
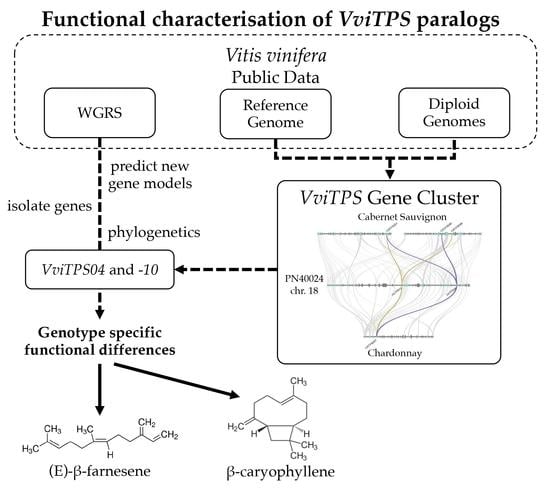
Seeing the Forest through the (Phylogenetic) Trees: Functional Characterisation of Grapevine Terpene Synthase (VviTPS) Paralogues and Orthologues


Abstract
:
1. Introduction
2. Results
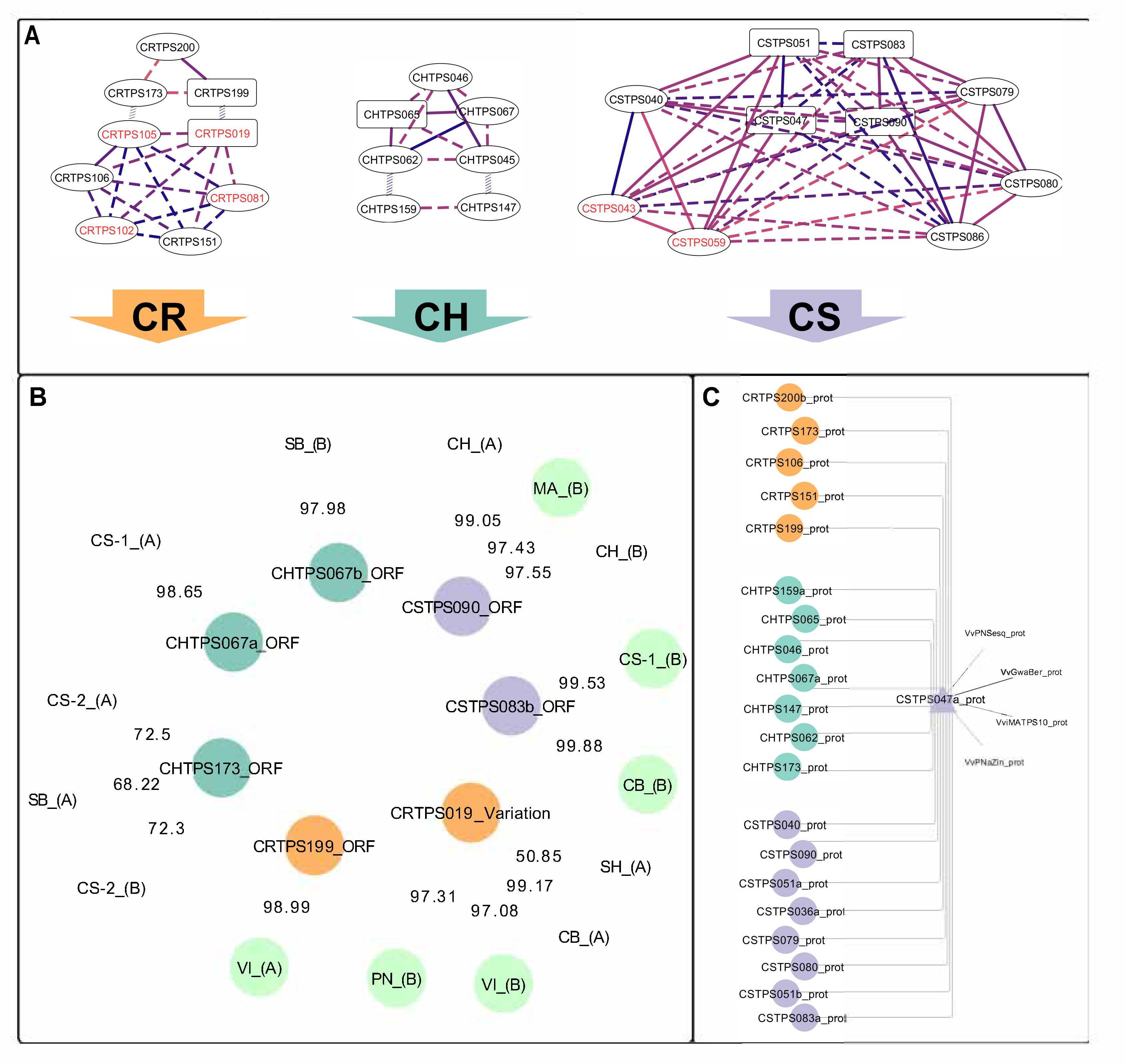
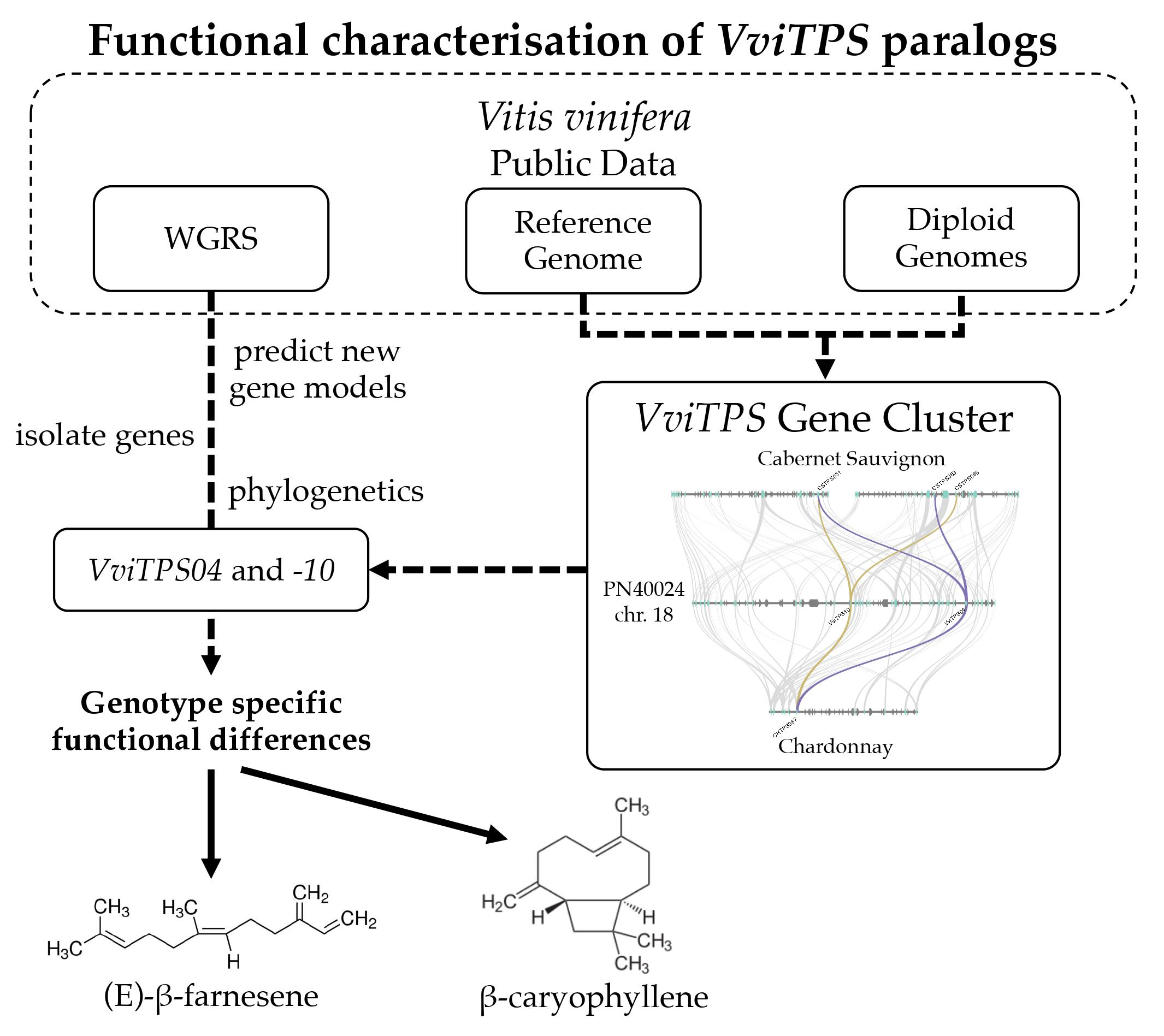
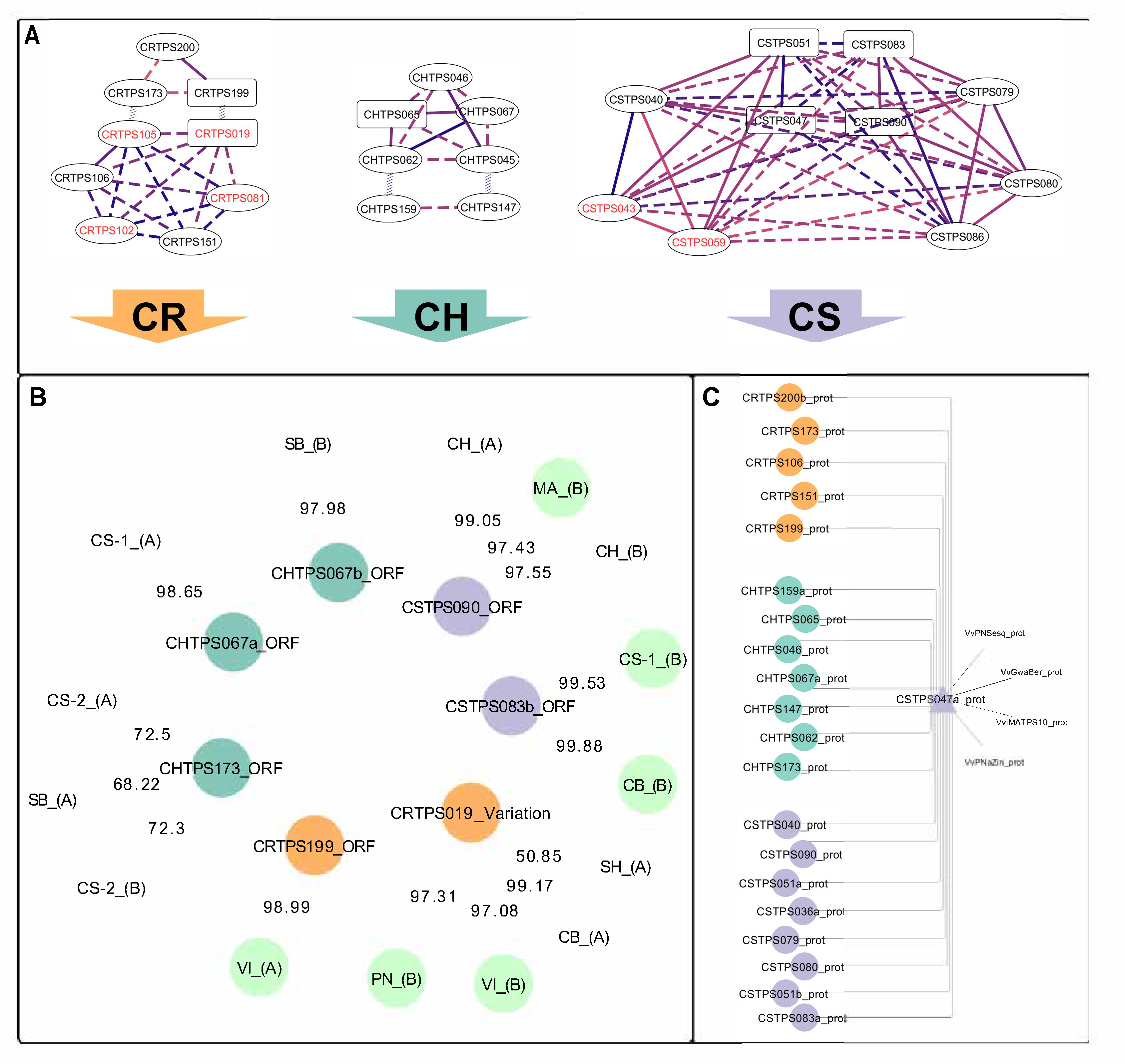
2.1. Isolated VviTPS04 and -10-Like Paralogues
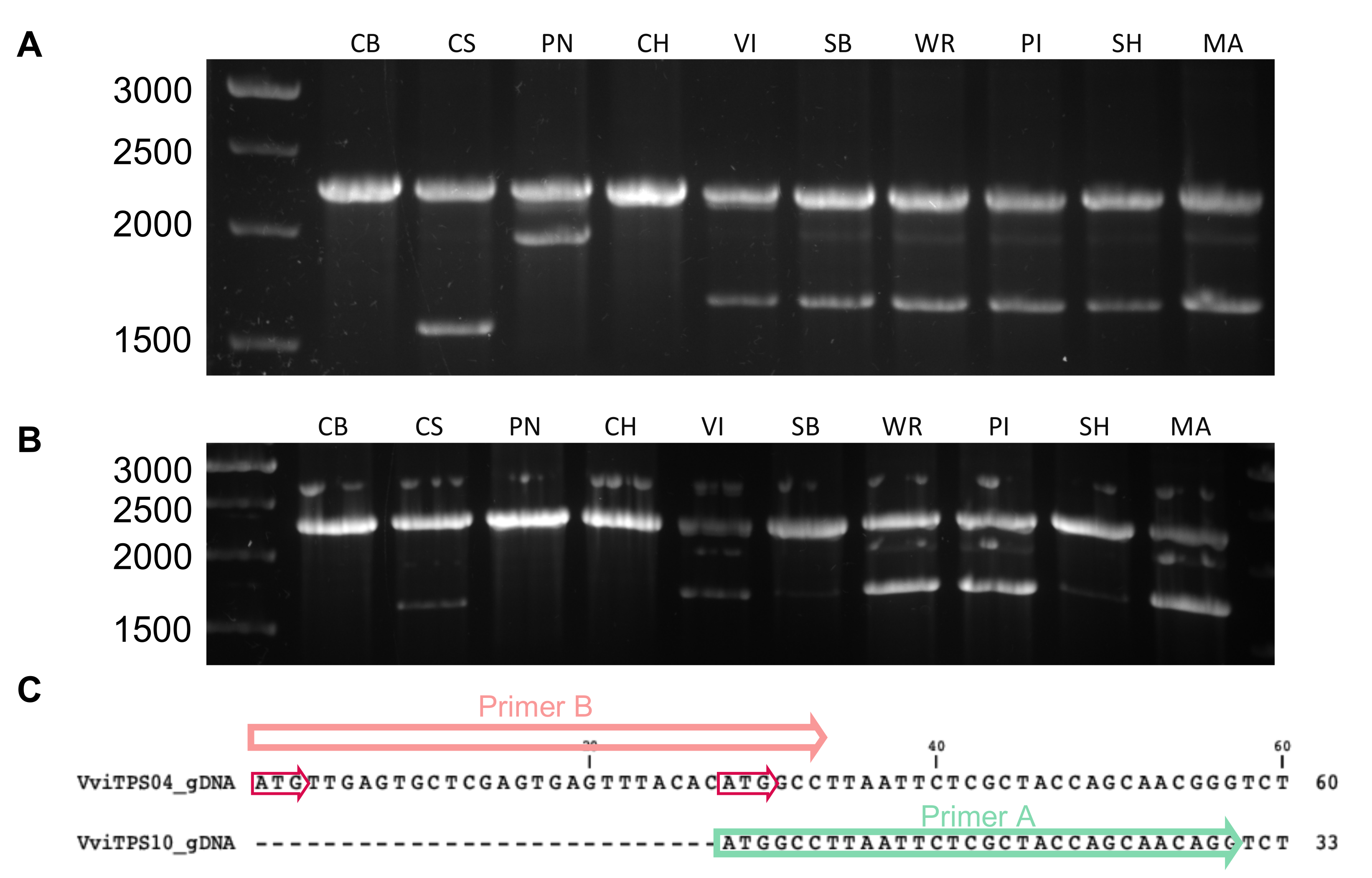
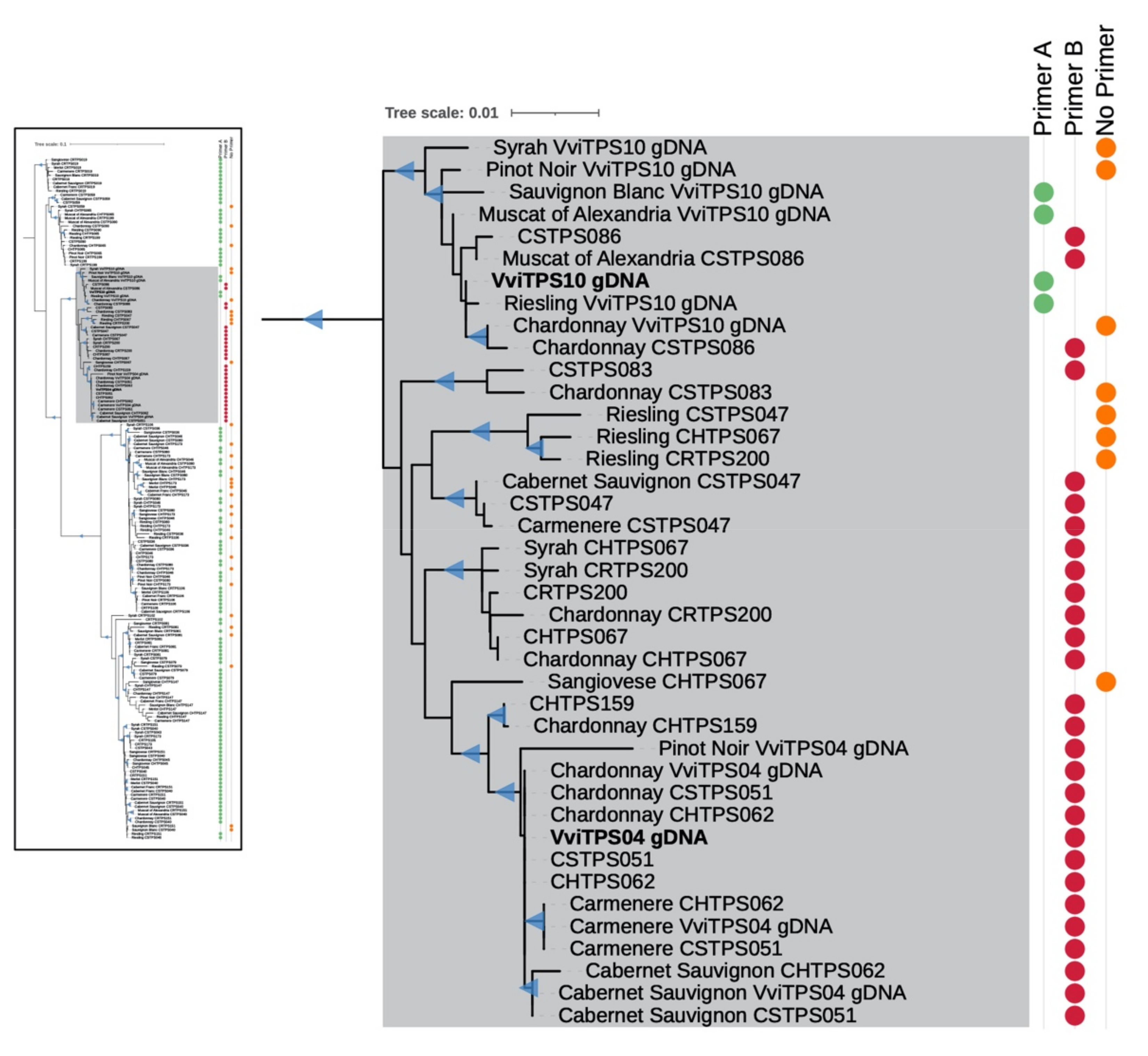
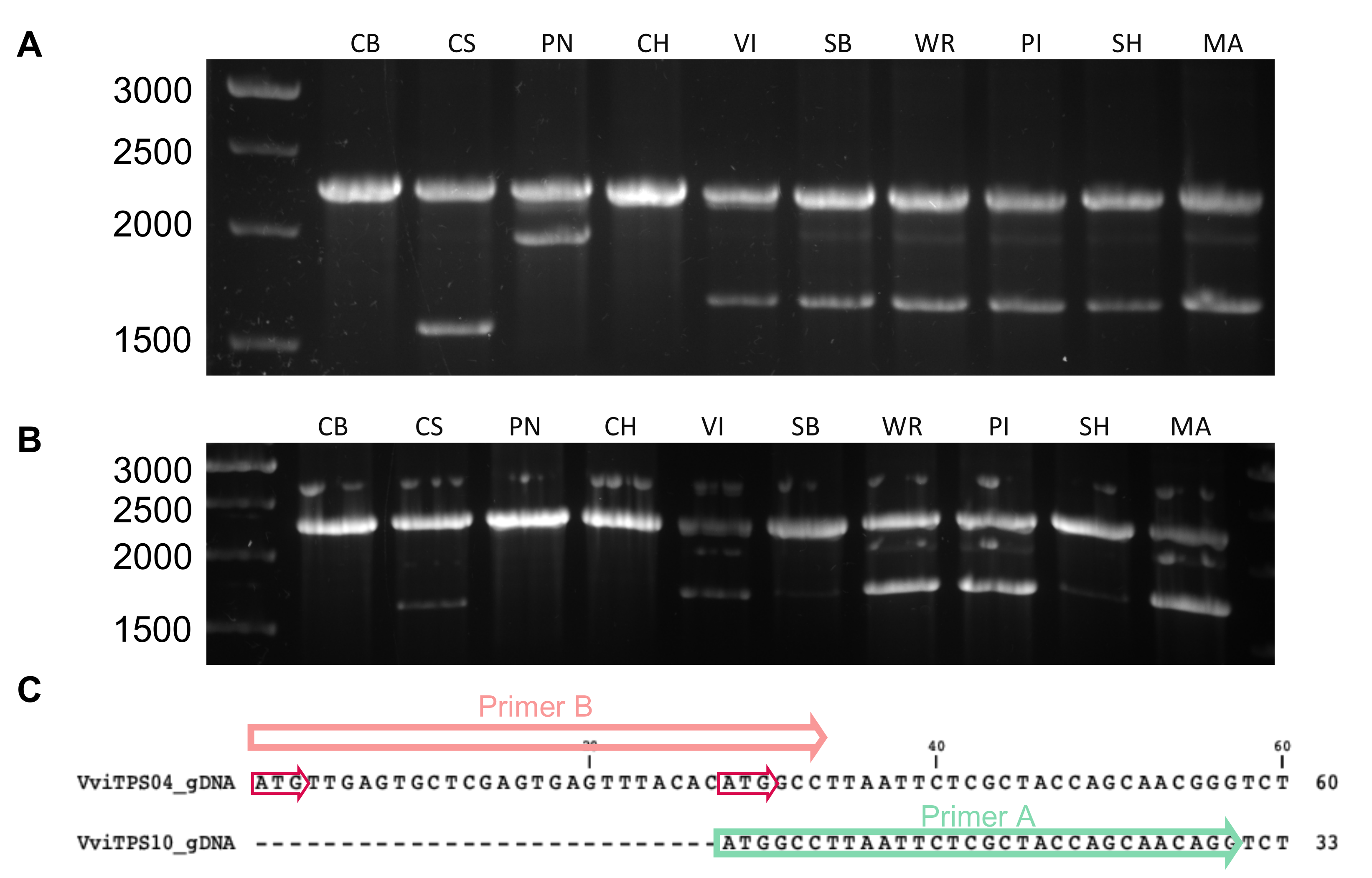
2.2. Isolation and Sequence Comparison of VviTPS04 and -10 Homologs from Nine Grapevine Genotypes
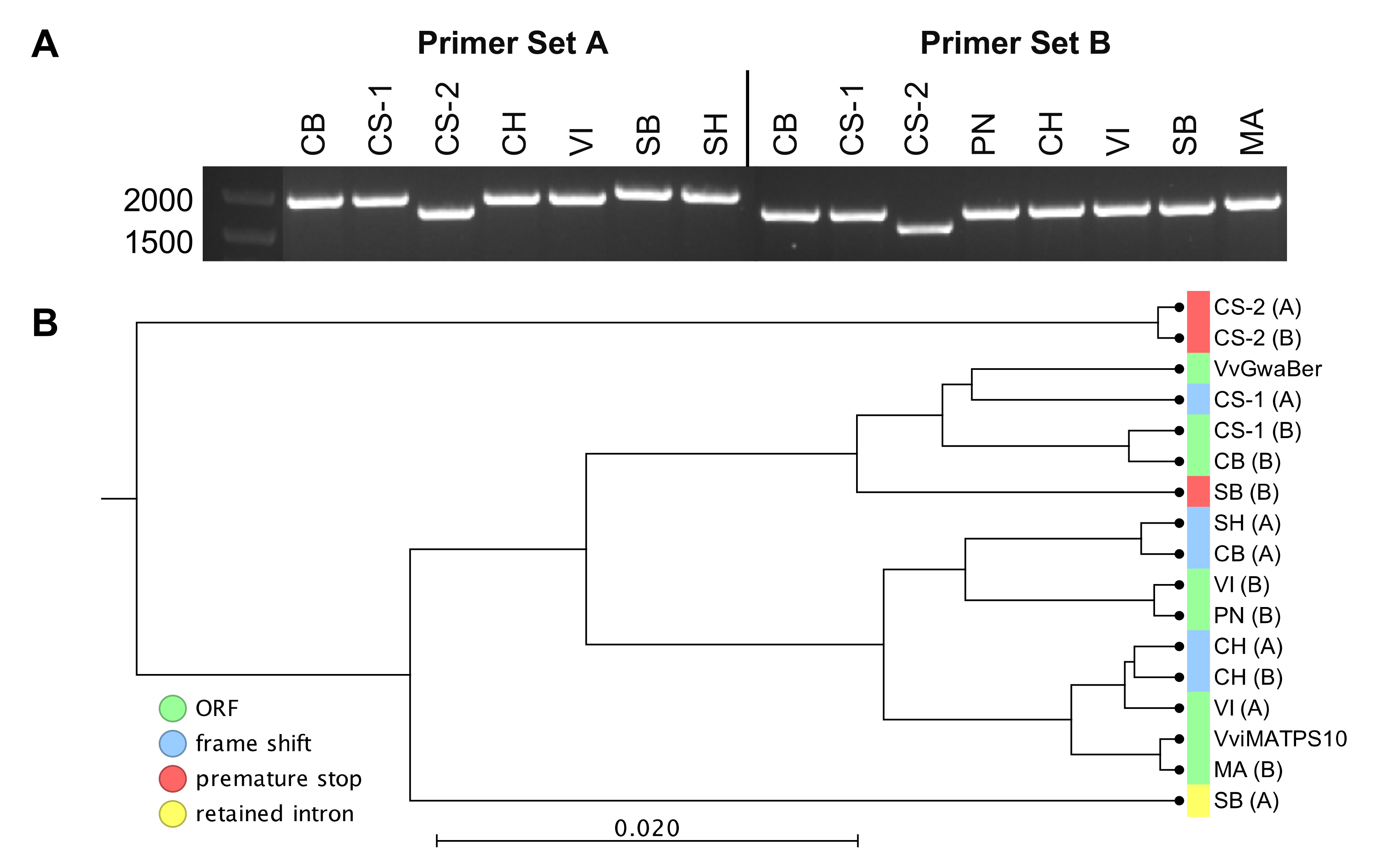
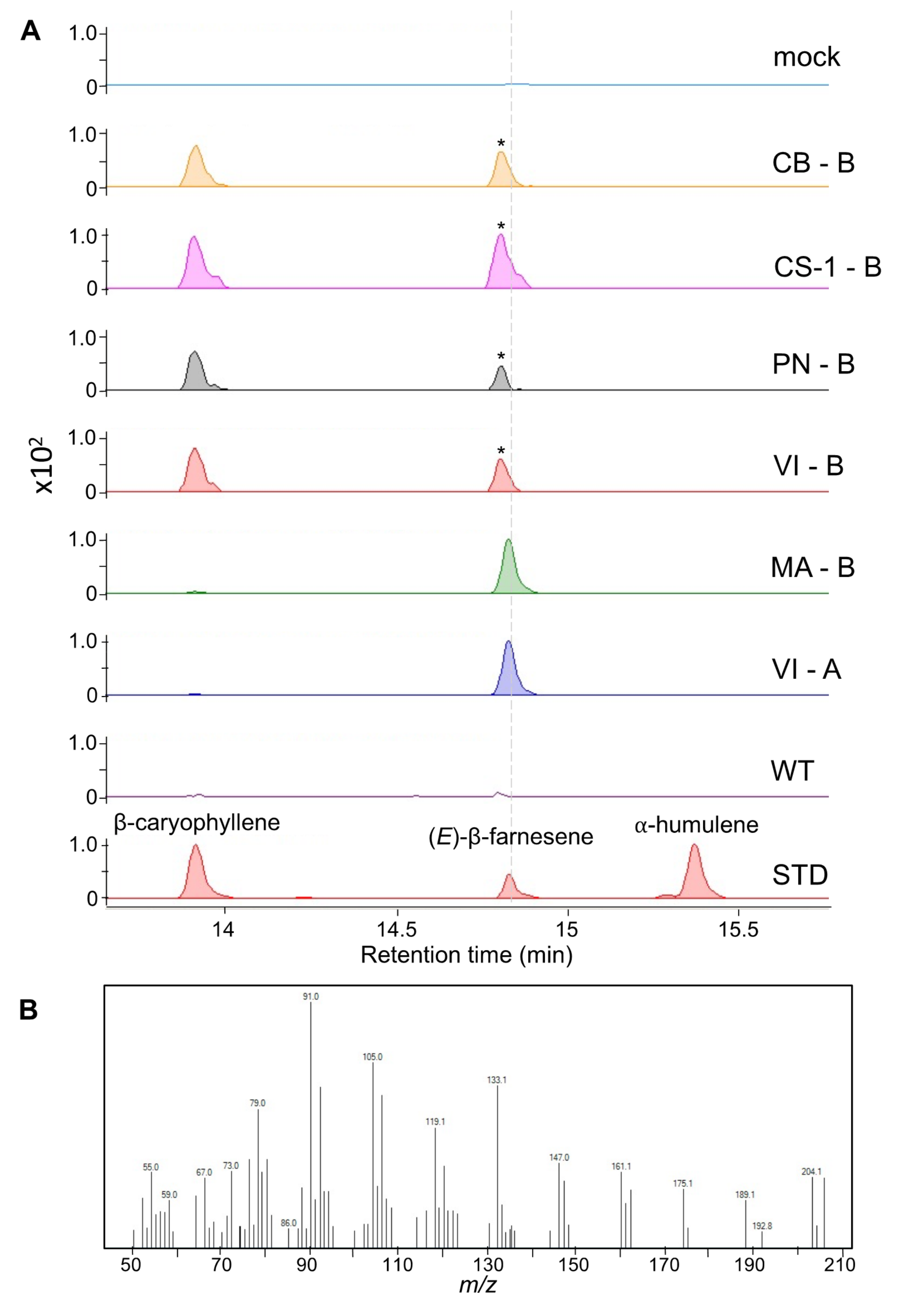
2.3. Transient Expression of fl-ORFs of VviTPS10 in Tobacco
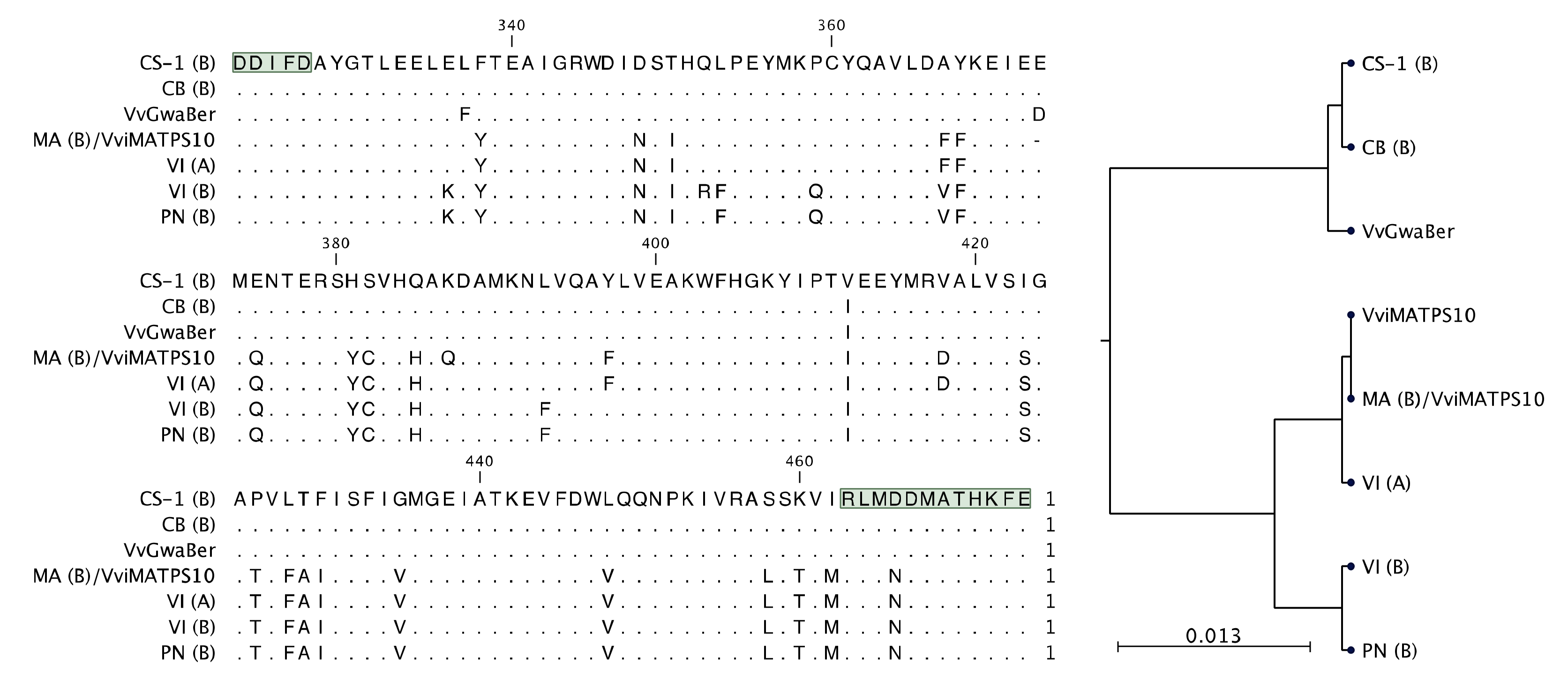
2.4. Sequence–Function Relationships of Isolated Paralogues
3. Discussion
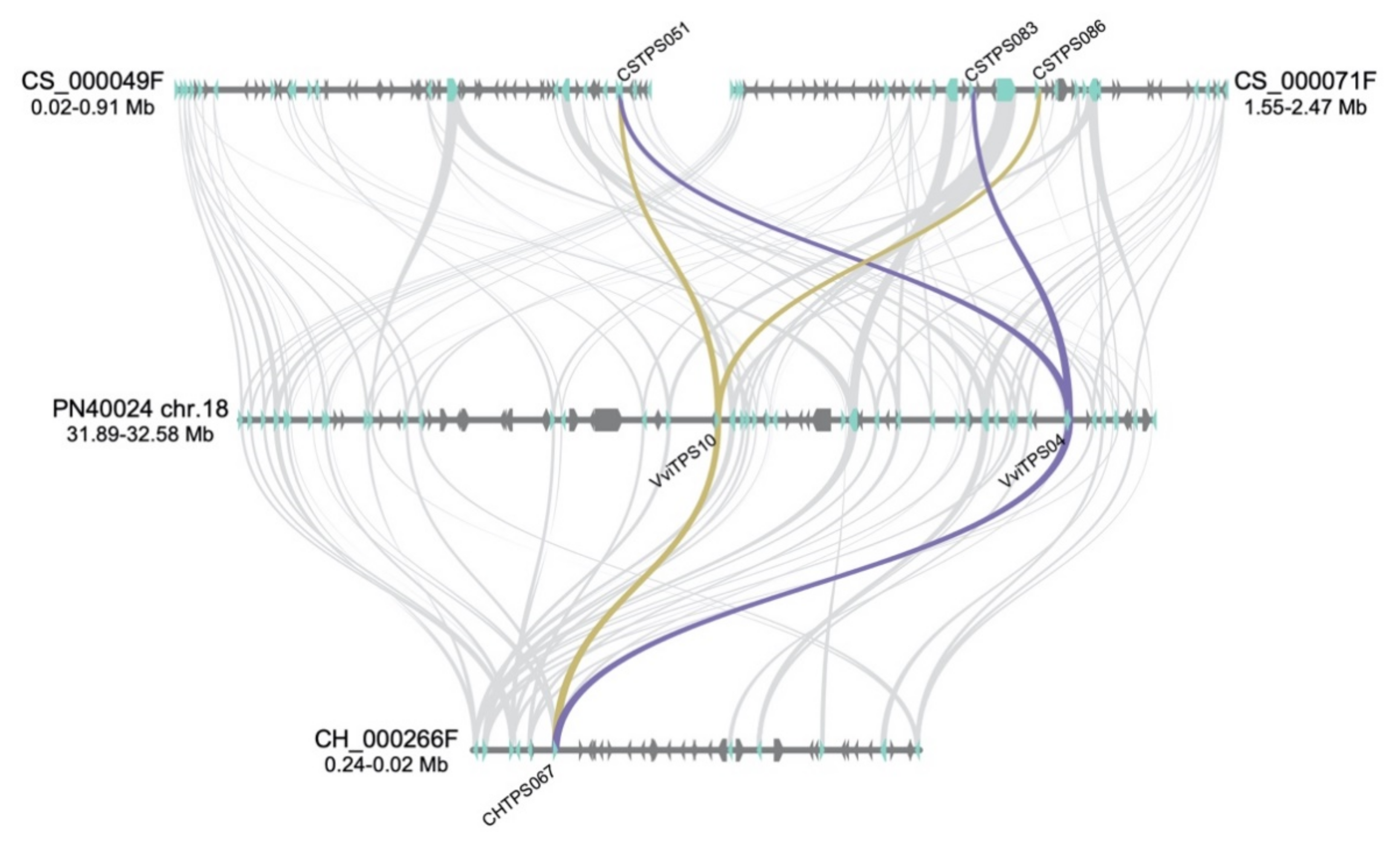
3.1. Genotype-Specific Structural Variation for Isolated VviTPS Paralogues
3.2. The Impact of Domestication on VviTPS Expansion
3.3. VviTPS04 and -10 Functions: An Ecophysiological Perspective
4. Conclusions
5. Materials and Methods
5.1. Identification of Putative Paralogues Using Diploid Grapevine Genomes
5.2. Identification of Putative VviTPS04 and -10 Structural Differences Using Whole-Genome Resequencing Data
5.3. Isolation and Cloning of VviTPS04 and -10 Paralogues
5.4. Sequence Analysis of Isolated Paralogues
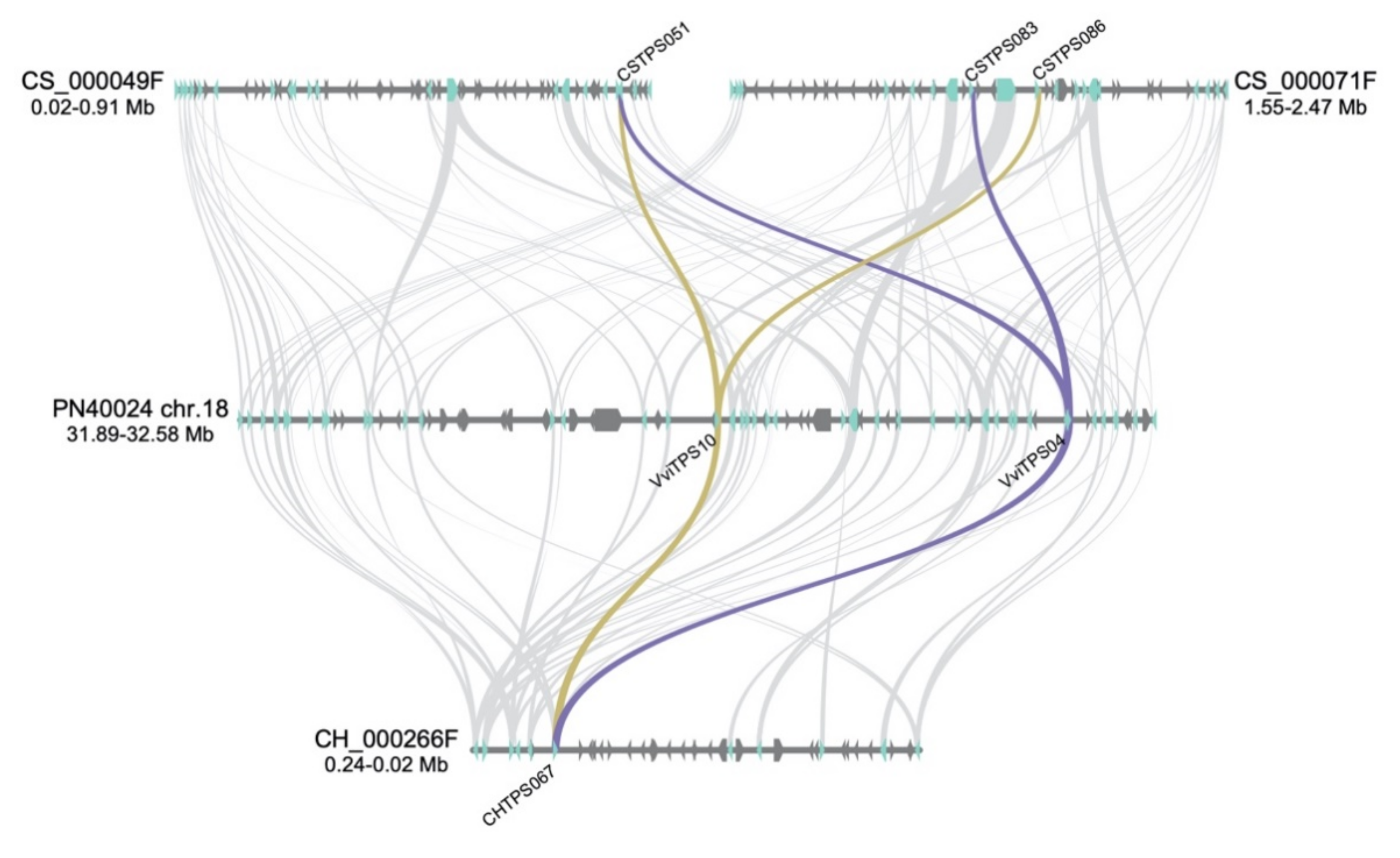
5.5. Gene Synteny Analysis
5.6. Agrobacterium-Mediated Transient Expression
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaillon, O.; Aury, J.-M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef]
- Lin, J.; Massonnet, M.; Cantu, D. The genetic basis of grape and wine aroma. Hortic. Res. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, C.A.; Parker, M.; Siebert, T.E.; Capone, D.L.; Francis, I.L. Terpenoids and their role in wine flavour: Recent advances. Aust. J. Grape Wine Res. 2015, 21. [Google Scholar] [CrossRef]
- Skinkis, P.A.; Bordelon, B.P.; Wood, K.V. Comparison of monoterpene constituents in Traminette, Gewürztraminer, and Riesling winegrapes. Am. J. Enol. Vitic. 2008, 59, 440–445. [Google Scholar]
