
Soil Moisture Levels Affect the Anatomy and Mechanical Properties of Basil Stems (Ocimum basilicum L.)




Abstract
1. Introduction
2. Results
2.1. Substrate Matric Potential during Cultivation
2.2. Xylem Anatomical Traits
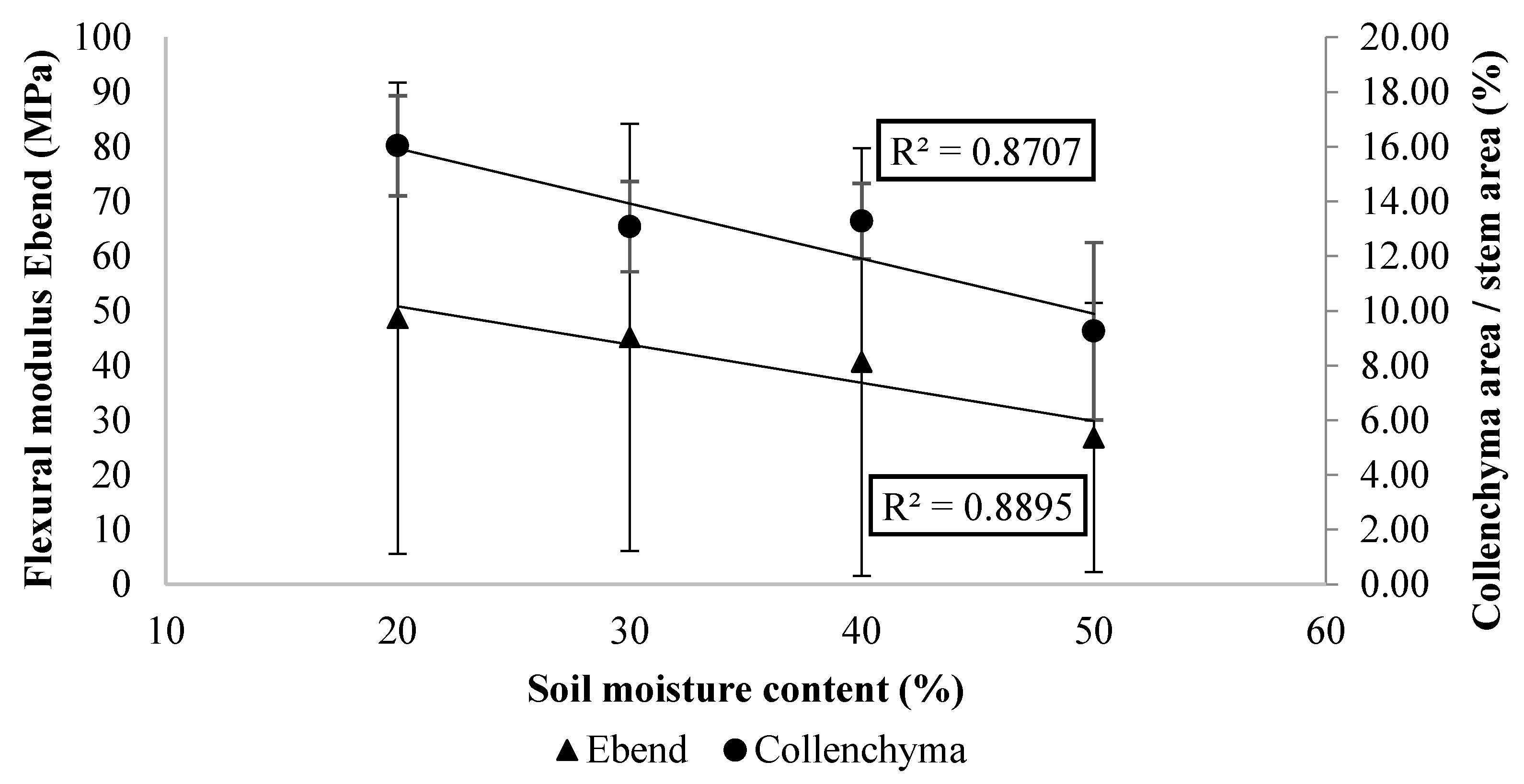
2.3. Collenchyma Anatomical Traits and Mechanical Strength of Apical Stem Segments
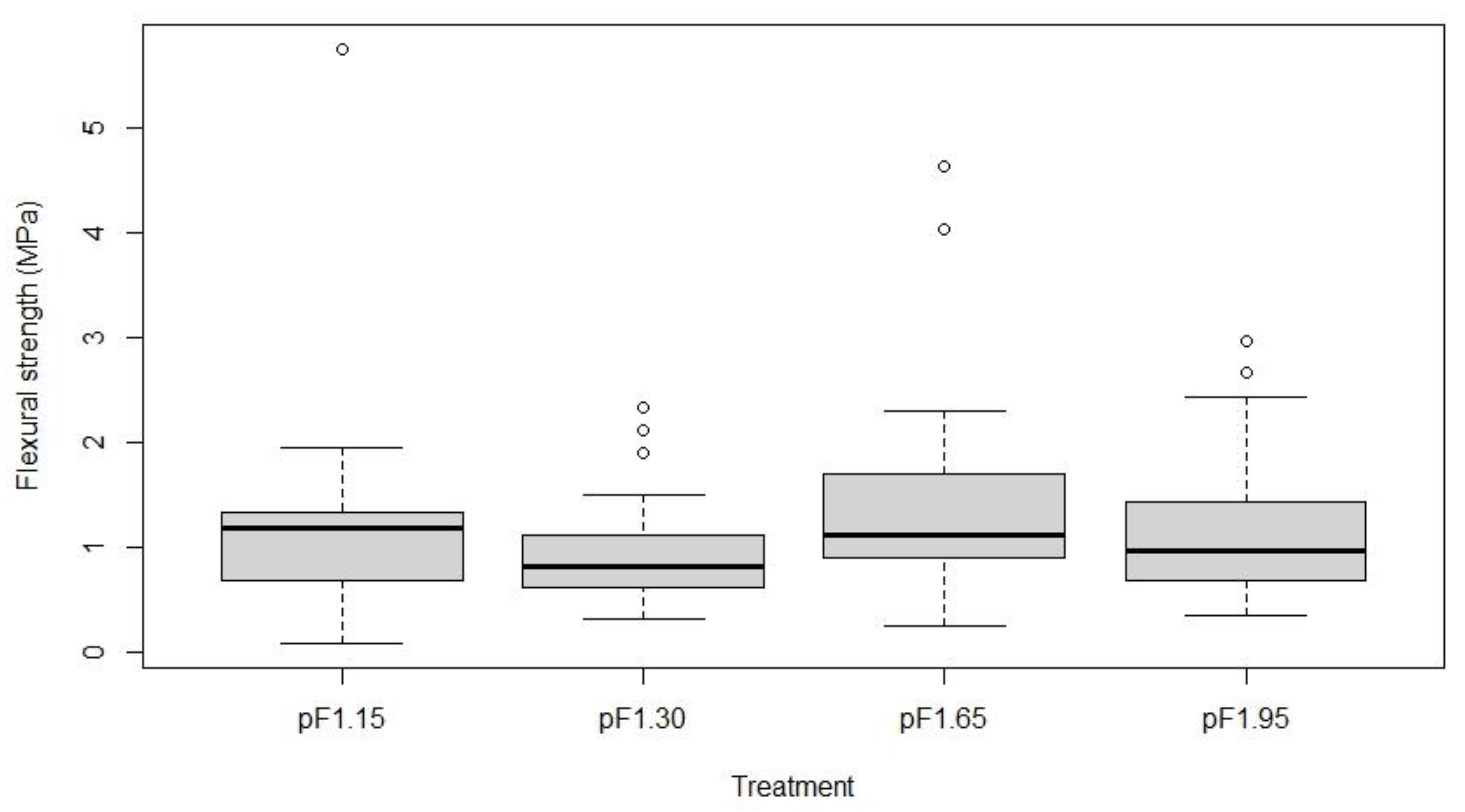
2.4. Tracheid and Sclerenchyma Anatomical Traits and Mechanical Strength of Basal Stem Segments
3. Discussion
3.1. Soil Water Availability Influences Xylem Anatomical Traits
3.2. Collenchyma and Sclerenchyma Formation Changes with Soil Water Availability
3.3. Stem Anatomical Structures Contribute to its Mechanical Strength against Bending
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Monitoring Soil Moisture Content
4.3. 3-Point Bending Tests
- Ebend—Flexural modulus, bending modulus or modulus of elasticity (MOE) (MPa)
- L—distance between the two metal supports (mm)
- Fmax—maximum load or bending force (N)
- d—diameter of the stem sample (mm)
- δ—deflection (mm) at maximum force
4.4. Biometric and Anatomical Measurements
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Růžička, K.; Ursache, R.; Hejatko, J.; Helariutta, Y. Xylem development—From the cradle to the grave. New Phytol. 2015, 207, 519–535. [Google Scholar] [CrossRef]
- Kim, H.K.; Park, J.; Hwang, I. Investigating water transport through the xylem network in vascular plants. J. Exp. Bot. 2014, 65, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Fournier, M.; Dlouhá, J.; Jaouen, G.; Almeras, T. Integrative biomechanics for tree ecology: Beyond wood density and strength. J. Exp. Bot. 2013, 64, 4793–4815. [Google Scholar] [CrossRef]
- Irigoyen, J.; Goicoechea, N.; Antolín, M.; Pascual, I.; Sánchez-Díaz, M.; Aguirreolea, J.; Morales, F. Growth, photosynthetic acclimation and yield quality in legumes under climate change simulations: An updated survey. Plant Sci. 2014, 226, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Qaderi, M.M.; Martel, A.B.; Dixon, S.L. Environmental Factors Influence Plant