
Selective Extraction of Bioactive Phenylethanoids from Digitalis obscura

 ,
, 
 ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. Digitalis Obscura Cardenolide-Free Extracts and Their Biological Effects
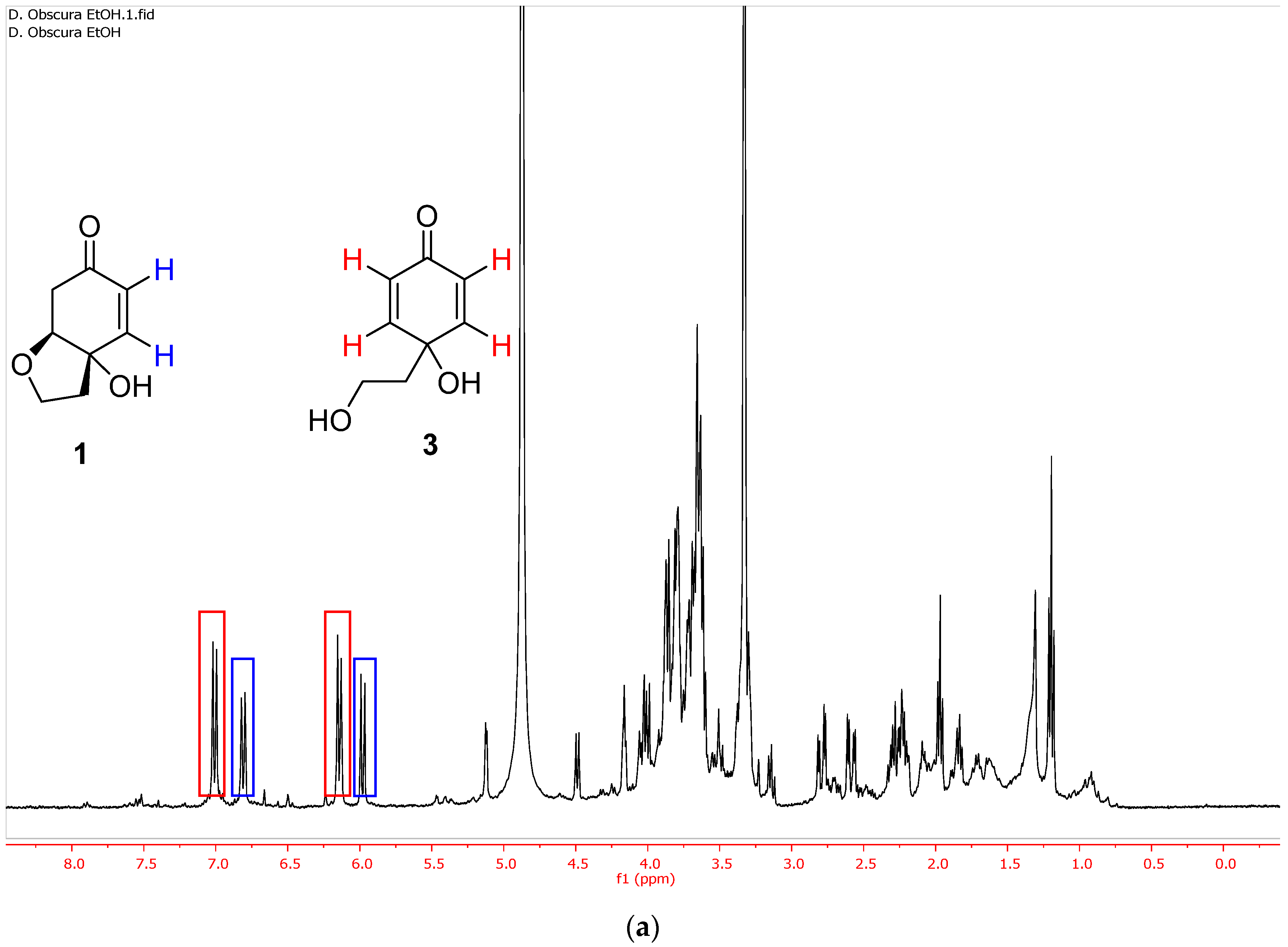
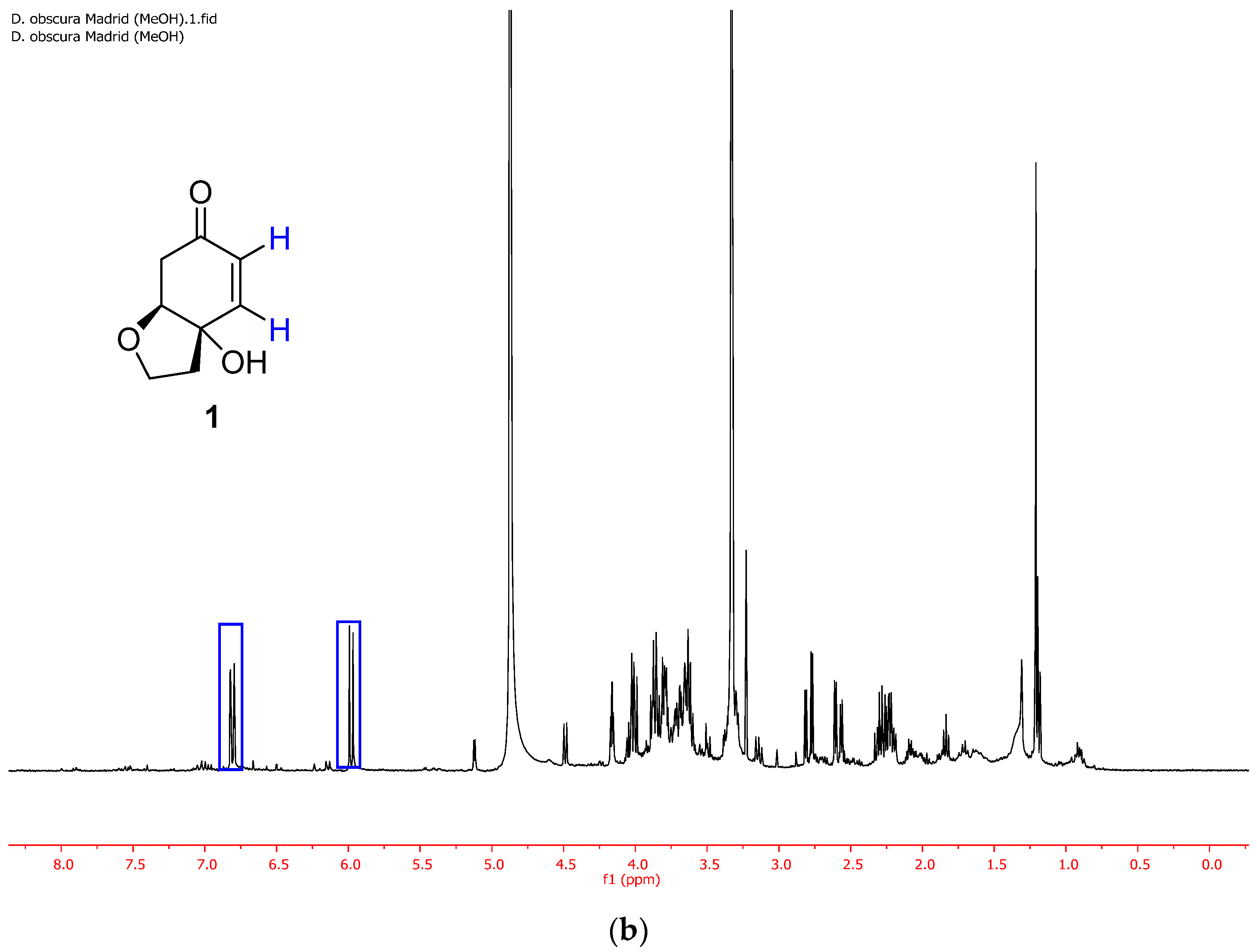
2.2. Selective Extraction of Rengyolone (1)
2.3. Selective Extraction of Cornoside Aglycon (3)


2.4. Bioactivity of Phenylethanoids
3. Conclusions
4. Experimental Section
4.1. Plant Material
4.2. Selective Extraction of Rengyolone
4.3. Selective Extraction of Compound 3
4.4. Antifeedant Activity
4.5. Nematicidal Bioassay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gerwick, B.C.; Sparks, T.C. Natural products for pest control: An analysis of their role, value and future. Pest Manag. Sci. 2014, 70, 1169–1185. [Google Scholar] [CrossRef] [PubMed]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef] [PubMed]
- Glare, T.; Caradus, J.; Gelernter, W.; Jackson, T.; Keyhani, N.; Köhl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Cantrell, C.L.; Dayan, F.E.; Duke, S.O. Natural products as sources for new pesticides. J. Nat. Prod. 2012, 75, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against insects—A review. Plant Prot. Sci. 2016, 52, 229–241. [Google Scholar] [CrossRef]
- Galisteo Pretel, A.; Pérez del Pulgar, H.; Olmeda, A.S.; Gonzalez-Coloma, A.; Barrero, A.F.; del Moral, Q.J.F. Novel Insect Antifeedant and Ixodicidal Nootkatone Derivatives. Biomolecules 2019, 9, 742. [Google Scholar] [CrossRef] [PubMed]
- Barrero, A.F.; del Moral, Q.J.F.; Gonzalez-Coloma, A.; Andres, M.F.; Diaz, C.E. Derivados de germacrona para el control de plagas. Spain Patent ES2020070509, 10 August 2020. [Google Scholar]
- Kreis, W. The foxgloves (Digitalis) revisited. Planta Med. 2017, 83, 962–976. [Google Scholar] [CrossRef]
