
Culturable Yeasts as Biofertilizers and Biopesticides for a Sustainable Agriculture: A Comprehensive Review


Abstract
1. Introduction
2. European and International Regulations on the Use of Biostimulants and Biopesticides
2.1. Biostimulant Regulations
2.2. Biopesticides Regulations
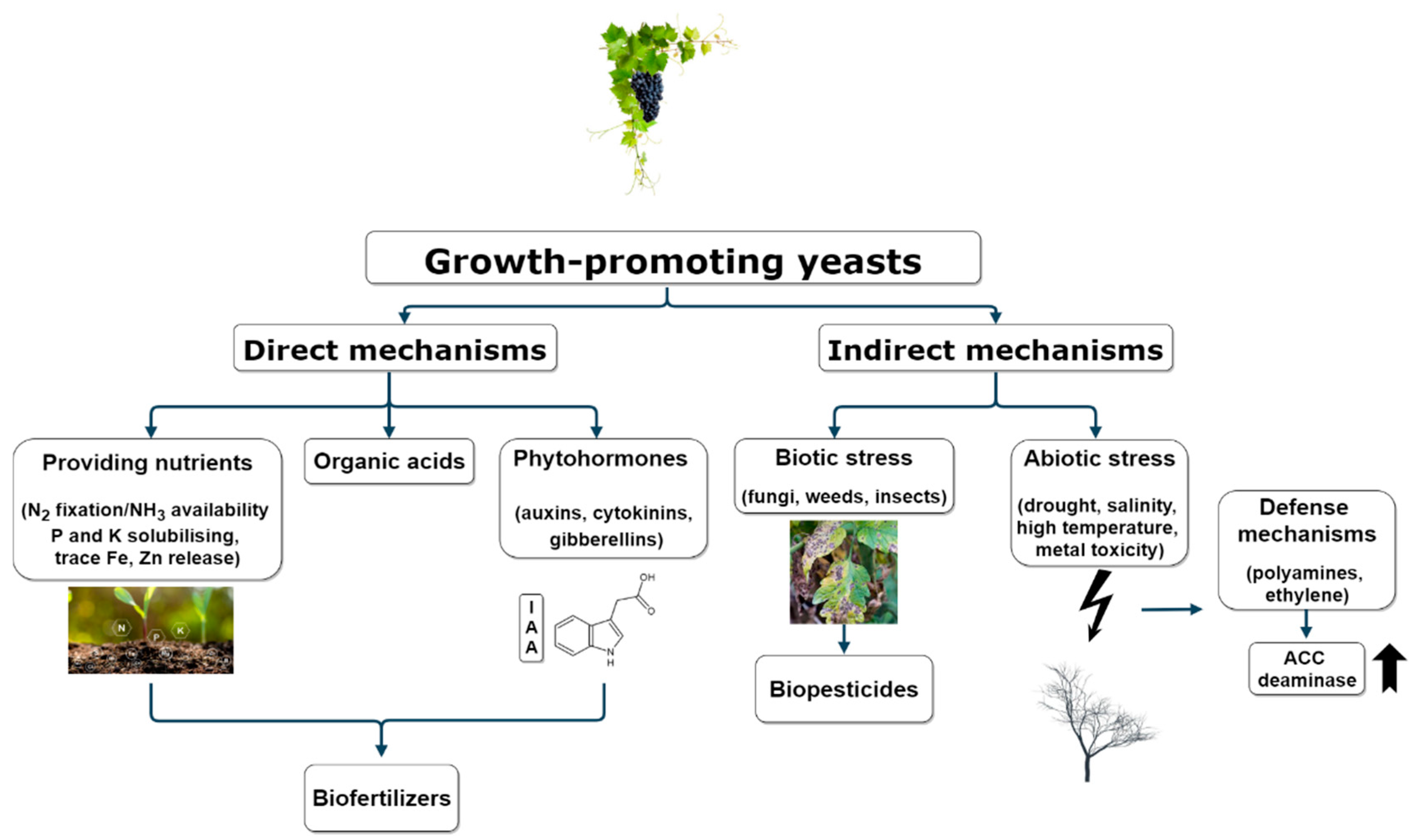
3. Yeasts as Biostimulant and Biofertilizer Agents
3.1. Direct Plant Growth Mechanisms
3.1.1. Nutrient Supply
3.1.2. Phytohormones
3.2. Indirect Plant Growth Mechanisms
4. Yeasts as Biocontrol against Fungal Pathogenic Microorganisms
4.1. Competition for Nutrients and Space
4.2. VOCs and Killer Toxins Production
4.3. Parasitism and iNduced Systemic Resistance
5. Yeasts as Bioinsecticide, Bioherbicide and Biodegrader
6. Application of Yeasts in Commercial Biofertilizers and Biopesticides
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, P.J.; Brown, P.H. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef]
- Gourley, C.J.P.; Dougherty, W.J.; Weaver, D.M.; Aarons, S.R.; Awty, I.M.; Gibson, D.M.; Hannah, M.C.; Smith, A.P.; Peverill, K.I. Farm-scale nitrogen, phosphorus, potassium and sulfur balances and use efficiencies on Australian dairy farms. Anim. Prod. Sci. 2012, 52, 929. [Google Scholar] [CrossRef]
- Ceccon, E. La revolución verde. Tragedia en dos actos. Ciencias Univ. Nac. Autónoma México 2008, 1, 21–29. [Google Scholar]
- Abdelhafez, A.A.; Abbas, H.H.; Abd-El-Aal, R.S.; Kandil, N.F.; Li, J.; Mahmoud, W. Environmental and Health Impacts of Successive Mineral Fertilization in Egypt. Clean Soil Air Water 2012, 40, 356–363. [Google Scholar] [CrossRef]
- Pimentel, D. Green revolution agriculture and chemical hazards. Sci. Total Environ. 1996, 188, S86–S98. [Google Scholar] [CrossRef]
- Stolte, J.; Tesfai, M.; Keizer, J. Soil Threats in Europe: Status, Methods, Drivers and Effects on Ecosystem Services; European Commission DG Joint Research Centre: Brussels, Belgium, 2016; ISBN 9789279540189. [Google Scholar]
- FAO. The future of food and agriculture—Alternative pathways to 2050|Global Perspectives Studies|Food and Agriculture Organization of the United Nations. In Food and Agriculture Organization; FAO: Rome, Italy, 2018; p. 224. ISBN 978-92-5-130158-6. [Google Scholar]
- Ju, X.; Zhang, F.; Bao, X.; Römheld, V.; Roelcke, M. Utilization and management of organic wastes in Chinese agriculture: Past, present and perspectives. Sci. China C. Life Sci. 2005, 48, 965–979. [Google Scholar] [CrossRef]
- Grantina-Ievina, L.; Rodze, I. Survival of Pathogenic and Antibiotic-Resistant Bacteria in Vermicompost, Sewage Sludge, and Other Types of Composts in Temperate Climate Conditions; Springer: Cham, Germany, 2020; pp. 107–124. [Google Scholar]
- Case, S.D.C.; Oelofse, M.; Hou, Y.; Oenema, O.; Jensen, L.S. Farmer perceptions and use of organic waste products as fertilisers—A survey study of potential benefits and barriers. Agric. Syst. 2017, 151, 84–95. [Google Scholar] [CrossRef]
- Singh, R.P.; Singh, P.; Araujo, A.S.F.; Hakimi Ibrahim, M.; Sulaiman, O. Management of urban solid waste: Vermicomposting a sustainable option. Resour. Conserv. Recycl. 2011, 55, 719–729. [Google Scholar] [CrossRef]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Alori, E.T.; Babalola, O.O. Microbial inoculants for improving crop quality and human health in Africa. Front. Microbiol. 2018, 9, 2213. [Google Scholar] [CrossRef]
- Pandey, V.; Chandra, K. Agriculturally important microorganisms as biofertilizers: Commercialization and regulatory requirements in Asia. In Agriculturally Important Microorganisms: Commercialization and Regulatory Requirements in Asia; Springer: Singapore, 2016; pp. 133–145. ISBN 9789811025761. [Google Scholar]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Fact. 2014, 13, 1–10. [Google Scholar] [CrossRef]
- Beneduzi, A.; Peres, D.; Vargas, L.K.; Bodanese-Zanettini, M.H.; Passaglia, L.M.P. Evaluation of genetic diversity and plant growth promoting activities of nitrogen-fixing bacilli isolated from rice fields in South Brazil. Appl. Soil Ecol. 2008, 39, 311–320. [Google Scholar] [CrossRef]
- Li, J.H.; Wang, E.T.; Chen, W.F.; Chen, W.X. Genetic diversity and potential for promotion of plant growth detected in nodule endophytic bacteria of soybean grown in Heilongjiang province of China. Soil Biol. Biochem. 2008, 40, 238–246. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Singh, H.; Reddy, S.M. Improvement of wheat and maize crops by inoculating Aspergillus spp. in alkaline soil fertilized with rock phosphate. Arch. Agron. Soil Sci. 2012, 58, 535–546. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 1–14. [Google Scholar] [CrossRef]
- Nehl, D.B.; Allen, S.J.; Brown, J.F. Deleterious rhizosphere bacteria: An integrating perspective. Appl. Soil Ecol. 1997, 5, 1–20. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 871, 1473. [Google Scholar] [CrossRef]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- E. Birch, A.N.; Begg, G.S.; Squire, G.R. How agro-ecological research helps to address food security issues under new IPM and pesticide reduction policies for global crop production systems. J. Exp. Bot. 2011, 62, 3251–3261. [Google Scholar] [CrossRef] [PubMed]