- Wood, C.; Siebert, T.E.; Parker, M.; Capone, D.L.; Elsey, G.M.; Pollnitz, A.P.; Eggers, M.; Meier, M.; Vössing, T.; Widder, S.; et al. From wine to pepper: Rotundone, an obscure sesquiterpene, is a potent spicy aroma compound. J. Agric. Food Chem. 2008, 56, 3738–3744. [Google Scholar] [CrossRef] [PubMed]
- Kalua, C.M.; Boss, P.K. Evolution of volatile compounds during the development of cabernet sauvignon grapes (Vitis vinifera L.). J. Agric. Food Chem. 2009, 57, 3818–3830. [Google Scholar] [CrossRef]
- Martin, D.; Aubourg, S.; Schouwey, M.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.; Bohlmann, J. Functional Annotation, Genome Organization and Phylogeny of the Grapevine (Vitis vinifera) Terpene Synthase Gene Family Based on Genome Assembly, FLcDNA Cloning, and Enzyme Assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [Green Version]
- Bloch, K.; Chaykin, S.; Phillips, A.; De Waard, A. Mevalonic acid pyrophospahte and isopentenylpyrphosphate. J. Biol. Chem. 1959, 234, 2595–2604. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef]
- Rohmer, M. A Mevalonate-independent Route to Isopentenyl Diphosphate. In Comprehensive Natural Product Chemistry. Isoprenoids Including Carotenoids and Steroids; Cane, D.E., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 2, pp. 45–65. ISBN 0-08-043154-2. [Google Scholar]
- Davis, E.M.; Croteau, R. Cyclization enzymes in the biosynthesis of monoterpenes, sesquiterpenes, and diterpenes. Top. Curr. Chem. 2000, 209, 53–95. [Google Scholar]
- Martin, D.M.; Bohlmann, J. Identification of Vitis vinifera (-)-α-terpineol synthase by in silico screening of full-length cDNA ESTs and functional characterization of recombinant terpene synthase. Phytochemistry 2004, 65, 1223–1229. [Google Scholar] [CrossRef]
- Yoshikuni, Y.; Ferrin, T.E.; Keasling, J.D. Designed divergent evolution of enzyme function. Nature 2006, 440, 1078–1082. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-X.; Fang, X.; Zhao, Q.; Ruan, J.-X.; Yang, C.-Q.; Wang, L.-J.; Miller, D.J.; Faraldos, J.A.; Allemann, R.K.; Chen, X.