Vascular System and Water Regulation. Plants 2019, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Montesano, F.F.; van Iersel, M.; Boari, F.; Cantore, V.; D’Amato, G.; Parente, A. Sensor-based irrigation management of soilless basil using a new smart irrigation system: Effects of set-point on plant physiological responses and crop performance. Agric. Water Manag. 2018, 203, 20–29. [Google Scholar] [CrossRef]
- Fereres, E.; Goldhamer, D.A.; Parsons, L.R. Irrigation Water Management of Horticultural Crops. HortScience 2003, 38, 1036–1042. [Google Scholar] [CrossRef]
- Wagner, K.R.; Ewers, F.W.; Davis, S.D. Tradeoffs between hydraulic efficiency and mechanical strength in the stems of four co-occurring species of chaparral shrubs. Oecologia 1998, 117, 53–62. [Google Scholar] [CrossRef]
- Boyce, C.K.; Zwieniecki, M.; Cody, G.D.; Jacobsen, C.; Wirick, S.; Knoll, A.H.; Holbrook, N.M. Evolution of xylem lignification and hydrogel transport regulation. Proc. Natl. Acad. Sci. USA 2004, 101, 17555–17558. [Google Scholar] [CrossRef]
- Tyree, M.T.; Davis, S.D.; Cochard, H. Biophysical Perspectives of Xylem Evolution: Is there a Tradeoff of Hydraulic Efficiency for Vulnerability to Dysfunction? IAWA J. 1994, 15, 335–360. [Google Scholar] [CrossRef]
- Niklas, K.J. Plant Biomechanics. An Engineering Approach to Plant Form and Function; University of Chicago: Chicago, IL, USA, 1992. [Google Scholar] [CrossRef]
- Bociąg, K.; Gałka, A.; Łazarkiewicz, T.; Szmeja, J. Mechanical strength of stems in aquatic macrophytes. Acta Soc. Bot. Pol. 2011, 78, 181–187. [Google Scholar] [CrossRef]
- Caliaro, M.; Schmich, F.; Speck, T.; Speck, O. Effect of drought stress on bending stiffness in petioles ofCaladium bicolor(Araceae). Am. J. Bot. 2013, 100, 2141–2148. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Leroux, O. Collenchyma: A versatile mechanical tissue with dynamic cell walls. Ann. Bot. 2012, 110, 1083–1098. [Google Scholar] [CrossRef]