- von Rumker, R.; Kelso, G.; Horay, F.; Lawrence, K.A. A study of the Efficiency of the Use of Pesticides in Agriculture; Office of Pesticide Programs, Environmental Protection Agency, Ed.; EPA Office of Pesticide Programs, Strategic Studies Unit: Washington, DC, USA, 1975; Volume 1.
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.S.; Saxena, A.K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatal. Agric. Biotechnol. 2020, 23, 101487. [Google Scholar] [CrossRef]
- Sporleder, M.; Lacey, L.A. Biopesticides. In Insect Pests of Potato; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 463–497. ISBN 9780123868954. [Google Scholar]
- Olson, S. An analysis of the biopesticide market now and where it is going. Outlooks Pest. Manag. 2015, 26, 203–206. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. (Eds.) The Yeasts: A Taxonomic Study, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, ISBN 978-0-123-84708-9. [Google Scholar]
- Yurkov, A.M. Yeasts of the soil—Obscure but precious. Yeast 2018, 35, 369–378. [Google Scholar] [CrossRef]
- Dai, Y.J.; Ji, W.W.; Chen, T.; Zhang, W.J.; Liu, Z.H.; Ge, F.; Sheng, Y. Metabolism of the neonicotinoid insecticides acetamiprid and thiacloprid by the yeast Rhodotorula mucilaginosa strain IM-2. J. Agric. Food Chem. 2010, 58, 2419–2425. [Google Scholar] [CrossRef] [PubMed]
- Amprayn, K.O.; Rose, M.T.; Kecskés, M.; Pereg, L.; Nguyen, H.T.; Kennedy, I.R. Plant growth promoting characteristics of soil yeast (Candida tropicalis HY) and its effectiveness for promoting rice growth. Appl. Soil Ecol. 2012, 61, 295–299. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Kannan, V.R. Evaluation of the production of exopolysaccharide by plant growth promoting yeast Rhodotorula sp. strain CAH2 under abiotic stress conditions. Int. J. Biol. Macromol. 2019, 121, 55–62. [Google Scholar] [CrossRef]
- Kumla, J.; Nundaeng, S.; Suwannarach, N.; Lumyong, S. Evaluation of multifarious plant growth promoting trials of yeast isolated from the soil of assam tea (Camellia sinensis var. assamica) plantations in Northern Thailand. Microorganisms 2020, 8, 1168. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Mangieri, N.; Maghradze, D.; Foschino, R.; Valdetara, F.; Cantoral, J.M.; Vigentini, I. Wild Grape-Associated Yeasts as Promising Biocontrol Agents against Vitis vinifera Fungal Pathogens. Front. Microbiol. 2017, 8, 2025. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Murphy, K.A.; Tabuloc, C.A.; Cervantes, K.R.; Chiu, J.C. Ingestion of genetically modified yeast symbiont reduces fitness of an insect pest via RNA interference. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef]
- Epp, J.B.; Alexander, A.L.; Balko, T.W.; Buysse, A.M.; Brewster, W.K.; Bryan, K.; Daeuble, J.F.; Fields, S.C.; Gast, R.E.; Green, R.A.; et al. The discovery of ArylexTM active and RinskorTM active: Two novel auxin herbicides. Bioorganic Med. Chem. 2016, 24, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Gava, C.A.T.; de Castro, A.P.C.; Pereira, C.A.; Fernandes-Júnior, P.I. Isolation of fruit colonizer yeasts and screening against mango decay caused by multiple pathogens. Biol. Control. 2018, 117, 137–146. [Google Scholar] [CrossRef]
- Fu, S.F.; Sun, P.F.; Lu, H.Y.; Wei, J.Y.; Xiao, H.S.; Fang, W.T.; Cheng, B.Y.; Chou, J.Y. Plant growth-promoting traits of yeasts isolated from the phyllosphere and rhizosphere of Drosera spatulata Lab. Fungal Biol. 2016, 120, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Damalas, C.A.; Koutroubas, S.D. Current Status and Recent Developments in Biopesticide Use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef]
- Caradonia, F.; Battaglia, V.; Righi, L.; Pascali, G.; La Torre, A. Plant Biostimulant Regulatory Framework: Prospects in Europe and Current Situation at International Level. J. Plant Growth Regul. 2019, 38, 438–448. [Google Scholar] [CrossRef]
- Thakur, N.; Kaur, S.; Tomar, P.; Thakur, S.; Yadav, A.N. Microbial biopesticides: Current status and advancement for sustainable agriculture and environment. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2020; pp. 243–282. [Google Scholar]
- Mascarin, G.M.; Lopes, R.B.; Delalibera, Í.; Fernandes, É.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef]
- Gasser, B.; Prielhofer, R.; Marx, H.; Maurer, M.; Nocon, J.; Steiger, M.; Puxbaum, V.; Sauer, M.; Mattanovich, D. Pichia pastoris: Protein production host and model organism for biomedical research. Future Microbiol. 2013, 8, 191–208. [Google Scholar] [CrossRef]
- Kumar, R.; Angov, E.; Kumar, N. Potent malaria transmission-blocking antibody responses elicited by Plasmodium falciparum Pfs25 expressed in Escherichia coli after successful protein refolding. Infect. Immun. 2014, 82, 1453–1459. [Google Scholar] [CrossRef]
- Kwak, S.; Jo, J.H.; Yun, E.J.; Jin, Y.S.; Seo, J.H. Production of biofuels and chemicals from xylose using native and engineered yeast strains. Biotechnol. Adv. 2019, 37, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Buijs, N.A.; Zhu, Z.; Qin, J.; Siewers, V.; Nielsen, J. Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat. Commun. 2016, 7, 11709. [Google Scholar] [CrossRef]