-Y.; et al. Rational engineering of plasticity residues of sesquiterpene synthases from Artemisia annua: Product specificity and catalytic efficiency. Biochem. J. 2013, 451, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leferink, N.G.H.; Ranaghan, K.E.; Karuppiah, V.; Currin, A.; van der Kamp, M.W.; Mulholland, A.J.; Scrutton, N.S. Experiment and Simulation Reveal How Mutations in Functional Plasticity Regions Guide Plant Monoterpene Synthase Product Outcome. ACS Catal. 2018, 3780–3791. [Google Scholar] [CrossRef] [PubMed]
- Cane, D.E. Enzymic formation of sesquiterpenes. Chem. Rev. 1990, 90, 1089–1103. [Google Scholar] [CrossRef]
- Christianson, D.W. Structural biology and chemistry of the terpenoid cyclases. Chem. Rev. 2006, 106, 3412–3442. [Google Scholar] [CrossRef] [PubMed]
- Wedler, H.; Pemberton, R.; Tantillo, D. Carbocations and the complex flavor and bouquet of wine: Mechanistic aspects of terpene biosynthesis in wine grapes. Molecules 2015, 20, 10781–10792. [Google Scholar] [CrossRef] [Green Version]
- Drew, D.P.; Andersen, T.B.; Sweetman, C.; Møller, B.L.; Ford, C.; Simonsen, H.T. Two key polymorphisms in a newly discovered allele of the Vitis vinifera TPS24 gene are responsible for the production of the rotundone precursor α-guaiene. J. Exp. Bot. 2015, 67, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Smit, S.J.; Vivier, M.A.; Young, P.R. Linking terpene synthases to sesquiterpene metabolism in grapevine flowers. Front. Plant Sci. 2019, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Canaguier, A.; Grimplet, J.; Gaspero, G.D.; Scalabrin, S.; Duchêne, E.; Choisne, N. A new version of the grapevine reference genome assembly (12X.v2) and of its annotation (VCost.v3). Genom. Data 2017, 14, 56–62. [Google Scholar] [CrossRef]
- Figueroa-Balderas, R.; Minio, A.; Morales-Cruz, A.; Vondras, A.M.; Cantu, D. Strategies for sequencing and assembling grapevine genomes. In The Grape Genome; Cantu, D., Walker, M.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 77–88. [Google Scholar]