- El-Mallakh, R.S.; Brar, K.S.; Yeruva, R.R. Cardiac glycosides in human physiology and disease: Update for Entomologists. Insects 2019, 10, 102. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Petschenka, G.; Bingham, R.A.; Weber, M.G.; Rasmann, S. Toxic cardenolides: Chemical ecology and coevolution of specialized plant–herbivore interactions. New Phytol. 2012, 194, 28–45. [Google Scholar] [CrossRef]
- Wang, X.-B.; Li, G.-H.; Zheng, L.-J.; Ji, K.-Y.; Lü, H.; Liu, F.-F.; Dang, L.-Z.; Mo, M.-H.; Zhang, K.-Q. Nematicidal cardenolides from Nerium indicum mill. Chem. Biodivers. 2009, 6, 431–436. [Google Scholar] [CrossRef]
- Taskova, R.M.; Gotfredsen, C.H.; Jensen, S.R. Chemotaxonomic markers in Digitalideae (Plantaginaceae). Phytochemistry 2005, 66, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Nishino, C.; Kobayashi, K. 3A-hydroxy- or Acetoxy-3,3a,7,7a-tetrahydro-6(2H)-benzofuranone as Anticancer Agent. Japan Patent JP 01290627A, 22 November 1989. [Google Scholar]
- Nishino, C.; Kobayashi, K. Tetrahydrobenzofurans as Antitumor Agents and Agrochemical Fungicides and Their Preparation. Japan Patent JP 01290666A, 22 November 1989. [Google Scholar]
- Nagakura, A.; Sato, M.; Kikuchi, M.; Nakatsu, T. Anticancer Formulations Containing an Oxabicyclononenone Derivative. Japan Patent JP 02200631A, 8 August 1990. [Google Scholar]
- Seo, J.Y.; Pyo, E.; Park, J.; Kim, J.-S.; Sung, S.H.; Oh, W.K. Nrf2-Mediated HO-1induction and antineuroinflammatory activities of halleridone. J. Med. Food 2017, 20, 1091–1099. [Google Scholar] [CrossRef]
- Tuntiwachwuttikul, P.; Rayanil, K.; Taylor, W.C. Chemical constituents from the flowers of Nyctanthes arbor-tristis. ScienceAsia 2003, 29, 21–30. [Google Scholar] [CrossRef]
- Phanthong, P.; Morales, N.P.; Chancharunee, S.; Mangmool, S.; Anantachoke, N.; Bunyapraphatsara, N. Biological activity of Dolichandrone serrulata flowers and their active components. Nat. Prod. Commun. 2015, 10, 1387–1390. [Google Scholar] [CrossRef] [PubMed]
- Kutluay, V.M.; Makino, T.; Inoue, M.; Saracoglu, I. New knowledge about old drugs; a cardenolide type glycoside with cytotoxic effect and unusual secondary metabolites from Digitalis grandiflora Miller. Fitoterapia 2019, 134, 73–80. [Google Scholar] [CrossRef]