- Mukherjee, A.; Verma, J.P.; Gaurav, A.K.; Chouhan, G.K.; Patel, J.S.; Hesham, A.E.L. Yeast a potential bio-agent: Future for plant growth and postharvest disease management for sustainable agriculture. Appl. Microbiol. Biotechnol. 2020, 104, 1497–1510. [Google Scholar] [CrossRef]
- Botha, A. The importance and ecology of yeasts in soil. Soil Biol. Biochem. 2011, 43, 1–8. [Google Scholar] [CrossRef]
- Rosa-Magri, M.M.; Avansini, S.H.; Lopes-Assad, M.L.; Tauk-Tornisielo, S.M.; Ceccato-Antonini, S.R. Release of potassium from rock powder by the yeast Torulaspora globosa. Brazilian Arch. Biol. Technol. 2012, 55, 577–582. [Google Scholar] [CrossRef]
- El-Maraghy, S.S.; Tohamy, T.A.; Hussein, K.A. Expression of SidD gene and physiological characterization of the rhizosphere plant growth-promoting yeasts. Heliyon 2020, 6, e04384. [Google Scholar] [CrossRef] [PubMed]
- Agamy, R.; Hashem, M.; Alamri, S. Effect of soil amendment with yeasts as bio-fertilizers on the growth and productivity of sugar beet. Afr. J. Agric. Res. 2013, 7, 6613–6623. [Google Scholar] [CrossRef]
- Nakayan, P.; Hameed, A.; Singh, S.; Young, L.S.; Hung, M.H.; Young, C.C. Phosphate-solubilizing soil yeast Meyerozyma guilliermondii CC1 improves maize (Zea mays L.) productivity and minimizes requisite chemical fertilization. Plant Soil 2013, 373, 301–315. [Google Scholar] [CrossRef]
- Khan, Z.; Guelich, G.; Phan, H.; Redman, R.; Doty, S.; Arencibia, A.D.; Chodak, M.; Perez-Artes, E.; Tsushima, S. Bacterial and Yeast Endophytes from Poplar and Willow Promote Growth in Crop Plants and Grasses. Int. Sch. Res. Not. 2012, 2012, 11. [Google Scholar] [CrossRef]
- Knoth, J.L.; Kim, S.-H.; Ettl, G.J.; Doty, S.L. Effects of cross host species inoculation of nitrogen-fixing endophytes on growth and leaf physiology of maize. GCB Bioenergy 2013, 5, 408–418. [Google Scholar] [CrossRef]
- Nutaratat, P.; Srisuk, N.; Arunrattiyakorn, P.; Limtong, S. Plant growth-promoting traits of epiphytic and endophytic yeasts isolated from rice and sugar cane leaves in Thailand. Fungal Biol. 2014, 118, 683–694. [Google Scholar] [CrossRef]
- Sun, P.-F.; Fang, W.-T.; Shin, L.-Y.; Wei, J.-Y.; Fu, S.-F.; Chou, J.-Y. Indole-3-Acetic Acid-Producing Yeasts in the Phyllosphere of the Carnivorous Plant Drosera indica L. PLoS ONE 2014, 9, e114196. [Google Scholar] [CrossRef]
- Ibraheim, S.K.A. Effect of Foliar Spray with Some Biostimulants on Growth, Yield and Seeds Quality of Pea Plants Grown in Sandy Soil. J. Appl. Sci. Res. 2014, 10, 400–407. [Google Scholar]
- Złotek, U.; Świeca, M. Elicitation effect of Saccharomyces cerevisiae yeast extract on main health-promoting compounds and antioxidant and anti-inflammatory potential of butter lettuce (Lactuca sativa L.). J. Sci. Food Agric. 2016, 96, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Naik, K.; Mishra, S.; Srichandan, H.; Singh, P.K.; Sarangi, P.K. Plant growth promoting microbes: Potential link to sustainable agriculture and environment. Biocatal. Agric. Biotechnol. 2019, 21, 101326. [Google Scholar] [CrossRef]
- Nutaratat, P.; Srisuk, N.; Arunrattiyakorn, P.; Limtong, S. Indole-3-acetic acid biosynthetic pathways in the basidiomycetous yeast Rhodosporidium paludigenum. Arch. Microbiol. 2016, 198, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Montaño, F.; Alías-Villegas, C.; Bellogín, R.A.; Del Cerro, P.; Espuny, M.R.; Jiménez-Guerrero, I.; López-Baena, F.J.; Ollero, F.J.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef]
- Leghari, S.J.; Wahocho, N.A.; Laghari, G.M.; HafeezLaghari, A.; MustafaBhabhan, G.; HussainTalpur, K.; Bhutto, T.A.; Wahocho, S.A.; Lashari, A.A. Role of nitrogen for plant growth and development: A review. Adv. Environ. Biol. 2016, 10, 209–219. [Google Scholar]
- Mukherjee, S.; Sen, S.K. Exploration of novel rhizospheric yeast isolate as fertilizing soil inoculant for improvement of maize cultivation. J. Sci. Food Agric. 2015, 95, 1491–1499. [Google Scholar] [CrossRef]
- Fernandez-San Millan, A.; Farran, I.; Larraya, L.; Ancin, M.; Arregui, L.M.; Veramendi, J. Plant growth-promoting traits of yeasts isolated from Spanish vineyards: Benefits for seedling development. Microbiol. Res. 2020, 237, 126480. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.X.; Rossi, M.J.; Soares, C.R.F.S.; McConkey, B.J.; Glick, B.R. New Insights into 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminase Phylogeny, Evolution and Ecological Significance. PLoS ONE 2014, 9, e99168. [Google Scholar] [CrossRef]
- Deng, Z.; Wang, W.; Tan, H.; Cao, L. Characterization of heavy metal-resistant endophytic yeast Cryptococcus sp. CBSB78 from rapes (Brassica chinensis) and its potential in promoting the growth of Brassica spp. in metal-contaminated soils. Water Air Soil Pollut. 2012, 223, 5321–5329. [Google Scholar] [CrossRef]
- Mothapo, N.; Chen, H.; Cubeta, M.A.; Grossman, J.M.; Fuller, F.; Shi, W. Phylogenetic, taxonomic and functional diversity of fungal denitrifiers and associated N2O production efficacy. Soil Biol. Biochem. 2015, 83, 160–175. [Google Scholar] [CrossRef]