- Minio, A.; Massonnet, M.; Figueroa-Balderas, R.; Vondras, A.M.; Blanco-Ulate, B.; Cantu, D. Iso-Seq Allows Genome-Independent Transcriptome Profiling of Grape Berry Development. G3 Genes Genomes Genet. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, M.J.; Johnson, D.L.; Bohlmann, J.; van Vuuren, H.J.J.; Jones, S.J.M.; Pretorius, I.S.; Schmidt, S.A.; Borneman, A.R. Population sequencing reveals clonal diversity and ancestral inbreeding in the grapevine cultivar Chardonnay. PLoS Genet. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minio, A.; Massonnet, M.; Figueroa-Balderas, R.; Castro, A.; Cantu, D. Diploid Genome Assembly of the Wine Grape Carménère. G3 Genes Genomes Genet. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.; Peluso, P.; Sedlazeck, F.J.; Nattestad, M.; Concepcion, G.T.; Clum, A.; Dunn, C.; O’Malley, R.; Figueroa-Balderas, R.; Morales-Cruz, A.; et al. Phased diploid genome assembly with single-molecule real-time sequencing. Nat. Methods 2016, 13, 1050–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, S.J.; Vivier, M.A.; Young, P.R. Comparative (within species) genomics of the Vitis vinifera L. terpene synthase family to explore the impact of genotypic variation using phased diploid genomes. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Carretero-Paulet, L.; Librado, P.; Chang, T.H.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Rozas, J.; Albert, V.A. High gene family turnover rates and gene space adaptation in the compact genome of the carnivorous plant utricularia gibba. Mol. Biol. Evol. 2015, 32, 1284–1295. [Google Scholar] [CrossRef] [Green Version]
- Del Pozo, J.C.; Ramirez-Parra, E. Whole genome duplications in plants: An overview from Arabidopsis. J. Exp. Bot. 2015, 66, 6991–7003. [Google Scholar] [CrossRef] [Green Version]
- Giannuzzi, G.; D’Addabbo, P.; Gasparro, M.; Martinelli, M.; Carelli, F.N.; Antonacci, D.; Ventura, M. Analysis of high-identity segmental duplications in the grapevine genome. BMC Genom. 2011, 12, 436. [Google Scholar] [CrossRef] [Green Version]
- Aradhya, M.K.; Dangl, G.S.; Prins, B.H.; Boursiquot, J.-M.; Walker, M.A.; Meredith, C.P.; Simon, C.J. Genetic structure and differentiation in cultivated grape, Vitis vinifera L. Genet. Res. 2003, 81, 179–192. [Google Scholar] [CrossRef] [Green Version]
