhentz, M.; Lafargue, B.; Abad, P.; Rosso, M.-N.; Chalhoub, B.; et al. The Ma Gene for Complete-Spectrum Resistance to Meloidogyne Species in Prunus Is a TNL with a Huge Repeated C-Terminal Post-LRR Region. Plant. Physiol. 2011, 156, 779–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ghelder, C.; Esmenjaud, D. TNL genes in peach: Insights into the post-LRR domain. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, R.; Fang, H.; Zhang, C.; Zhang, F.; Hao, Z.; You, X.; Shi, X.; Park, C.H.; Hua, K.; et al. Two VOZ transcription factors link an E3 ligase and an NLR immune receptor to modulate immunity in rice. Mol. Plant. 2021, 14, 253–266. [Google Scholar] [CrossRef]
- Zang, Z.; Lv, Y.; Liu, S.; Yang, W.; Ci, J.; Ren, X.; Wang, Z.; Wu, H.; Ma, W.; Jiang, L.; et al. A Novel ERF Transcription Factor, ZmERF105, Positively Regulates Maize Resistance to Exserohilum turcicum. Front. Plant. Sci. 2020, 11, 850. [Google Scholar] [CrossRef]
- Van Der Hoorn, R.A.; Kamoun, S. From Guard to Decoy: A New Model for Perception of Plant Pathogen Effectors. Plant. Cell 2008, 20, 2009–2017. [Google Scholar] [CrossRef] [Green Version]
- Warmerdam, S.; Sterken, M.G.; Sukarta, O.C.A.; Van Schaik, C.C.; Oortwijn, M.E.P.; Lozano-Torres, J.L.; Bakker, J.; Smant, G.; Goverse, A. The TIR-NB-LRR pair DSC1 and WRKY19 contributes to basal immunity of Arabidopsis to the root-knot nematode Meloidogyne incognita. BMC Plant. Biol. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Xiao, S.; Calis, O.; Patrick, E.; Zhang, G.; Charoenwattana, P.; Muskett, P.; Parker, J.E.; Turner, J.G. The atypical resistance gene, RPW8, recruits components of basal defence for powdery mildew resistance in Arabidopsis. Plant. J. 2005, 42, 95–110. [Google Scholar] [CrossRef]
- Chandra-Shekara, A.; Navarre, D.; Kachroo, A.; Kang, H.-G.; Klessig, D.; Kachroo, P. Signaling requirements and role of salicylic acid in HRT- and rrt-mediated resistance to turnip crinkle virus in Arabidopsis. Plant. J. 2004, 40, 647–659. [Google Scholar] [CrossRef]
- Wagner, S.; Stuttmann, J.; Rietz, S.; Guerois, R.; Brunstein, E.; Bautor, J.; Niefind, K.; Parker, J.E. Structural Basis for Signaling by Exclusive EDS1 Heteromeric Complexes with SAG101 or PAD4 in Plant Innate Immunity. Cell Host Microbe 2013, 14, 619–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Jeong, R.-D.; Venugopal, S.C.; Lapchyk, L.; Navarre, D.; Kachroo, A.; Kachroo, P. SAG101 Forms a Ternary Complex with EDS1 and PAD4 and Is Required for Resistance Signaling against Turnip Crinkle Virus. PLoS Pathog. 2011, 7, e1002318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Gobbato, E.; Kracher, B.; Qiu, J.; Bautor, J.; Parker, J.E. A core function of EDS1 with PAD4 is to protect the salicylic acid defense sector in Arabidopsis immunity. New Phytol. 2016, 213, 1802–1817. [Google Scholar] [CrossRef] [Green Version]
- García, A.V.; Blanvillain-Baufumé, S.; Huibers, R.P.; Wiermer, M.; Li, G.; Gobbato, E.; Rietz, S.; Parker, J.E. Balanced Nuclear and Cytoplasmic Activities of EDS1 Are Required for a Complete Plant Innate Immune Response. PLoS Pathog. 2010, 6, e1000970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiermer, M.; Feys, B.J.; Parker, J.E. Plant immunity: The EDS1 regulatory node. Curr. Opin. Plant. Biol. 2005, 8, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K. Functional Verification and Mechanism Analysis of PsoRPM3 from Prunus sogdiana in Reponse to Meloidogyne Incognita. Ph.D. Thesis, China Agricultural University, Beijing, China, 2020. [Google Scholar]