- Vero, S.; Garmendia, G.; Martínez-Silveira, A.; Cavello, I.; Wisniewski, M. Yeast Activities Involved in Carbon and Nitrogen Cycles in Antarctica. In The Ecological Role of Micro-Organisms in the Antarctic Environment; Springer: Cham, Germany, 2019; pp. 45–64. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Ahmad, E. Mechanism of phosphate solubilization and physiological functions of phosphate-solubilizing microorganisms. In Phosphate Solubilizing Microorganisms: Principles and Application of Microphos Technology; Springer International Publishing: Berlin/Heidelberg, Germany, 2014; pp. 31–62. ISBN 9783319082165. [Google Scholar]
- Ramaekers, L.; Remans, R.; Rao, I.M.; Blair, M.W.; Vanderleyden, J. Strategies for improving phosphorus acquisition efficiency of crop plants. F. Crop. Res. 2010, 117, 169–176. [Google Scholar] [CrossRef]
- Sharma, S.; Kumar, V.; Tripathi, R.B. Isolation of Phosphate Solubilizing Microorganism (PSMs) From Soil. J. Microbiol. Biotechnol. Res. Sch. Res. Libr. J. Microbiol. Biotech. Res. 2011, 1, 90–95. [Google Scholar]
- Mundra, S.; Arora, R.; Stobdan, T. Solubilization of insoluble inorganic phosphates by a novel temperature-, pH-, and salt-tolerant yeast, Rhodotorula sp. PS4, isolated from seabuckthorn rhizosphere, growing in cold desert of Ladakh, India. World J. Microbiol. Biotechnol. 2011, 27, 2387–2396. [Google Scholar] [CrossRef]
- El-Latif, A.; Mohamed, H.M. Molecular Genetic Identification of Yeast Strains Isolated from Egyptian Soils for Solubilization of Inorganic Phosphates and Growth Promotion of Corn Plants. J. Microbiol. Biotechnol. 2011, 21, 55–61. [Google Scholar] [CrossRef]
- Velázquez, E.; Silva, L.R.; Ramírez-Bahena, M.H.; Peix, A. Diversity of potassium-solubilizing microorganisms and their interactions with plants. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 99–110. ISBN 9788132227762. [Google Scholar]
- Mohamed, H.M.; El-Homosy, R.F.; Abd-Ellatef, A.-E.H.; Salh, F.M.; Hussein, M.Y. Identification of Yeast Strains Isolated from Agricultural Soils for Releasing Potassium-bearing Minerals. Geomicrobiol. J. 2017, 34, 261–266. [Google Scholar] [CrossRef]
- Rojas-Tapias, D.; Moreno-Galván, A.; Pardo-Díaz, S.; Obando, M.; Rivera, D.; Bonilla, R. Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize (Zea mays). Appl. Soil Ecol. 2012, 61, 264–272. [Google Scholar] [CrossRef]
- Aiysha, D.; Latif, Z. Insights of organic fertilizer micro flora of bovine manure and their useful potentials in sustainable agriculture. PLoS ONE 2019, 14, e0226155. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Stöven, K.; Haneklaus, S.; Singh, B.R.; Schnug, E. Elemental Sulfur Oxidation by Thiobacillus spp. and Aerobic Heterotrophic Sulfur-Oxidizing Bacteria. Pedosphere 2010, 20, 71–79. [Google Scholar] [CrossRef]
- Falih, A.M.; Wainwright, M. Nitrification, S-oxidation and P-solubilization by the soil yeast Williopsis californica and by Saccharomyces cerevisiae. Mycol. Res. 1995, 99, 200–204. [Google Scholar] [CrossRef]
- Hafeez, B. Role of Zinc in Plant Nutrition- A Review. Am. J. Exp. Agric. 2013, 3, 374–391. [Google Scholar] [CrossRef]
- Ansari, R.A.; Mahmood, I.; Rizvi, R.; Sumbul, A. Safiuddin Siderophores: Augmentation of soil health and crop productivity. In Probiotics in Agroecosystem; Springer: Singapore, 2017; pp. 291–312. ISBN 9789811040597. [Google Scholar]
- Rajkumar, M.; Ae, N.; Prasad, M.N.V.; Freitas, H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef]
- Kasahara, H. Current aspects of auxin biosynthesis in plants. Biosci. Biotechnol. Biochem. 2016, 80, 34–42. [Google Scholar] [CrossRef]
- Bunsangiam, S.; Sakpuntoon, V.; Srisuk, N.; Ohashi, T.; Fujiyama, K.; Limtong, S. Biosynthetic Pathway of Indole-3-Acetic Acid in Basidiomycetous Yeast Rhodosporidiobolus fluvialis. Mycobiology 2019, 47, 292–300. [Google Scholar] [CrossRef]
- Limtong, S.; Koowadjanakul, N. Yeasts from phylloplane and their capability to produce indole-3-acetic acid. World J. Microbiol. Biotechnol. 2012, 28, 3323–3335. [Google Scholar] [CrossRef] [PubMed]
- Prabina, B.J.; Kumutha, K.; Anandham, R.; Durga, P. Isolation and Characterization of Multifunctional Yeast as Plant Probiotics for Better Crop Nutrition in Pulses. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2711–2718. [Google Scholar] [CrossRef]
- Rao, R.P.; Hunter, A.; Kashpur, O.; Normanly, J. Aberrant Synthesis of Indole-3-Acetic Acid in Saccharomyces cerevisiae Triggers Morphogenic Transition, a Virulence Trait of Pathogenic Fungi. Genetics 2010, 185, 211–220. [Google Scholar] [CrossRef]
- Giri, R.; Sharma, R.K. Fungal pretreatment of lignocellulosic biomass for the production of plant hormone by Pichia fermentans under submerged conditions. Bioresour. Bioprocess. 2020, 7, 1–11. [Google Scholar] [CrossRef]
- Streletskii, R.A.; Kachalkin, A.V.; Glushakova, A.M.; Yurkov, A.M.; Demin, V.V. Yeasts producing zeatin. PeerJ 2019, 2019, e6474. [Google Scholar] [CrossRef]
- Gupta, R.; Chakrabarty, S.K. Gibberellic acid in plant. Plant Signal. Behav. 2013, 8, e25504. [Google Scholar] [CrossRef] [PubMed]
- Pandi, R.; Velu, G.; Devi, P.; Dananjeyan, B. Isolation and screening of soil yeasts for plant growth promoting traits. Madras Agric. J. 2019. [Google Scholar] [CrossRef]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef]