- Benítez, G.; González-Tejero, M.R.; Molero-Mesa, J. Knowledge of ethnoveterinary medicine in the Province of Granada, Andalusia, Spain. J. Ethnopharmacol. 2012, 139, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Roca-Pérez, L.; Boluda, R.; Gavidia, I.; Pérez-Bermúdez, P. Seasonal cardenolide production and Dop5βr gene expression in natural populations of Digitalis obscura. Phytochemistry 2004, 65, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Huber, K.; Meyer, K. Die Glykoside der Blätter von Digitalis obscura L. ssp. obscura. Helv. Chim. Acta 1966, 49, 259–272. [Google Scholar] [CrossRef]
- Verma, S.K.; Das, A.K.; Cingoz, G.S.; Gurel, E. In vitro culture of Digitalis L. (Foxglove) and the production of cardenolides: An up-to-date review. Ind. Crops Prod. 2016, 94, 20–51. [Google Scholar] [CrossRef]
- Albuquerque, B.R.; Prieto, M.A.; Barreiro, M.F.; Rodrigues, A.; Curran, T.P.; Barros, L.; Ferreira, I.C.F.R. Catechin-based extract optimization obtained from Arbutus unedo L. fruits using maceration/microwave/ultrasound extraction techniques. Ind. Crops Prod. 2017, 95, 404–415. [Google Scholar] [CrossRef]
- Zhang, Q.-W.; Lin, L.-G.; Ye, W.-C. Techniques for extraction and isolation of natural products: A comprehensive review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef]
- Munoz, L.; Rosell, G.; Quero, C.; Guerrero, A. Biosynthetic pathways of the pheromone of the Egyptian armyworm Spodoptera littoralis. Physiol. Entomol. 2008, 33, 275–290. [Google Scholar] [CrossRef]
- Kim, J.H.; Jander, G. Myzus persicae (green peach aphid) feeding on Arabidopsis induces the formation of a deterrent indole glucosinolate. Plant J. 2007, 49, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Greenslade, A.F.C.; Ward, J.L.; Martin, J.L.; Corol, D.I.; Clark, S.J.; Smart, L.E.; Aradottir, G.I. Triticum monococcum lines with distinct metabolic phenotypes and phloem-based partial resistance to the bird cherry-oat aphid Rhopalosiphum padi. Ann. Appl. Biol. 2016, 168, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Castagnone-Sereno, P. Genetic variability and adaptive evolution in parthenogenetic root-knot nematodes. Heredity 2006, 96, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Zhao, Q.