
Distribution and Genetic Diversity of Grapevine Viruses in Russia


Abstract
1. Introduction
2. Results
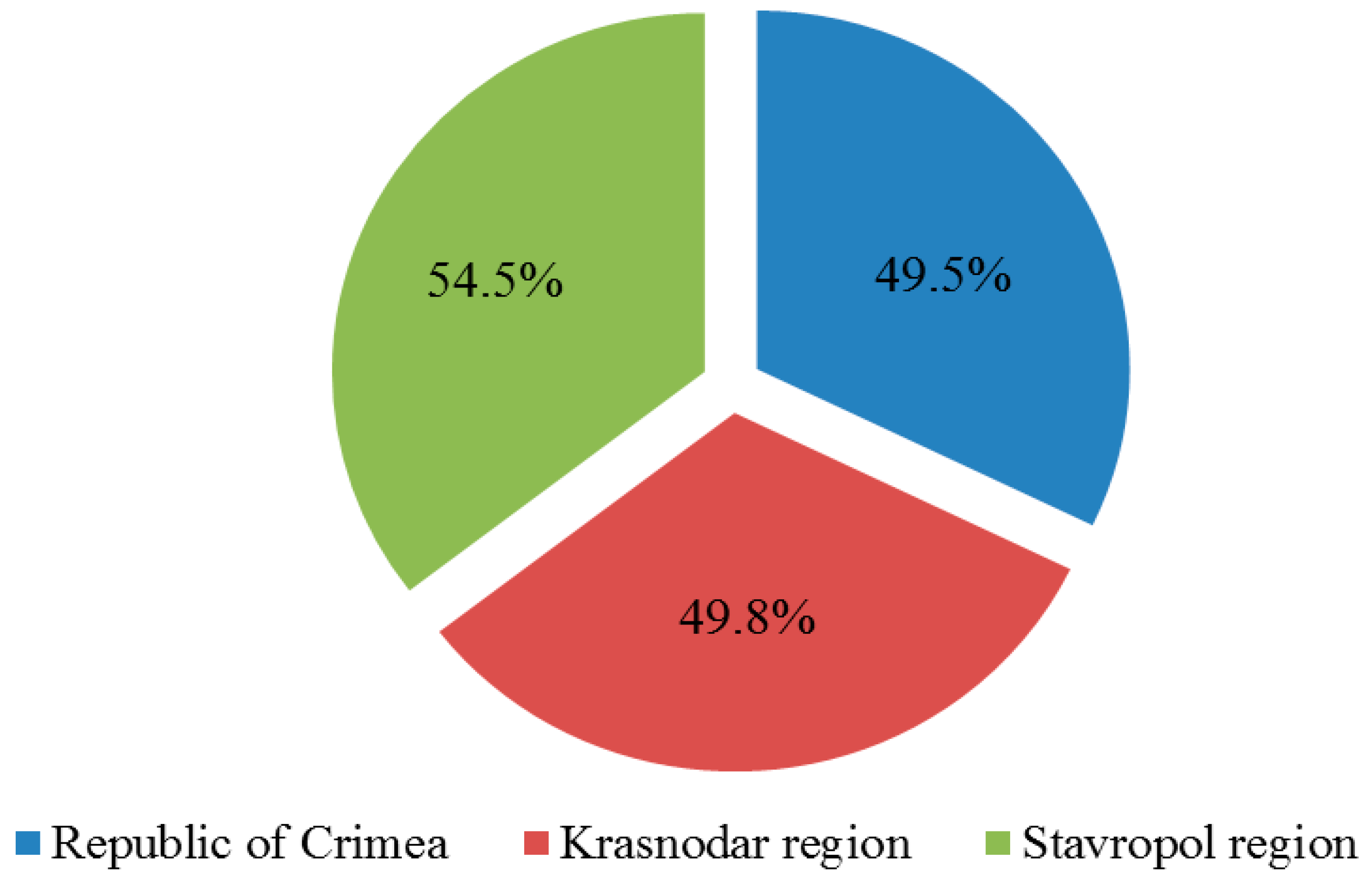
2.1. Phytosanitary Monitoring
2.2. Phylogenetic Studies and Determination of Molecular Groups of Viruses
2.3. Viruses of Rugose Wood Complex
2.4. Fleck Complex and Grapevine Fanleaf Viruses
2.5. Grapevine Leafroll Disease Viruses
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Detection of Grapevine Viruses
4.3. Phylogenetic Studies
4.4. Recombination Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maliogka, V.I.; Martelli, G.P.; Fuchs, M.; Katis, N.I. Control of viruses infecting grapevine. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2015; Volume 91, pp. 175–227. [Google Scholar]
- OIV 2019 Report on the World Vitivinicultural Situation. Available online: http://www.oiv.int/en/oiv-life/oiv-2019-report-on-the-world-vitivinicultural-situation (accessed on 23 April 2021).