- Salmaso, M.; Faes, G.; Segala, C.; Stefanini, M.; Salakhutdinov, I.; Zyprian, E.; Toepfer, R.; Grando, M.S.; Velasco, R. Genome diversity and gene haplotypes in the grapevine (Vitis vinifera L.), as revealed by single nucleotide polymorphisms. Mol. Breed. 2004, 14, 385–395. [Google Scholar] [CrossRef]
- Laucou, V.; Launay, A.; Bacilieri, R.; Lacombe, T.; Adam-Blondon, A.F.; Bérard, A.; Chauveau, A.; De Andrés, M.T.; Hausmann, L.; Ibáñez, J.; et al. Extended diversity analysis of cultivated grapevine Vitis vinifera with 10K genome-wide SNPs. PLoS ONE 2018, 13, e192540. [Google Scholar] [CrossRef]
- Zhou, Y.; Muyle, A.; Gaut, B.S. Evolutionary genomics and the domestication of grapes. In The Grape Genome; Cantu, D., Walker, M.A., Eds.; Springer International Publishing: Cham, Swizterland, 2019; pp. 39–55. [Google Scholar]
- Falara, V.; Akhtar, T.A.; Nguyen, T.T.H.H.; Spyropoulou, E.A.; Bleeker, P.M.; Schauvinhold, I.; Matsuba, Y.; Bonini, M.E.; Schilmiller, A.L.; Last, R.L.; et al. The tomato terpene synthase gene family. Plant Physiol. 2011, 157, 770–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Köllner, T.G.; Li, G.; Wei, G.; Chen, X.; Zeng, D.; Qian, Q.; Chen, F. Combinatorial evolution of a terpene synthase gene cluster explains terpene variations in oryza. Plant Physiol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Duan, S.; Sheng, J.; Zhu, S.; Ni, X.; Shao, J.; Liu, C.; Nick, P.; Du, F.; Fan, P.; et al. Whole-genome resequencing of 472 Vitis accessions for grapevine diversity and demographic history. Nat. Commun. 2019, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.H.; Gu, Z.; Wang, H.; Nekrutenko, A. Evolutionary analyses of the human genome. Nature 2001, 409, 847–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dueholm, B.; Drew, D.P.; Sweetman, C.; Simonsen, H.T. In planta and in silico characterization of five sesquiterpene synthases from Vitis vinifera (cv. Shiraz) berries. Planta 2019, 249, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Myles, S.; Chia, J.M.; Hurwitz, B.; Simon, C.; Zhong, G.Y.; Buckler, E.; Ware, D. Rapid genomic characterization of the genus Vitis. PLoS ONE 2010, 5, e0008219. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Massonnet, M.; Sanjak, J.S.; Cantu, D.; Gaut, B.S. Evolutionary genomics of grape (Vitis vinifera ssp. vinifera) domestication. Proc. Natl. Acad. Sci. USA 2017, 114, 11715–11720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabidze, V.; Pipia, I.; Gogniashvili, M.; Kunelauri, N.; Ujmajuridze, L.; Pirtskhalava, M.; Vishnepolsky, B.; Hernandez, A.G.; Fields, C.J.; Beridze, T. Whole genome comparative analysis of four Georgian grape cultivars. Mol. Genet. Genom. 2017, 292, 1377–1389. [Google Scholar] [CrossRef]
- Mercenaro, L.; Nieddu, G.; Porceddu, A.; Pezzotti, M.; Camiolo, S. Sequence polymorphisms and structural variations among four grapevine (Vitis vinifera L.) cultivars representing sardinian agriculture. Front. Plant Sci. 2017, 8, 1279. [Google Scholar] [CrossRef] [Green Version]
- Di Genova, A.; Almeida, A.M.; Muñoz-Espinoza, C.; Vizoso, P.; Travisany, D.; Moraga, C.; Pinto, M.; Hinrichsen, P.; Orellana, A.; Maass, A. Whole genome comparison between table and wine grapes reveals a comprehensive catalog of structural variants. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [Green Version]
- Keller, M. Botany and anatomy. In The Science of Grapevines: Anatomy and Physiology; Academic Press: San Diego, CA, USA, 2010; pp. 1–47. ISBN 978-0-12-374881-2. [Google Scholar]
- Kronenberg, Z.N.; Rhie, A.; Koren, S.; Concepcion, G.T.; Peluso, P.; Munson, K.M.; Hiendleder, S.; Fedrigo, O.; Jarvis, E.D.; Adam, M.; et al. Extended haplotype phasing of de novo genome assemblies with FALCON-Phase. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2018, 220, 692–702. [Google Scholar] [CrossRef]
- Myles, S.; Boyko, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F.; Aradhya, M.K.; Prins, B.; Reynolds, A.; Chia, J.-M.; Ware, D.; et al. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. USA 2011, 108, 3530–3535. [Google Scholar] [CrossRef] [Green Version]