-S.; Zhang, H.-J.; Lin, Z.-W.; Sun, H.-D. New cleroindicins from Clerodendrum indicum. J. Nat. Prod. 1997, 60, 766–769. [Google Scholar] [CrossRef]
- Endo, K.; Hikino, H. Structures of rengyol, rengyoxide, and rengyolone, new cyclohexylethane derivatives from Forsythia suspensa fruits. Can. J. Chem. 1984, 62, 2011–2014. [Google Scholar] [CrossRef]
- Messana, I.; Sperandei, M.; Multari, G.; Galeffi, C.; Bettolo, G.B.M. A cyclohexadienone and a cyclohexenone from Halleria lucida. Phytochemistry 1984, 23, 2617–2619. [Google Scholar] [CrossRef]
- Breton, J.L.; Llera, L.D.; Navarro, E.; Trujillo, J. Photochemical synthesis of halleridone, hallerone, rengyol and derivatives. Tetrahedron 1987, 43, 4447–4451. [Google Scholar] [CrossRef]
- Endo, K.; Seya, K.; Hikino, H. Biogenesis-like transformation of salidroside to rengyol and its related cyclohexyletanoids of Forsythia suspensa. Tetrahedron 1989, 45, 3673–3682. [Google Scholar] [CrossRef]
- Carreño, M.C.; González-López, M.; Urbano, A. Oxidative de-aromatization of para-alkyl phenols into para-peroxyquinols and para-quinols mediated by oxone as a source of singlet oxygen. Angew. Chem. Int. Ed. 2006, 45, 2737–2741. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Krull, R.E. Iridoid glycosides of Cornus canadensis: A comparison with some other Cornus species. Biochem. Syst. Ecol. 1998, 26, 845–849. [Google Scholar] [CrossRef]
- Bianco, A.; Scalzo, R.L.; Scarpati, M.L. Isolation of cornoside from Olea europaea and its transformation into halleridone. Phytochemistry 1993, 32, 455–457. [Google Scholar] [CrossRef]
- Jensen, S.R. Chemical relationships of Polypremum procumbens, Tetrachondra hamiltonii and Peltanthera floribunda. Biochem. Syst. Ecol. 2000, 28, 45–51. [Google Scholar] [CrossRef]