- Leroux, O.; Eder, M.; Saxe, F.; Dunlop, J.W.C.; Popper, Z.A.; Viane, R.L.L.; Knox, J.P. Comparative in situ analysis reveals the dynamic nature of sclerenchyma cell walls of the fern Asplenium rutifolium. Ann. Bot. 2017, 121, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Esau, K. Ontogeny and structure of collenchyma and of vascular tissues in celery petioles. Hilgardia 1936, 10, 429–476. [Google Scholar] [CrossRef][Green Version]
- Karam, G.; Gibson, L. Biomimicking of animal quills and plant stems: Natural cylindrical shells with foam cores. Mater. Sci. Eng. C 1994, 2, 113–132. [Google Scholar] [CrossRef]
- Shah, D.U.; Reynolds, T.; Ramage, M.H. The strength of plants: Theory and experimental methods to measure the mechanical properties of stems. J. Exp. Bot. 2017, 68, 4497–4516. [Google Scholar] [CrossRef] [PubMed]
- Hales, T.C.; Miniat, C.F. Soil moisture causes dynamic adjustments to root reinforcement that reduce slope stability. Earth Surf. Process. Landforms 2017, 42, 803–813. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B.; Fourcaud, T.; Dumail, J.-F.; Van Beek, R. The Influence of Cellulose Content on Tensile Strength in Tree Roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Rossi, S.; DesLauriers, A.; Anfodillo, T. Assessment of Cambial Activity and Xylogenesis by Microsampling Tree Species: An Example at the Alpine Timberline. IAWA J. 2006, 27, 383–394. [Google Scholar] [CrossRef]
- Pinto, C.A.; David, J.; Cochard, H.; Caldeira, M.C.; Henriques, M.O.; Quilhó, T.; Paço, T.A.; Pereira, J.; David, T.S. Drought-induced embolism in current-year shoots of two Mediterranean evergreen oaks. For. Ecol. Manag. 2012, 285, 1–10. [Google Scholar] [CrossRef]
- February, E. Sensitivity of xylem vessel size and frequency to rainfall and temperature: Implications for palaeontology, Palaeontol. Africana 1993, 30, 91–95. [Google Scholar]
- Arend, M.; Fromm, J. Seasonal change in the drought response of wood cell development in poplar. Tree Physiol. 2007, 27, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Mencuccini, M.; Grace, J.; Fioravanti, M. Biomechanical and hydraulic determinants of tree structure in Scots pine: Anatomical characteristics. Tree Physiol. 1997, 17, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Cochard, H. Cavitation in trees. Comptes Rendus Phys. 2006, 7, 1018–1026. [Google Scholar] [CrossRef]