- Sun, Y.; Zhu, Y.-X.; Balint-Kurti, P.J.; Wang, G.-F. Fine-Tuning Immunity: Players and Regulators for Plant NLRs. Trends Plant. Sci. 2020, 25, 695–713. [Google Scholar] [CrossRef]
- Hamada, N.; Yimer, H.; Williamson, V.M.; Siddique, S. Chemical Hide and Seek: Nematode’s Journey to Its Plant Host. Mol. Plant. 2020, 13, 541–543. [Google Scholar] [CrossRef]
- Mejias, J.; Truong, N.M.; Abad, P.; Favery, B.; Quentin, M. Plant Proteins and Processes Targeted by Parasitic Nematode Effectors. Front. Plant. Sci. 2019, 10, 970. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.J.; Sohn, K.H.; Wan, L.; Bernoux, M.; Sarris, P.F.; Segonzac, C.; Ve, T.; Ma, Y.; Saucet, S.B.; Ericsson, D.J.; et al. Structural Basis for Assembly and Function of a Heterodimeric Plant Immune Receptor. Science 2014, 344, 299–303. [Google Scholar] [CrossRef]
- Wan, L.; Essuman, K.; Anderson, R.G.; Sasaki, Y.; Monteiro, F.; Chung, E.-H.; Nishimura, E.O.; DiAntonio, A.; Milbrandt, J.; Dangl, J.L.; et al. TIR domains of plant immune receptors are NAD+-cleaving enzymes that promote cell death. Science 2019, 365, 799–803. [Google Scholar] [CrossRef]
- Nimma, S.; Ve, T.; Williams, S.; Kobe, B. Towards the structure of the TIR-domain signalosome. Curr. Opin. Struct. Biol. 2017, 43, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Ng, D.W.-K.; Abeysinghe, J.K.; Kamali, M. Regulating the Regulators: The Control of Transcription Factors in Plant Defense Signaling. Int. J. Mol. Sci. 2018, 19, 3737. [Google Scholar] [CrossRef] [Green Version]
- Inoue, H.; Hayashi, N.; Matsushita, A.; Xinqiong, L.; Nakayama, A.; Sugano, S.; Jiang, C.-J.; Takatsuji, H. Blast resistance of CC-NB-LRR protein Pb1 is mediated by WRKY45 through protein–protein interaction. Proc. Natl. Acad. Sci. USA 2013, 110, 9577–9582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Wu, Y.; Wu, D.; Rao, W.; Guo, J.; Ma, Y.; Wang, Z.; Shangguan, X.; Wang, H.; Xu, C.; et al. The Coiled-Coil and Nucleotide Binding Domains of BROWN PLANTHOPPER RESISTANCE14 Function in Signaling and Resistance against Planthopper in Rice. Plant. Cell 2017, 29, 3157–3185. [Google Scholar] [CrossRef] [Green Version]
- Lapin, D.; Kovacova, V.; Sun, X.; Dongus, J.A.; Bhandari, D.; Von Born, P.; Bautor, J.; Guarneri, N.; Rzemieniewski, J.; Stuttmann, J.; et al. A Coevolved EDS1-SAG101-NRG1 Module Mediates Cell Death Signaling by TIR-Domain Immune Receptors. Plant. Cell 2019, 31, 2430–2455. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, K.; Sato, M.; Stoddard, T.; Glazebrook, J.; Katagiri, F. Network Properties of Robust Immunity in Plants. PLoS Genet. 2009, 5, e1000772. [Google Scholar] [CrossRef] [Green Version]
- Bartsch, M.; Gobbato, E.; Bednarek, P.; Debey, S.; Schultze, J.; Bautor, J.; Parker, J.E. Salicylic Acid–Independent ENHANCED DISEASE SUSCEPTIBILITY1 Signaling in Arabidopsis Immunity and Cell Death Is Regulated by the Monooxygenase FMO1 and the Nudix Hydrolase NUDT7. Plant. Cell 2006, 18, 1038–1051. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, S.C.; Jeong, R.-D.; Mandal, M.K.; Zhu, S.; Chandra-Shekara, A.C.; Xia, Y.; Hersh, M.; Stromberg, A.J.; Navarre, D.; Kachroo, A.; et al. Enhanced Disease Susceptibility 1 and Salicylic Acid Act Redundantly to Regulate Resistance Gene-Mediated Signaling. PLoS Genet. 2009, 5, e1000545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Qiu, J.; Zhou, Y.; Bhandari, D.D.; Zhao, C.; Bautor, J.; Parker, J.E. Antagonism of Transcription Factor MYC2 by EDS1/PAD4 Complexes Bolsters Salicylic Acid Defense in Arabidopsis Effector-Triggered Immunity. Mol. Plant. 2018, 11, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Rietz, S.; Stamm, A.; Malonek, S.; Wagner, S.; Becker, D.; Medina-Escobar, N.; Vlot, A.C.; Feys, B.J.; Niefind, K.; Parker, J.E. Different roles of Enhanced Disease Susceptibility1 (EDS1) bound to and dissociated from Phytoalexin Deficient4 (PAD4) in Arabidopsis immunity. New Phytol. 2011, 191, 107–119. [Google Scholar] [CrossRef]