- Carbonell-Bejerano, P.; Royo, C.; Torres-Pérez, R.; Grimplet, J.; Fernandez, L.; Franco-Zorrilla, J.M.; Lijavetzky, D.; Baroja, E.; Martínez, J.; García-Escudero, E.; et al. Catastrophic unbalanced genome rearrangements cause somatic loss of berry color in grapevine. Plant Physiol. 2017, 175, 786–801. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Gao, Z.; Tao, J.; Jiang, W.; Zhang, S.; Wang, Q.; Qu, S. Genome-wide detection of SNP and SV variations to reveal early ripening-related genes in grape. PLoS ONE 2016, 11, e0147749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardone, M.F.; D’Addabbo, P.; Alkan, C.; Bergamini, C.; Catacchio, C.R.; Anaclerio, F.; Chiatante, G.; Marra, A.; Giannuzzi, G.; Perniola, R.; et al. Inter-varietal structural variation in grapevine genomes. Plant J. 2016, 88. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.S.; Minio, A.; Massonnet, M.; Solares, E.A.; Lyu, Y.; Beridze, T.; Cantu, D.; Gaut, B.S. Structural variants, clonal propagation, and genome evolution in grapevine (Vitis vinifera). bioRxiv 2018. [Google Scholar] [CrossRef]
- Purugganan, M.D. Evolutionary insights into the nature of plant domestication. Curr. Biol. 2019, 29, R705–R714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmo, H. Selecting and breeding new grape varieties. Calif. Agric. 1980, 34, 23–24. [Google Scholar]
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; Hoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar] [CrossRef]
- Tasin, M.; Bäckman, A.C.; Bengtsson, M.; Ioriatti, C.; Witzgall, P. Essential host plant cues in the grapevine moth. Naturwissenschaften 2006, 93, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Tasin, M.; Betta, E.; Carlin, S.; Gasperi, F.; Mattivi, F.; Pertot, I. Volatiles that encode host-plant quality in the grapevine moth. Phytochemistry 2011, 72, 1999–2005. [Google Scholar] [CrossRef]
- Salvagnin, U.; Malnoy, M.; Thöming, G.; Tasin, M.; Carlin, S.; Martens, S.; Vrhovsek, U.; Angeli, S.; Anfora, G. Adjusting the scent ratio: Using genetically modified Vitis vinifera plants to manipulate European grapevine moth behaviour. Plant Biotechnol. J. 2018, 16, 264–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addison, P. Ecology and Management of Lobesia vanillana in Vineyards. Available online: https://www.wineland.co.za/new-winetech-projects-2017-part-2/ (accessed on 17 December 2019).