- Pereira, A. Plant Abiotic Stress Challenges from the Changing Environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.S.; Ali, M.; Ahmad, M.; Siddique, K.H.M. Polyamines: Natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol. Adv. 2011, 29, 300–311. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, S. Ethylene Biosynthesis and Regulation in Plants. In Ethylene in Plants; Springer: Dordrecht, The Netherlands, 2015; pp. 1–25. [Google Scholar]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: A weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [CrossRef]
- Chanchaichaovivat, A.; Ruenwongsa, P.; Panijpan, B. Screening and identification of yeast strains from fruits and vegetables: Potential for biological control of postharvest chilli anthracnose (Colletotrichum capsici). Biol. Control. 2007, 42, 326–335. [Google Scholar] [CrossRef]
- Grebenisan, I.; Cornea, P.; Mateescu, R.; Cimpeanu, C.; Olteanu, V.; Campenu, G.; Stefan, L.A.; Oancea, F.; Lupu, C. Metschnikowia pulcherrima, a new yeast with potential for biocontrol of postharvest fruit rots. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science, Seoul, Korea, 13 August 2006; Volume 767, pp. 355–360. [Google Scholar]
- Muccilli, S.; Restuccia, C. Bioprotective Role of Yeasts. Microorganisms 2015, 3, 588. [Google Scholar] [CrossRef]
- Klein, M.N.; Kupper, K.C. Biofilm production by Aureobasidium pullulans improves biocontrol against sour rot in citrus. Food Microbiol. 2018, 69, 1–10. [Google Scholar] [CrossRef]
- Bencheqroun, S.K.; Bajji, M.; Massart, S.; Labhilili, M.; El Jaafari, S.; Jijakli, M.H. In vitro and in situ study of postharvest apple blue mold biocontrol by Aureobasidium pullulans: Evidence for the involvement of competition for nutrients. Postharvest Biol. Technol. 2007, 46, 128–135. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; D’Aquino, S.; Pagnotta, E.; Mari, M. Biocontrol of Monilinia laxa by Aureobasidium pullulans strains: Insights on competition for nutrients and space. Int. J. Food Microbiol. 2017, 248, 32–38. [Google Scholar] [CrossRef]
- Gore-Lloyd, D.; Sumann, I.; Brachmann, A.O.; Schneeberger, K.; Ortiz-Merino, R.A.; Moreno-Beltrán, M.; Schläfli, M.; Kirner, P.; Santos Kron, A.; Rueda-Mejia, M.P.; et al. Snf2 controls pulcherriminic acid biosynthesis and antifungal activity of the biocontrol yeast Metschnikowia Pulcherrima. Mol. Microbiol. 2019, 112, 317–332. [Google Scholar] [CrossRef]
- Ferramola, M.I.S.; Benuzzi, D.; Calvente, V.; Calvo, J.; Sansone, G.; Cerutti, S.; Raba, J. The use of siderophores for improving the control of postharvest diseases in stored fruits and vegetables. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex Research Center: Badajoz, Spain, 2013; pp. 1385–1394. [Google Scholar]
- Mari, M.; Bautista-Baños, S.; Sivakumar, D. Decay control in the postharvest system: Role of microbial and plant volatile organic compounds. Postharvest Biol. Technol. 2016, 122, 70–81. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Hernández, A.; Galvan, A.I.; Córdoba, M.G.; Casquete, R.; Serradilla, M.J.; Martín, A. Selection and application of antifungal VOCs-producing yeasts as biocontrol agents of grey mould in fruits. Food Microbiol. 2020, 92, 103556. [Google Scholar] [CrossRef]
- Di Francesco, A.; Zajc, J.; Gunde-Cimerman, N.; Aprea, E.; Gasperi, F.; Placì, N.; Caruso, F.; Baraldi, E. Bioactivity of volatile organic compounds by Aureobasidium species against gray mold of tomato and table grape. World J. Microbiol. Biotechnol. 2020, 36, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Budroni, M.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Alonso, A.; Marquina, D.; Santos, A. The biology of pichia membranifaciens killer toxins. Toxins 2017, 9, 112. [Google Scholar] [CrossRef] [PubMed]
- Izgu, D.A.; Kepekci, R.A.; Izgu, F. Inhibition of Penicillium digitatum and Penicillium italicum in vitro and in planta with Panomycocin, a novel exo-β-1,3-glucanase isolated from Pichia anomala NCYC 434. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2011, 99, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Tzelepis, G.; Karlsson, M. Killer toxin-like chitinases in filamentous fungi: Structure, regulation and potential roles in fungal biology. Fungal Biol. Rev. 2019, 33, 123–132. [Google Scholar] [CrossRef]
- Magallon-Andalon, C.G.; Luna-Solano, G.; Ragazzo-Sanchez, J.; Calderon-Santoyo, M. Parasitism and substrate competitions effect of antagonistic yeasts for biocontrol of Colletotrichum gloeosporioides in papaya (Carica papaya L.) var Maradol. Mex. J. Sci. Res. 2012, 1, 2–9. [Google Scholar]
- Lima, J.R.; Gondim, D.M.F.; Oliveira, J.T.A.; Oliveira, F.S.A.; Gonçalves, L.R.B.; Viana, F.M.P. Use of killer yeast in the management of postharvest papaya anthracnose. Postharvest Biol. Technol. 2013, 83, 58–64. [Google Scholar] [CrossRef]
- Castoria, R.; Wright, S.A.I. Host Responses to Biological Control Agents. In Post-Harvest Pathology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 2, pp. 171–181. [Google Scholar]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.; Gutiérrez Martínez, P.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Lu, L.; Lu, H.; Wu, C.; Fang, W.; Yu, C.; Ye, C.; Shi, Y.; Yu, T.; Zheng, X. Rhodosporidium paludigenum induces resistance and defense-related responses against Penicillium digitatum in citrus fruit. Postharvest Biol. Technol. 2013, 85, 196–202. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, L.; Li, Z.; Li, C.; Li, B.; Gu, X.; Zhang, X.; Zhang, H. Screening and identification of an antagonistic yeast controlling postharvest blue mold decay of pears and the possible mechanisms involved. Biol. Control 2019, 133, 26–33. [Google Scholar] [CrossRef]