- Gantner, J.; Ordon, J.; Kretschmer, C.; Guerois, R.; Stuttmann, J. An EDS1-SAG101 Complex Is Essential for TNL-Mediated Immunity in Nicotiana benthamiana. Plant. Cell 2019, 31, 2456–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, J.; Priya, P.; Dastidar, S.G.; Das, S. Physical interaction between nuclear accumulated CC-NB-ARC-LRR protein and WRKY64 promotes EDS1 dependent Fusarium wilt resistance in chickpea. Plant. Sci. 2018, 276, 111–133. [Google Scholar] [CrossRef]
- Youssef, R.M.; Macdonald, M.H.; Brewer, E.P.; Bauchan, G.R.; Kim, K.-H.; Matthews, B.F. Ectopic expression of AtPAD4 broadens resistance of soybean to soybean cyst and root-knot nematodes. BMC Plant. Biol. 2013, 13, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priya, D.B.; Somasekhar, N.; Prasad, J.; Kirti, P. Transgenic tobacco plants constitutively expressing Arabidopsis NPR1 show enhanced resistance to root-knot nematode, Meloidogyne incognita. BMC Res. Notes 2011, 4, 231. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Chen, W.; Chen, X.; Zhu, X.; Guan, P.; Hu, J. CCS52 and DEL1 function in root-knot nematode giant cell development in Xinjiang wild myrobalan plum (Prunus sogdiana Vassilcz). Protoplasma 2020, 257, 1333–1344. [Google Scholar] [CrossRef]
- Tran, B.Q.; Jung, S. Modulation of chloroplast components and defense responses during programmed cell death in tobacco infected with Pseudomonas syringae. Biochem. Biophys. Res. Commun. 2020, 528, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chang, C.; Gu, C.; Tang, S.Y.; Xie, Q.; Shen, Q.-H. An E3 Ligase Affects the NLR Receptor Stability and Immunity to Powdery Mildew. Plant. Physiol. 2016, 172, 2504–2515. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Xiao, K.; Pu, W.; Liu, Z.; Wang, Y.; Gao, C.; Luo, S.; Xu, Y.; Guan, P.; Hu, J. Transcription Factor Pso9TF Assists Xinjiang Wild Myrobalan Plum (Prunus sogdiana) PsoRPM3 Disease Resistance Protein to Resist Meloidogyne incognita. Plants 2021, 10, 1561. https://doi.org/10.3390/plants10081561
Zhu H, Xiao K, Pu W, Liu Z, Wang Y, Gao C, Luo S, Xu Y, Guan P, Hu J. Transcription Factor Pso9TF Assists Xinjiang Wild Myrobalan Plum (Prunus sogdiana) PsoRPM3 Disease Resistance Protein to Resist Meloidogyne incognita. Plants. 2021; 10(8):1561. https://doi.org/10.3390/plants10081561
Chicago/Turabian StyleZhu, Haifeng, Kun Xiao, Wenjiang Pu, Zhenhua Liu, Yan Wang, Chaoyuan Gao, Sifang Luo, Yue Xu, Pingyin Guan, and Jianfang Hu. 2021. "Transcription Factor Pso9TF Assists Xinjiang Wild Myrobalan Plum (Prunus sogdiana) PsoRPM3 Disease Resistance Protein to Resist Meloidogyne incognita" Plants 10, no. 8: 1561. https://doi.org/10.3390/plants10081561
APA StyleZhu, H., Xiao, K., Pu, W., Liu, Z., Wang, Y., Gao, C., Luo, S., Xu, Y., Guan, P., & Hu, J. (2021). Transcription Factor Pso9TF Assists Xinjiang Wild Myrobalan Plum (Prunus sogdiana) PsoRPM3 Disease Resistance Protein to Resist Meloidogyne incognita. Plants, 10(8), 1561. https://doi.org/10.3390/plants10081561