- Federal State Statistics Service. Available online: https://rosstat.gov.ru/search (accessed on 19 March 2021).
- Atallah, S.S.; Gomez, M.I.; Fuchs, M.F.; Martinson, T.E. Economic Impact of Grapevine Leafroll Disease on Vitis Vinifera Cv. Cabernet Franc in Finger Lakes Vineyards of New York. Am. J. Enol. Vitic. 2012, 63, 73–79. [Google Scholar] [CrossRef]
- Naidu, R.; Rowhani, A.; Fuchs, M.; Golino, D.; Martelli, G.P. Grapevine Leafroll: A Complex Viral Disease Affecting a High-Value Fruit Crop. Plant Dis. 2014, 98, 1172–1185. [Google Scholar] [CrossRef]
- Fuchs, M. Grapevine Viruses: A Multitude of Diverse Species with Simple but Overall Poorly Adopted Management Solutions in the Vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Martelli, G.P. An Overview on Grapevine Viruses, Viroids, and the Diseases They Cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 31–46. ISBN 978-3-319-57706-7. [Google Scholar]
- Angelini, E.; Aboughanem, N.; Dolja, V.; Meng, B. Grapevine leafroll-associated virus 2. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 141–165. ISBN 978-3-319-57704-3. [Google Scholar]
- Beuve, M.; Sempé, L.; Lemaire, O. A Sensitive One-Step Real-Time RT-PCR Method for Detecting Grapevine Leafroll-Associated Virus 2 Variants in Grapevine. J. Virol. Methods 2007, 141, 117–124. [Google Scholar] [CrossRef]
- Naidu Grapevine leafroll-associated virus 1. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 127–139. ISBN 978-3-319-57704-3.
- Reynolds, A. The Grapevine, Viticulture, and Winemaking: A Brief Introduction. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 3–29. ISBN 978-3-319-57704-3. [Google Scholar]
- Alabi, O.J.; Casassa, L.F.; Gutha, L.R.; Larsen, R.C.; Henick-Kling, T.; Harbertson, J.F.; Naidu, R.A. Impacts of Grapevine Leafroll Disease on Fruit Yield and Grape and Wine Chemistry in a Wine Grape (Vitis Vinifera L.) Cultivar. PLoS ONE 2016, 11, e0149666. [Google Scholar] [CrossRef] [PubMed]
- Bertamini, M.; Malossini, U.; Muthuchelian, K.; Nedunchezhian, N. Physiological Response of Field Grown Grapevine (Vitis Vinifera L. Cv. Marzemino) to Grapevine Leafroll-Associated Virus (GLRaV-1). Phytopathol. Mediterr. 2005, 44, 10. [Google Scholar]
- Jones, T.J.; Rayapati, N.A.; Nita, M. Occurrence of Grapevine Leafroll Associated Virus-2, −3 and Grapevine Fleck Virus in Virginia, U.S.A., and Factors Affecting Virus Infected Vines. Eur. J. Plant Pathol. 2015, 142, 209–222. [Google Scholar] [CrossRef]
- Sabanadzovic, S.; Aboughanem-Sabanadzovic, N.; Martelli, G.P. Grapevine fleck and similar viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 331–349. ISBN 978-3-319-57706-7. [Google Scholar]
- Dmitrenko, U.D.; Porotikova, E.V.; Gorislavets, S.M.; Risovannaya, V.I.; Volkov, Y.A.; Stranishevskaya, E.P.; Kamionskaya, A.M.; Vinogradova, S.V. First Report of Grapevine Rupestris Stem Pitting-Associated Virus in Russia. Plant Dis. 2016, 100, 2542. [Google Scholar] [CrossRef]