- Singh, B.; Singh, K. Microbial degradation of herbicides. Crit. Rev. Microbiol. 2016, 42, 245–261. [Google Scholar] [CrossRef]
- Bempelou, E.D.; Vontas, J.G.; Liapis, K.S.; Ziogas, V.N. Biodegradation of chlorpyrifos and 3,5,6-trichloro-2-pyridinol by the epiphytic yeasts Rhodotorula glutinis and Rhodotorula rubra. Ecotoxicology 2018, 27, 1368–1378. [Google Scholar] [CrossRef]
- Zhang, J.; Khan, S.A.; Heckel, D.G.; Bock, R. Next-Generation Insect-Resistant Plants: RNAi-Mediated Crop Protection. Trends Biotechnol. 2017, 35, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.L.; Witzgall, P. Combining Mutualistic Yeast and Pathogenic Virus—A Novel Method for Codling Moth Control. J. Chem. Ecol. 2013, 39, 1019–1026. [Google Scholar] [CrossRef]
- Bueno, E.; Martin, K.R.; Raguso, R.A.; Mcmullen, J.G.; Hesler, S.P.; Loeb, G.M.; Douglas, A.E. Response of Wild Spotted Wing Drosophila (Drosophila suzukii) to Microbial Volatiles. J. Chem. Ecol. 2020, 46, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Mori, B.A.; Whitener, A.B.; Leinweber, Y.; Revadi, S.; Beers, E.H.; Witzgall, P.; Becher, P.G. Enhanced yeast feeding following mating facilitates control of the invasive fruit pest Drosophila suzukii. J. Appl. Ecol. 2017, 54, 170–177. [Google Scholar] [CrossRef]
- Bianchi, F.; Spitaler, U.; Castellan, I.; Cossu, C.S.; Brigadoi, T.; Duménil, C.; Angeli, S.; Robatscher, P.; Vogel, R.F.; Schmidt, S.; et al. Persistence of a yeast-based (Hanseniaspora uvarum) attract-and-kill formulation against drosophila Suzukii on grape leaves. Insects 2020, 11, 810. [Google Scholar] [CrossRef] [PubMed]
- Holkenbrink, C.; Ding, B.J.; Wang, H.L.; Dam, M.I.; Petkevicius, K.; Kildegaard, K.R.; Wenning, L.; Sinkwitz, C.; Lorántfy, B.; Koutsoumpeli, E.; et al. Production of moth sex pheromones for pest control by yeast fermentation. Metab. Eng. 2020, 62, 312–321. [Google Scholar] [CrossRef]
- Myers, J.P.; Antoniou, M.N.; Blumberg, B.; Carroll, L.; Colborn, T.; Everett, L.G.; Hansen, M.; Landrigan, P.J.; Lanphear, B.P.; Mesnage, R.; et al. Concerns over use of glyphosate-based herbicides and risks associated with exposures: A consensus statement. Environ. Health A Glob. Access Sci. Source 2016, 15, 19. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.P.K.; Sethi, N.; Mohan, A.; Datta, S.; Girdhar, M. Glyphosate toxicity for animals. Environ. Chem. Lett. 2018, 16, 401–426. [Google Scholar] [CrossRef]
- Stosiek, N.; Terebieniec, A.; Ząbek, A.; Młynarz, P.; Cieśliński, H.; Klimek-Ochab, M. N-phosphonomethylglycine utilization by the psychrotolerant yeast Solicoccozyma terricola M 3.1.4. Bioorg. Chem. 2019, 93, 102866. [Google Scholar] [CrossRef]
- Han, Y.; Tang, Z.; Bao, H.; Wu, D.; Deng, X.; Guo, G.; Ye, B.C.; Dai, B. Degradation of pendimethalin by the yeast YC2 and determination of its two main metabolites. RSC Adv. 2019, 9, 491–497. [Google Scholar] [CrossRef]
- Abigail, E.A.; Abdul Salam, J.; Das, N. Atrazine degradation in liquid culture and soil by a novel yeast pichia kudriavzevii strain Atz-EN-01 and its potential application for bioremediation. J. Appl. Pharm. Sci. 2013, 3, 35–43. [Google Scholar] [CrossRef]
- Quareshy, M.; Prusinska, J.; Li, J.; Napier, R. A cheminformatics review of auxins as herbicides. J. Exp. Bot. 2018, 69, 265–275. [Google Scholar] [CrossRef]
- Vargas, M.; Garrido, F.; Zapata, N.; Tapia, M. Isolation and Selection of Epiphytic Yeat for Biocontrol of Botrytis cinerea Pers. on Table Grapes. Chil. J. Agric. Res. 2012, 72, 332–337. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Nally, M.C.; Pesce, V.M.; Maturano, Y.P.; Muñoz, C.J.; Combina, M.; Toro, M.E.; de Figueroa, L.I.C.; Vazquez, F. Biocontrol of Botrytis cinerea in table grapes by non-pathogenic indigenous Saccharomyces cerevisiae yeasts isolated from viticultural environments in Argentina. Postharvest Biol. Technol. 2012, 64, 40–48. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Xue, C.; Yu, Z.; Yang, R.; Cai, Z.; Si, L. Biocontrol of gray mold in grapes with the yeast Hanseniaspora uvarum alone and in combination with salicylic acid or sodium bicarbonate. Postharvest Biol. Technol. 2015, 100, 160–167. [Google Scholar] [CrossRef]
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P. Botrytis cinerea and table grapes: A review of the main physical, chemical, and bio-based control treatments in post-harvest. Foods 2020, 9, 1138. [Google Scholar] [CrossRef]
- Kasfi, K.; Taheri, P.; Jafarpour, B.; Tarighi, S. Identification of epiphytic yeasts and bacteria with potential for biocontrol of grey mold disease on table grapes caused by Botrytis cinerea. Span. J. Agric. Res. 2018, 16. [Google Scholar] [CrossRef]