- Fonti, P.; von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.S.; Fridley, J.D.; Yin, J.; Bauerle, T.L. Contrasting xylem vessel constraints on hydraulic conductivity between native and non-native woody understory species. Front. Plant Sci. 2013, 4, 486. [Google Scholar] [CrossRef]
- González, I.G.; Eckstein, D. Climatic signal of earlywood vessels of oak on a maritime site. Tree Physiol. 2003, 23, 497–504. [Google Scholar] [CrossRef]
- Pritchard, J. The control of cell expansion in roots. New Phytol. 1994, 127, 3–26. [Google Scholar] [CrossRef]
- Grafi, G.; Florentin, A.; Ransbotyn, V.; Morgenstern, Y. The Stem Cell State in Plant Development and in Response to Stress. Front. Plant Sci. 2011, 2, 53. [Google Scholar] [CrossRef]
- Grafi, G.; Chalifa-Caspi, V.; Nagar, T.; Plaschkes, I.; Barak, S.; Ransbotyn, V. Plant response to stress meets dedifferentiation. Planta 2011, 233, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Olmos, E.; Sánchez-Blanco, M.J.; Ferrández, T.; Alarcon, J.J. Subcellular Effects of Drought Stress inRosmarinus officinalis. Plant Biol. 2007, 9, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Badel, E.; Ewers, F.W.; Cochard, H.; Telewski, F.W. Acclimation of mechanical and hydraulic functions in trees: Impact of the thigmomorphogenetic process. Front. Plant Sci. 2015, 6, 266. [Google Scholar] [CrossRef] [PubMed]
- Sperry, J.S.; Hacke, U.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.L.; Ewers, F.W.; Pratt, R.B.; Paddock, W.A.; Davis, S.D. Do Xylem Fibers Affect Vessel Cavitation Resistance? Plant Physiol. 2005, 139, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.B.; Jacobsen, A.; Ewers, F.W.; Davis, S.D. Relationships among xylem transport, biomechanics and storage in stems and roots of nine Rhamnaceae species of the California chaparral. New Phytol. 2007, 174, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Pratt, R.B.; Jacobsen, A. Conflicting demands on angiosperm xylem: Tradeoffs among storage, transport and biomechanics. Plant Cell Environ. 2017, 40, 897–913. [Google Scholar] [CrossRef] [PubMed]