- Porotikova, E.V.; Dmitrenko, U.D.; Volodin, V.A.; Volkov, Y.A.; Gorislavets, S.M.; Stranishevskaya, E.P.; Risovannaya, V.I.; Kamionskaya, A.M.; Vinogradova, S.V. First Report of Grapevine Virus A in Russian Grapevines. Plant Dis. 2016, 100, 2541. [Google Scholar] [CrossRef]
- Porotikova, E.V.; Risovannaya, V.I.; Volkov, Y.A.; Dmitrenko, U.D.; Volodin, V.A.; Gorislavets, S.M.; Stranishevskaya, E.P.; Agranovsky, A.A.; Kamionskaya, A.M.; Vinogradova, S.V. Occurrence of Grapevine Leafroll-Associated Viruses-1 and -3 in Crimea. Mosc. Univ. Biol. Sci. Bull. 2016, 71, 76–79. [Google Scholar] [CrossRef][Green Version]
- Porotikova, E.V.; Dmitrenko, U.D.; Yurchenko, E.G.; Vinogradova, S.V. First Report of Grapevine Leafroll-Associated Virus 2 in Russian Grapevines (Vitis Vinifera). Plant Dis. 2018, 103, 164. [Google Scholar] [CrossRef]
- Risovannaya, V.; Volodin, V.; Volkov, Y.; Stranishevskaya, E.; Goryslavets, S. Mixed Infecting of Grapevine with Viruses in the Commercial Vineyards of the Crimean Peninsula. BIO Web Conf. 2020, 25, 06005. [Google Scholar] [CrossRef]
- Digiaro, M.; Elbeaino, T.; Martelli, G.P. Grapevine fanleaf virus and Other Old World Nepoviruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 47–82. ISBN 978-3-319-57706-7. [Google Scholar]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and Analysis of Recombination Patterns in Virus Genomes. Virus Evol. 2015, 1. [Google Scholar] [CrossRef]
- Sabella, E.; Pierro, R.; Luvisi, A.; Panattoni, A.; D’Onofrio, C.; Scalabrelli, G.; Nutricati, E.; Aprile, A.; De Bellis, L.; Materazzi, A. Phylogenetic Analysis of Viruses in Tuscan Vitis Vinifera Sylvestris (Gmeli) Hegi. PLoS ONE 2018, 13, e0200875. [Google Scholar] [CrossRef]
- Goszczynski, D.E. Rugose Wood-Associated Viruses Do Not Appear to Be Involved in Shiraz (Syrah) Decline in South Africa. Arch. Virol. 2010, 155, 1463–1469. [Google Scholar] [CrossRef]
- Hu, G.-J.; Dong, Y.-F.; Zhu, H.-J.; Zhang, Z.-P.; Fan, X.-D.; Ren, F.; Zhou, J. Molecular Characterizations of Two Grapevine Rupestris Stem Pitting-Associated Virus Isolates from China. Arch. Virol. 2015, 160, 2641–2645. [Google Scholar] [CrossRef]
- Meng, B.; Rowhani, A. Grapevine rupestris stem pitting-associated virus. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 257–287. ISBN 978-3-319-57706-7. [Google Scholar]
- Bertazzon, N.; Borgo, M.; Vanin, S.; Angelini, E. Genetic Variability and Pathological Properties of Grapevine Leafroll-Associated Virus 2 Isolates. Eur. J. Plant Pathol. 2010, 127, 185–197. [Google Scholar] [CrossRef]
- Jarugula, S.; Alabi, O.J.; Martin, R.R.; Naidu, R.A. Genetic Variability of Natural Populations of Grapevine Leafroll-Associated Virus 2 in Pacific Northwest Vineyards. Phytopathology® 2010, 100, 698–707. [Google Scholar] [CrossRef]