- Picaud, S.; Brodelius, M.; Brodelius, P.E. Expression, purification and characterization of recombinant (E)-β-farnesene synthase from Artemisia annua. Phytochemistry 2005, 66, 961–967. [Google Scholar] [CrossRef]
- Dèrozier, S.; Samson, F.; Tamby, J.-P.; Guichard, C.; Brunaud, V.; Grevet, P.; Gagnot, S.; Label, P.; Leplé, J.-C.; Lecharny, A.; et al. Exploration of plant genomes in the FLAGdb++ environment. Plant Methods 2011, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boratyn, G.M.; Thierry-Mieg, J.; Thierry-Mieg, D.; Busby, B.; Madden, T.L. Magic-BLAST, an accurate RNA-seq aligner for long and short reads. BMC Bioinform. 2019, 20. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R.; Teeling, E. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coombe, B.G. Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 100–110. [Google Scholar] [CrossRef]
- Peyret, H.; Lomonossoff, G.P. The pEAQ vector series: The easy and quick way to produce recombinant proteins in plants. Plant Mol. Biol. 2013, 83, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, F.; Thuenemann, E.C.; Lomonossoff, G.P. pEAQ: Versatile expression vectors for easy and quick transient expression of heterologous proteins in plants. Plant Biotechnol. J. 2009, 7, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acid Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Bowers, J.E.; Wang, X.; Ming, R.; Alam, M.; Paterson, A.H. Synteny and collinearity in plant genomes. Science 2008, 320, 486–488. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Krishnakumar, V.; Li, J. jcvi: JCVI utility libraries. Zenodo 2015. [Google Scholar] [CrossRef]
- Bach, S.S.; Bassard, J.-É.; Andersen-Ranberg, J.; Møldrup, M.E.; Simonsen, H.T.; Hamberger, B. High-throughput testing of terpenoid biosynthesis candidate genes using transient expression in Nicotiana benthamiana. In Plant Isoprenoids: Methods and Protocols; Rodríguez-Concepción, M., Ed.; Springer Science: New York, NY, USA, 2014; Volume 1153, pp. 245–255. ISBN 978-1-4939-0605-5. [Google Scholar]
- Williams, C.; Buica, A. Comparison of an offline-SPE-GC-MS and online-HS-SPME-GC-MS method for the analysis of volatile terpenoids in wine. Molecules 2019, 25, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| gDNA Model | Genome | Cabernet Franc (TA-183) | Carménère (TA-254) | Cabernet Sauvignon (TA-280) | Chardonnay (TA-238) | Merlot (TA-241) | Muscat of Alexandria (TA-170) | Pinot Noir (TA-379) | Riesling (TA-329) | Sangiovese (TA-334) | Sauvignon Blanc (TA-417) | Syrah (TA-242) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CHTPS045 | CH | A | A | |||||||||
| CHTPS046 | CH | A | A | A | A | C | A | A | A | A | A | A |
| CHTPS062 | CH | B | B | B | ||||||||
| CHTPS065 | CH | C | A | A | A | A | ||||||
| CHTPS067 | CH | B | C | C | C | B | ||||||
| CHTPS147 | CH | A | A | A | A | A | A | A | A | A | A | |
| CHTPS159 | CH | B | ||||||||||
| CHTPS173 | CH | C | C | C | C | C | C | C | C | C | C | C |
| CRTPS019 | CR | A | A | A | A | A | A | A | A | |||
| CRTPS081 | CR | A | A | C | A | C | A | A | A | |||
| CRTPS102 | CR | C | ||||||||||
| CRTPS105 | CR | |||||||||||
| CRTPS106 | CR | A | A | A | A | A | C | A | C | |||
| CRTPS151 | CR | A | A | A | A | A | A | A | A | C | A | |
| CRTPS173 | CR | A | ||||||||||
| CRTPS199 | CR | A | A | A | A | |||||||
| CRTPS200 | CR | B | C | C | B | |||||||
| CSTPS036 | CS | A | A | A | A | A | ||||||
| CSTPS040 | CS | A | A | A | A | A | A | A | A | C | A | |
| CSTPS043 | CS | A | ||||||||||
| CSTPS047 | CS | C | B | C | C | |||||||
| CSTPS051 | CS | B | B | B | ||||||||
| CSTPS059 | CS | A | A | C | ||||||||
| CSTPS079 | CS | A | A | C | A | A | ||||||
| CSTPS080 | CS | A | A | A | A | A | A | A | A | |||
| CSTPS083 | CS | C | ||||||||||
| CSTPS086 | CS | B | B | |||||||||
| CSTPS090 | CS | C | A | A | ||||||||
| VviTPS04 | PN40024 | B | B | B | B | |||||||
| VviTPS10 | PN40025 | C | A | C | A | A | C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smit, S.J.; Vivier, M.A.; Young, P.R. Seeing the Forest through the (Phylogenetic) Trees: Functional Characterisation of Grapevine Terpene Synthase (VviTPS) Paralogues and Orthologues. Plants 2021, 10, 1520. https://doi.org/10.3390/plants10081520
Smit SJ, Vivier MA, Young PR. Seeing the Forest through the (Phylogenetic) Trees: Functional Characterisation of Grapevine Terpene Synthase (VviTPS) Paralogues and Orthologues. Plants. 2021; 10(8):1520. https://doi.org/10.3390/plants10081520
Chicago/Turabian StyleSmit, Samuel J., Melané A. Vivier, and Philip R. Young. 2021. "Seeing the Forest through the (Phylogenetic) Trees: Functional Characterisation of Grapevine Terpene Synthase (VviTPS) Paralogues and Orthologues" Plants 10, no. 8: 1520. https://doi.org/10.3390/plants10081520
APA StyleSmit, S. J., Vivier, M. A., & Young, P. R. (2021). Seeing the Forest through the (Phylogenetic) Trees: Functional Characterisation of Grapevine Terpene Synthase (VviTPS) Paralogues and Orthologues. Plants, 10(8), 1520. https://doi.org/10.3390/plants10081520