- Kim, D.-H.; Han, K.-M.; Bang, M.-H.; Lee, Y.-H.; Chung, I.-S.; Kim, D.-K.; Kim, S.-H.; Kwon, B.-M.; Park, M.-H.; Baek, N.-I. Cyclohexylethanoids from the flower of Campsis grandiflora. Bull. Korean Chem. Soc. 2007, 28, 1851–1853. [Google Scholar] [CrossRef][Green Version]
- Jensen, S.R.; Kjaer, A.; Nielsen, B.J. Quinol glucoside isolated from Cornus species. Acta Chem. Scand. 1973, 27, 367–369. [Google Scholar] [CrossRef]
- Ishihara, K.; Kurihara, H.; Yamamoto, H. An extremely simple, convenient, and selective method for acetylating primary alcohols in the presence of secondary alcohols. J. Org. Chem. 1993, 58, 3791–3793. [Google Scholar] [CrossRef]
- Navarro, E.; Alonso, S.J.; Boada, J.; Trujillo, J.; Ayuso, M.J.; Gutierrez Navarro, A. Quinolic derivatives: Biological activity. Fitoterapia 1992, 63, 251–254. [Google Scholar]
- Navarro-Rocha, J.; Andrés, M.F.; Díaz, C.E.; Burillo, J.; González-Coloma, A. Composition and biocidal properties of essential oil from pre-domesticated Spanish Satureja montana. Ind. Crops Prod. 2020, 145, 111958. [Google Scholar] [CrossRef]
- Püntener, W. Manual for Field Trials in Plant Protection; Ciba-Geigy Limited: Basel, Switzerland, 1981; 205p. [Google Scholar]
- Lopes, M.B.P.; Mendonça, P.M.; Mallet, J.R.S.; Carvalho, M.G.; Queiroz, M.M.C. Bioactivity of the latex from Parahancornia amapa (Apocynaceae) on the development of Rhodnius nasutus (Hemiptera, Reduviidae, Triatominae) under laboratory conditions. Rev. Bras. Entomol. 2014, 58, 379–383. [Google Scholar] [CrossRef]
- Fraga, B.M.; Díaz, C.E.; Amador, L.J.; Reina, M.; Santana, O.; González-Coloma, A. Bioactive compounds from transformed root cultures and aerial parts of Bethencourtia hermosae. Phytochemistry 2014, 108, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Method | Fats | Phenyletanoids | Phenyletanoids (Glycosylated) | Cardenolides | Free Sugars |
|---|---|---|---|---|---|---|
| 1 | 70% EtOH (reflux) b | tr e | + | + + | + | + + |
| 2 | EtOH (reflux) | tr | + | + + | + | + + |