- Bertazzon, N.; Angelini, E. Advances in the Detection of Grapevine Leafroll-Associated Virus 2 Variants. J. Plant Pathol. 2004, 86, 283–290. [Google Scholar]
- Komar, V.; Vigne, E.; Demangeat, G.; Fuchs, M. Beneficial Effect of Selective Virus Elimination on the Performance of Vitis Vinifera Cv. Chardonnay. Am. J. Enol. Vitic. 2007, 58, 202–210. [Google Scholar]
- Mannini, F.; Digiaro, M. The Effects of Viruses and Viral Diseases on Grapes and Wine. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 453–482. ISBN 978-3-319-57704-3. [Google Scholar]
- Fajardo, T.V.M.; Kin, A.C.; Nickel, O.; Fajardo, T.V.M.; Kin, A.C.; Nickel, O. Molecular Variants of Grapevine Rupestris Stem Pitting-Associated Virus Infecting Grapevines (Vitis Spp.) in Brazil. Ciênc. Rural 2017, 47. [Google Scholar] [CrossRef]
- Selmi, I.; Pacifico, D.; Lehad, A.; Stigliano, E.; Crucitti, D.; Carimi, F.; Mahfoudhi, N. Genetic Diversity of Grapevine Rupestris Stem Pitting-Associated Virus Isolates from Tunisian Grapevine Germplasm. Plant Pathol. 2020, 69, 1051–1059. [Google Scholar] [CrossRef]
- Alkowni, R. Detection of Genomic Variations in Grapevine Rupestris Stem Pitting Associated Virus, Leading for Identification of New Virus Isolates and Developing of Molecular Tool for Their Specific Detection. Hebron Univ. Res. J. 2008, 3, 36–48. [Google Scholar]
- Selmi, I.; Lehad, A.; Pacifico, D.; Carimi, F.; Mahfoudhi, N. Prevalence and Genetic Diversity of Grapevine Virus A in Tunisia. Phytopathol. Mediterr. 2018, 57, 237–244. [Google Scholar]
- Predajňa, L.; Glasa, M. Partial Sequence Analysis of Geographically Close Grapevine Virus A Isolates Reveals Their High Regional Variability and an Intra-Isolate Heterogeneity. J. Phytopathol. 2016, 164, 427–431. [Google Scholar] [CrossRef]
- Moradi, R.; Koolivand, D.; Eini, O.; Hajizadeh, M. Phylogenetic Analysis of Two Iranian Grapevine Virus A Isolates Using Coat Protein Gene Sequence. IJGPB 2017, 6, 10. [Google Scholar]
- Balsak, S.C.; Buzkan, N. Prevalence and Genetic Variability of Grapevine Virus A in Turkish Autochthonous Grapevine Varieties. Arch. Virol. 2021, 166, 943–947. [Google Scholar] [CrossRef]
- Alabi, O.J.; Rwahnih, M.A.; Mekuria, T.A.; Naidu, R.A. Genetic Diversity of Grapevine Virus A in Washington and California Vineyards. Phytopathology 2014, 104, 548–560. [Google Scholar] [CrossRef]
- Predajňa, L.; Gažiová, A.; Holovičová, E.; Glasa, M. Analysis of a Short Genomic Region of Grapevine Leafroll-Associated Virus 1 (GLRaV-1) Reveals the Presence of Two Different Molecular Groups of Isolates in Slovakia. Acta Virol. 2013, 57, 353–356. [Google Scholar] [CrossRef]
- Elçi, E. Genomic Variability and Recombination Analysis of Grapevine Leafroll-Associated Virus-1 Isolates from Turkey. Tarım Bilim. Derg. 2019, 319–327. [Google Scholar] [CrossRef]