- Lemos, W.J.; Bovo, B.; Nadai, C.; Crosato, G.; Carlot, M.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol ability and action mechanism of Starmerella bacillaris (synonym Candida zemplinina) isolated from wine musts against gray mold disease agent Botrytis cinerea on grape and their effects on alcoholic fermentation. Front. Microbiol. 2016, 7, 1249. [Google Scholar] [CrossRef]
- Mewa-Ngongang, M.; Du Plessis, H.W.; Ntwampe, S.K.O.; Chidi, B.S.; Hutchinson, U.F.; Mekuto, L.; Jolly, N.P. The use of Candida pyralidae and Pichia kluyveri to control spoilage microorganisms of raw fruits used for beverage production. Foods 2019, 8, 454. [Google Scholar] [CrossRef]
- Mew-Ngongang, M.; Du Plessis, H.; Ntwampe, S.K.O.; Chidi, D.B.S.; Hutchinson, U.; Mekuto, L.; Jolly, N. Fungistatic and Fungicidal Behaviour of Candida pyralidae Y1117, Pichia kluyveri Y1125 and Pichia kluyveri Y1164 on the Biocontrol of Botrytis cinerea. In Proceedings of the 10th International Conference on Advances in Science, Engineering, Technology and Healthcare (ASETH-18), Cape Town, South Africa, 19–20 November 2018; pp. 19–20. [Google Scholar]
- Kawalekar, J.S. Role of biofertilizers and biopesticides for sustainable agriculture. Ed. J. Bio 2013, 2, 73–78. [Google Scholar]
- Nguyen, T.H.; Phan, T.C.; Choudhury, A.T.M.A.; Rose, M.T.; Deaker, R.J.; Kennedy, I.R. Biogro: A plant growth-promoting biofertilizer validated by 15 years’ research from laboratory selection to rice farmer’s fields of the mekong delta. In Agro-Environmental Sustainability; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; Volume 1, pp. 237–254. ISBN 9783319497242. [Google Scholar]
- Rose, M.T.; Phuong, T.L.; Nhan, D.K.; Cong, P.T.; Hien, N.T.; Kennedy, I.R. Up to 52% N fertilizer replaced by biofertilizer in lowland rice via farmer participatory research. Agron. Sustain. Dev. 2014, 34, 857–868. [Google Scholar] [CrossRef]
- Pang, S.F.; Lui, A.; Goold, G.; Chu, A.; Wong, W.; Li, S.; Chan, E.; Kwok, I.; Cheung, L. NutriSmart®: A fertiliser capable of re-establishing the sustainability of ecosystems and enhancing the productivity of farmland. In Proceedings of the 11th Australian Agronomy Conference, Geelong, VIC, Australia, 2–6 February 2003. [Google Scholar]
- Mondino, P.; Casanova, L.; Calero, G.; Bentancur, O.; Alaniz, S. Zimevit: Un biofungicida que combina la acción de una bacteria y una levadura para el control del moho gris de la vid ocasionado por Botrytis cinerea. Rev. Bras. Agroecol. 2012, 7, 127–134. [Google Scholar]
- Zhimo, V.Y.; Kumar, A.; Biasi, A.; Salim, S.; Feygenberg, O.; Toamy, M.A.; Abdelfattaah, A.; Medina, S.; Freilich, S.; Wisniewski, M.; et al. Compositional shifts in the strawberry fruit microbiome in response to near-harvest application of Metschnikowia fructicola, a yeast biocontrol agent. Postharvest Biol. Technol. 2021, 175, 111469. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Droby, S. Metschnikowia fructicola, a new ascosporic yeast with potential for biocontrol of postharvest fruit rots. Syst. Appl. Microbiol. 2001, 24, 395–399. [Google Scholar] [CrossRef]
- Karabulut, O.A.; Tezcan, H.; Daus, A.; Cohen, L.; Wiess, B.; Droby, S. Control of preharvest and postharvest fruit rot in strawberry by Metschnikowia fructicola. Biocontrol Sci. Technol. 2004, 14, 513–521. [Google Scholar] [CrossRef]
- Prodorutti, D.; Ferrari, A.; Pellegrini, A.; Pertot, I. Efficacy of Metschnikowia fructicola (Shemer®) against post-harvest soft fruit (berries) rots in northern Italy (Trentino). IOBC/WPRS Bull. 2008, 39, 107–113. [Google Scholar]
- Lahlali, R.; Jijakli, M.H. Enhancement of the biocontrol agent Candida oleophila (strain O) survival and control efficiency under extreme conditions of water activity and relative humidity. Biol. Control. 2009, 51, 403–408. [Google Scholar] [CrossRef]
- Sebastien, M.; Jijakli, M.H. Pichia anomala and Candida oleophila in Biocontrol of Postharvest Diseases of Fruits: 20 Years of Fundamental and Practical Research. In Post-Harvest Pathology; Springer: Berlin/Heidelberg, Germany, 2014; pp. 111–122. ISBN 19077017. [Google Scholar]
- Ballet, N.; Souche, J.L.; Vandekerckove, P. Efficacy of Candida oleophila, strain O, In preventing postharvest diseases of fruits. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science, Florence, Italy, 7 June 2016; Volume 1144, pp. 105–111. [Google Scholar]
- Weiss, A.; Weißhaupt, S.; Krawiec, P.; Kunz, S. Use of Aureobasidium pullulans for resistance management in chemical control of Botrytis cinerea in berries. Acta Hortic. 2014, 1017, 237–242. [Google Scholar] [CrossRef]
- Weiss, A.; Mögel, G.; Kunz, S. Development of “Boni-Protect”—A yeast preparation for use in the control of postharvest diseases of apples. In Proceedings of the 12th International Conference on Cultivation Technique and Phytopathological Problems in Organic, Weinsberg, Germany, 31 January–2 February 2006; Fördergemeinschaft Ökologischer Obstbau e.V. (FÖKO): Weinsberg, Germany, 2006; pp. 1–5. [Google Scholar]
- del Fabbro, R.; Crivelli, L.; Lacertosa, G.; Digeronimo, G.; Calari, A.; Edler, B.; D’Ascenzo, D. Activity of Aureobasidium pullulans (Botector) against grey mold on grape, strawberry and tomato. Atti Giornate Fitopatol. Chianciano terme (Siena) 8-11 marzo 2016, 2, 231–239. [Google Scholar]