- Schenkels, L.; Saeys, W.; Lauwers, A.; De Proft, M.P. Green light induces shade avoidance to alter plant morphology and increases biomass production in Ocimum basilicum L. Sci. Hortic. 2020, 261, 109002. [Google Scholar] [CrossRef]
- Niklas, K.J. Nodal Septa and the Rigidity of Aerial Shoots of Equisetum hyemale. Am. J. Bot. 1989, 76, 521. [Google Scholar] [CrossRef]
- Evans, L.S.; Kahn-Jetter, Z.; Marks, C.; Harmoney, K.R. Mechanical Properties and Anatomical Components of Stems of 42 Grass Species. J. Torrey Bot. Soc. 2007, 134, 458–467. [Google Scholar] [CrossRef]
- Al-Zube, L.; Sun, W.; Robertson, D.; Cook, D. The elastic modulus for maize stems. Plant Methods 2018, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | n | Mean Vessel Diameter (µm) | Vessel Frequency per mm2 | Percent Vessel Area per Transverse Stem area (%) |
|---|---|---|---|---|
| Apical stem segment | ||||
| pF1.95 | 794 | 15.42 ± 1.62 (B) | 83 ± 17 (A) | 1.25 ± 0.20 (A) |
| pF1.65 | 625 | 20.51 ± 1.68 (A) | 49 ± 7 (B) | 1.39 ± 0.16 (A) |
| pF1.30 | 551 | 19.13 ± 1.98 (A) | 55 ± 11 (B) | 1.29 ± 0.22 (A) |
| pF1.15 | 523 | 19.93 ± 3.25 (A) | 48 ± 12 (B) | 1.11 ± 0.17 (A) |
| Basal stem segment | ||||
| pF1.95 | 3534 | 25.29 ± 2.39 (B) | 40 ± 5 (A) | 1.99 ± 0.43 (C) |
| pF1.65 | 3573 | 33.65 ± 2.60 (A) | 29 ± 4 (B) | 2.57 ± 0.43 (A) |
| pF1.30 | 3129 | 34.25 ± 3.20 (A) | 24 ± 3 (C) | 2.19 ± 0.33 (BC) |
| pF1.15 | 3482 | 34.73 ± 3.51 (A) | 26 ± 6 (BC) | 2.41 ± 0.46 (AB) |
| Treatment | n | Flexural Modulus Ebend (MPa) | Flexural Strength σ (MPa) |
|---|---|---|---|
| pF1.95 | 60 | 48.08 ± 43.07 (a) | 0.36 ± 0.23 (a) |
| pF1.65 | 56 | 45.14 ± 39.04 (a) | 0.34 ± 0.19 (a) |
| pF1.30 | 53 | 40.67 ± 39.10 (ab) | 0.29 ± 0.21 (ab) |
| pF1.15 | 56 | 26.82 ± 24.59 (b) | 0.27 ± 0.21 (b) |
| Treatment | n | Tracheid Area per Stem Area (%) | Mean Tracheid Diameter (µm) | Sclerenchyma per Stem Area (%) | Mean Sclerenchyma Diameter (µm) |
|---|---|---|---|---|---|