- Fan, X.; Hong, N.; Dong, Y.; Ma, Y.; Zhang, Z.P.; Ren, F.; Hu, G.; Zhou, J.; Wang, G. Genetic Diversity and Recombination Analysis of Grapevine Leafroll-Associated Virus 1 from China. Arch. Virol. 2015, 160, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Chooi, K.M.; Cohen, D.; Pearson, M.N. Generic and Sequence-Variant Specific Molecular Assays for the Detection of the Highly Variable Grapevine Leafroll-Associated Virus 3. J. Virol. Methods 2013, 189, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Farooq, A.B.U.; Ma, Y.; Wang, Z.; Zhuo, N.; Wenxing, X.; Wang, G.; Hong, N. Genetic Diversity Analyses Reveal Novel Recombination Events in Grapevine Leafroll-Associated Virus 3 in China. Virus Res. 2013, 171, 15–21. [Google Scholar] [CrossRef]
- Gouveia, P.; Santos, M.T.; Eiras-Dias, J.E.; Nolasco, G. Five Phylogenetic Groups Identified in the Coat Protein Gene of Grapevine Leafroll-Associated Virus 3 Obtained from Portuguese Grapevine Varieties. Arch. Virol. 2011, 156, 413–420. [Google Scholar] [CrossRef]
- Vončina, D.; Al Rwahnih, M.; Rowhani, A.; Gouran, M.; Almeida, R.P.P. Viral Diversity in Autochthonous Croatian Grapevine Cultivars. Plant Dis. 2017, 101, 1230–1235. [Google Scholar] [CrossRef]
- Glasa, M.; Predajňa, L.; Komínek, P. Grapevine Fleck Virus Isolates Split into Two Distinct Molecular Groups. J. Phytopathol. 2011, 159, 805–807. [Google Scholar] [CrossRef]
- Poojari, S.; Lowery, T.; Rott, M.; Schmidt, A.-M.; DeLury, N.; Boulé, J.; Úrbez-Torres, J.R. First Report and Prevalence of Grapevine Fleck Virus in Grapevines (Vitis Vinifera) in Canada. Plant Dis. 2015, 100, 1028. [Google Scholar] [CrossRef]
- Crnogorac, A.; Gašpar, M.; Davino, S.; Mandić, A.; Matić, S. First Report of Grapevine Fleck Virus in Vineyards of Bosnia and Herzegovina. J. Plant Pathol. 2020, 102, 1299. [Google Scholar] [CrossRef]
- Crnogorac, A.; Panno, S.; Mandić, A.; Gašpar, M.; Caruso, A.G.; Noris, E.; Davino, S.; Matić, S. Survey of five major grapevine viruses infecting Blatina and Žilavka cultivars in Bosnia and Herzegovina. PLoS ONE 2021, 16, e0245959. [Google Scholar] [CrossRef]
- Komínek, P. Distribution of Grapevine Viruses in Vineyards of the Czech Republic. J. Plant Pathol. 2008, 90, 357–358. [Google Scholar]
- Martelli, G.P. Directory of Virus and Virus-like Diseases of the Grapevine and Their Agents. J. Plant Pathol. 2014, 96, 1–136. [Google Scholar]
- Mekuria, T.A.; Gutha, L.R.; Martin, R.R.; Naidu, R.A. Genome Diversity and Intra-and Interspecies Recombination Events in Grapevine Fanleaf Virus. Phytopathology 2009, 99, 1394–1402. [Google Scholar] [CrossRef]
- Rott, M.; Jelkmann, W. Characterization and Detection of Several Filamentous Viruses of Cherry: Adaptation of an Alternative Cloning Method (DOP-PCR), and Modification of an RNA Extraction Protocol | SpringerLink. Available online: https://link.springer.com/article/10.1023/A:1011264400482 (accessed on 11 November 2020).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Gribaudo, I. Simultaneous Detection of Nine Grapevine Viruses by Multiplex Reverse Transcription-Polymerase Chain Reaction with Coamplification of a Plant RNA as Internal Control. Phytopathology 2006, 96, 1223–1229. [Google Scholar] [CrossRef]