| 3 | MTBE (rt c) | + + | + | − | − | − |
| 4 | EtOAc (rt) | + + | + | − | − | − |
| 5 | EtOH (40 °C) | tr | + + | − | − | + |
| 6 | EtOH (40 °C) d | tr | + + | − | − | + |
| Compound | % SI a | EC50 b (µg/cm2) |
|---|---|---|
| 1 | 86.31 ± 5.74 * | 17.6 (13.2–23.4) |
| 3 | 75.77 ± 6.13 * | 25.9 (18.5–36.1) |
| 5 | 76.81 ± 5.51 * | 12.5 (7.0–22.0) |
| 6 | 53.78 ± 8.58 | |
| Thymol | 81.8 ± 7.7 * | 7.6 (4.1–8.7) |
| Compound | % Mortality a |
|---|---|
| 1 | 34.17 ± 10.81 |
| 3 | 2.47 ± 1.56 |
| 5 | 92.87 ± 1.26 |
| 6 | 20.34 ± 2.52 |
| Compound | Dose (µg/mg) | J2 Mortality (%) a | LD50 b |
|---|---|---|---|
| 5 | 1.0 | 92.87 ± 1.23 | 0.034 (0.017–0.091) |
| 0.5 | 82.13 ± 4.1 | ||
| 0.25 | 75.83 ± 1.39 | ||
| 0.12 | 46.82 ± 5.11 | ||
| 0.06 | 40.93 ± 2.59 | ||
| Thymol | 1.0 | 100 | 0.14 (0.131–0.143) |
| 0.50 | 100 | ||
| 0.25 | 98 ± 0.44 | ||
| 0.12 | 29.14 ± 2.92 | ||
| 0.06 | 15.40 ± 2.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quílez del Moral, J.F.; Pérez, Á.; Navarro, M.J.S.; Galisteo, A.; Gonzalez-Coloma, A.; Andrés, M.F.; Barrero, A.F. Selective Extraction of Bioactive Phenylethanoids from Digitalis obscura. Plants 2021, 10, 959. https://doi.org/10.3390/plants10050959
Quílez del Moral JF, Pérez Á, Navarro MJS, Galisteo A, Gonzalez-Coloma A, Andrés MF, Barrero AF. Selective Extraction of Bioactive Phenylethanoids from Digitalis obscura. Plants. 2021; 10(5):959. https://doi.org/10.3390/plants10050959
Chicago/Turabian StyleQuílez del Moral, José Francisco, Álvaro Pérez, María José Segura Navarro, Alberto Galisteo, Azucena Gonzalez-Coloma, María Fe Andrés, and Alejandro F. Barrero. 2021. "Selective Extraction of Bioactive Phenylethanoids from Digitalis obscura" Plants 10, no. 5: 959. https://doi.org/10.3390/plants10050959
APA StyleQuílez del Moral, J. F., Pérez, Á., Navarro, M. J. S., Galisteo, A., Gonzalez-Coloma, A., Andrés, M. F., & Barrero, A. F. (2021). Selective Extraction of Bioactive Phenylethanoids from Digitalis obscura. Plants, 10(5), 959. https://doi.org/10.3390/plants10050959