| pF1.95 | 20 | 18.14 ± 7.60 (a) | 10.53 ± 3.46 (a) | 3.54 ± 0.81 (a) | 20.95 ± 5.87 (a) |
| pF1.65 | 20 | 18.31 ± 8.78 (a) | 10.89 ± 3.83 (a) | 3.33 ± 0.85 (a) | 20.20 ± 6.06 (a) |
| pF1.30 | 20 | 15.94 ± 5.69 (a) | 11.02 ± 3.49 (a) | 3.29 ± 0.46 (a) | 22.62 ± 6.65 (a) |
| pF1.15 | 20 | 19.56 ± 9.54 (a) | 10.81 ± 3.44 (a) | 3.46 ± 0.44 (a) | 22.64 ± 6.80 (a) |
| Parameters | Tracheid Area per Stem Area (%) | Sclerenchyma per Stem Area (%) | Vessel Frequency per mm2 | Vessel Diameter (µm) | Percent Vessel Area Per Stem Area (%) |
|---|---|---|---|---|---|
| VWC (%) | 0.03 | −0.05 | −0.68 | 0.67 | 0.21 |
| Flexural modulus Ebend (MPa) | 0.76 | 0.06 | 0.18 | −0.15 | 0.00 |
| Flexural strength σ (MPa) | 0.86 | 0.10 | 0.21 | −0.04 | 0.27 |
| Irrigation Set-Point (%) | Calculated Irrigation Thresholds (%) | Actual Average Volumetric Water Content of the Substrate during the 5-Week Experiment (%) 1st Replication | Actual Average Volumetric Water Content of the Substrate during the 5-Week Experiment (%) 2nd Replication |
|---|---|---|---|
| 20 | 11 | 26 ± 4 | 21 ± 9 |
| 30 | 22 | 31 ± 5 | 32 ± 8 |
| 40 | 34 | 44 ± 9 | 39 ± 13 |
| 50 | 45 | 49 ± 8 | 51 ± 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Driesen, E.; De Proft, M.; Saeys, W. Soil Moisture Levels Affect the Anatomy and Mechanical Properties of Basil Stems (Ocimum basilicum L.). Plants 2021, 10, 1320. https://doi.org/10.3390/plants10071320
Driesen E, De Proft M, Saeys W. Soil Moisture Levels Affect the Anatomy and Mechanical Properties of Basil Stems (Ocimum basilicum L.). Plants. 2021; 10(7):1320. https://doi.org/10.3390/plants10071320
Chicago/Turabian StyleDriesen, Elisa, Maurice De Proft, and Wouter Saeys. 2021. "Soil Moisture Levels Affect the Anatomy and Mechanical Properties of Basil Stems (Ocimum basilicum L.)" Plants 10, no. 7: 1320. https://doi.org/10.3390/plants10071320
APA StyleDriesen, E., De Proft, M., & Saeys, W. (2021). Soil Moisture Levels Affect the Anatomy and Mechanical Properties of Basil Stems (Ocimum basilicum L.). Plants, 10(7), 1320. https://doi.org/10.3390/plants10071320