- Minafra, A.; Hadidi, A. Sensitive Detection of Grapevine Virus A, B, or Leafroll-Associated III from Viruliferous Mealybugs and Infected Tissue by CDNA Amplification. J. Virol. Methods 1994, 47, 175–187. [Google Scholar] [CrossRef]
- Sanchez, F.; Chay, C.; Borja, M.J.; Rowhani, A.; Romero, J.; Bruening, G.; Ponz, F. CDNA Sequence of the Capsid Protein Gene and 3’ Untranslated Region of a Fanleaf Isolate of Grapevine Fanleaf Virus. Nucleic Acids Res. 1991, 19, 5440. [Google Scholar] [CrossRef][Green Version]
- Bashir, N.S.; Nourinejhad-Zarghani, S.; Hajizadeh, M. Status of Infection with Grapevine Fanleaf Virus in Vineyards of Iran and Molecular Characteristics of the Isolates. Res. Rev. Biosci. 2015, 10, 267–277. [Google Scholar]
- Aboughanem, N.; Sabanadzovic, S.; Castellano, M.A.; Boscia, D.; Martelli, G.P. Properties of a New Isolate of Grapevine Leafroll-Associated Virus 2. Vitis Geilweilerhof 2001, 39, 119–123. [Google Scholar]
- Naidu, R.A.; Mekuria, T.A. First Report of Grapevine Fleck Virus from Washington Vineyards. Plant Dis. 2010, 94, 784. [Google Scholar] [CrossRef]
- Fonseca, F.; Esteves, F.; Teixeira Santos, M.; Brazao, J.; Eiras-Dias, J.E. Genetic Variants of Grapevine Leafroll-Associated Virus 2 Infecting Portuguese Grapevine Cultivars. Phytopathol. Mediterr. 2016, 55. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Amplicons | Group of Isolates | ||
|---|---|---|---|---|
| GLR2CP1/GLR2CP2 | V2dCPf2/V2dCPrl | |||
| TaqI | TaqI | RsaI | ||
| 3 (MH074870) | I | B | B | 3 |
| 346 | I | B | E | unknown |
| 602 | II | A | A | 1a |
| 638 | I | B | B | 3 |
| 899 | II | A | A | 1a |
| 1853 | I | - | - | not determined |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porotikova, E.; Terehova, U.; Volodin, V.; Yurchenko, E.; Vinogradova, S. Distribution and Genetic Diversity of Grapevine Viruses in Russia. Plants 2021, 10, 1080. https://doi.org/10.3390/plants10061080
Porotikova E, Terehova U, Volodin V, Yurchenko E, Vinogradova S. Distribution and Genetic Diversity of Grapevine Viruses in Russia. Plants. 2021; 10(6):1080. https://doi.org/10.3390/plants10061080
Chicago/Turabian StylePorotikova, Elena, Uliana Terehova, Vitalii Volodin, Eugeniya Yurchenko, and Svetlana Vinogradova. 2021. "Distribution and Genetic Diversity of Grapevine Viruses in Russia" Plants 10, no. 6: 1080. https://doi.org/10.3390/plants10061080
APA StylePorotikova, E., Terehova, U., Volodin, V., Yurchenko, E., & Vinogradova, S. (2021). Distribution and Genetic Diversity of Grapevine Viruses in Russia. Plants, 10(6), 1080. https://doi.org/10.3390/plants10061080