- Weißhaupt, S.; Hinze, M.; Weiß, A.; Kunz, S. Application of Boni Protect® against postharvest diseases in integrated apple production. IOBC-WPRS Bull. 2012, 84, 143–149. [Google Scholar]
- Garrido, C.C.; Usall, J.; Torres, R.; Teixidó, N. Effective control of Botrytis bunch rot in commercial vineyards by large-scale application of Candida sake CPA-1. BioControl 2017, 62, 161–173. [Google Scholar] [CrossRef]
- Calvo-Garrido, C.; Elmer, P.A.G.; Viñas, I.; Usall, J.; Bartra, E.; Teixidó, N. Biological control of botrytis bunch rot in organic wine grapes with the yeast antagonist Candida sake CPA-1. Plant Pathol. 2013, 62, 510–519. [Google Scholar] [CrossRef]
- Calvo-Garrido, C.; Viñas, I.; Usall, J.; Rodríguez-Romera, M.; Ramos, M.C.; Teixidó, N. Survival of the biological control agent Candida sake CPA-1 on grapes under the influence of abiotic factors. J. Appl. Microbiol. 2014, 117, 800–811. [Google Scholar] [CrossRef]
- Mbili, N.C.; Laing, M.D.; Yobo, K.S. Integrated control of Penicillium expansum and Botrytis cinerea of apples using potassium silicate, yeast antagonists and YieldPlus®. Acta Hortic. 2020, 1269, 75–79. [Google Scholar] [CrossRef]
- Kowalska, J.; Drozdzyński, D.; Remlein-Starosta, D.; Sas-Paszt, L.; Malusá, E. Use of Cryptococcus albidus for controlling grey mould in the production and storage of organically grown strawberries. J. Plant Dis. Prot. 2012, 119, 174–178. [Google Scholar] [CrossRef]
- Droby, S.; Cohen, L.; Daus, A.; Weiss, B.; Horev, B.; Chalutz, E.; Katz, H.; Keren-Tzur, M.; Shachnai, A. Commercial testing of aspire: A yeast preparation for the biological control of postharvest decay of citrus. Biol. Control. 1998, 12, 97–101. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Y.; Sui, Y.; Xie, Z.; Liu, Y.; Jiang, M.; Liu, J. Exposure of Candida oleophila to sublethal salt stress induces an antioxidant response and improves biocontrol efficacy. Biol. Control. 2018, 127, 109–115. [Google Scholar] [CrossRef]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.-C. Perspectives and Challenges of Microbial Application for Crop Improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef]
- De Simone, N.; Capozzi, V.; Amodio, M.L.; Colelli, G.; Spano, G.; Russo, P. Microbial-based Biocontrol Solutions for Fruits and Vegetables: Recent Insight, Patents, and Innovative Trends. Recent Pat. Food Nutr. Agric. 2021, 12, 3–18. [Google Scholar] [CrossRef]
- Bayer CropScience AG. Combinations of Biological Control Agents and Insecticides. U.S. Patent Application EP2269455A1, 2011. [Google Scholar]
- Droby, S. A Novel Antagonistic Yeast Useful in Controlling Spolage of Agricultural Produce, Methods of Ude Thereof and Compositions Containing Same. European Patent Application WO2002072777A2, 19 September 2002. [Google Scholar]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food Nutr. Agric. 2019, 11, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Raimi, A.; Roopnarain, A.; Adeleke, R. Biofertilizer production in Africa: Current status, factors impeding adoption and strategies for success. Sci. Afr. 2021, 11, e00694. [Google Scholar] [CrossRef]
- Barquero, M.; Pastor-Buies, R.; Urbano, B.; González-Andrés, F. Challenges, Regulations and Future Actions in Biofertilizers in the European Agriculture: From the Lab to the Field; Springer: Cham, Germany, 2019; pp. 83–107. [Google Scholar]


{kind=link}
| Mechanism | Attribute | Yeasts | Reference |
|---|---|---|---|
| N2 fixation | Plant growth promotion | C. tropicalis | [68] |
| NH3 production | Plant growth promotion | M. guilliermondii, P. rugulosa, C. flavus, P. antarctica,Meyerozyma sp., M. caribbica | [43,60,69] |
| P solubilization | Plant growth promotion | C. tropicalis, L. thermotolerans, Rhodotorula sp., H. uvarum, Y. lipolytica, S. cerevisiae | [32,68,69,77,78] |
| K solubilization | Plant growth promotion | T. globosa, R. glutinis, P. anomala | [54,80] |
| S oxidizing | Plant growth promotion | W. californica, S. cerevisiae | [84] |
| Zn solubilization | Plant growth promotion | C. tropicalis, L. thermotolerans, Dothideomycetes sp. | [43,68,69] |
| Polyamine production | Plant growth promotion | C. tropicalis, P. rugulosa, P. antarctica, A. pullulans, Dothideomycetes sp. | [32,43,60] |
| ACC deaminase activity | Plant growth promotion | C. tropicalis, Cryptococcus sp. | [32,71] |
| IAA production | Plant growth promotion | C. tropicalis, M. guilliermondii, S. cerevisiae, A. pullulans, H. uvarum,M. caribbica, W. californica, C. laurentii, R. fluvialis, C. maltosa, P. kudriavzevii, R. paludigenum | [43,65,68,69,89,90,91,92] |
| Cytokinin production | Plant growth promotion | A. pullulans, S. roseus, M. pulcherrima | [94] |
| Pulcherrimine production | Biofungicide | M. pulcherrima | [38,109] |
| Siderophore production | Biofungicide | R. glutinis against B. cinerea and P. expansum | [110] |
| Competition for nutrients and space | Biofungicide | A. pullulans against P. expansum and M. laxa; Issatchenkia terricola, P. anomala, M. pulcherrima, S. cerevisiae, Schizosaccharomyces pombe, H. uvarum and P. kluyveri against B. cinerea | [38,107,108,139,140,141,142,143] |
| Parasitism | Biofungicide | M. guilliermondii, P. anomala, C. famata and R. mucilaginosa against C. gloeosporioides; P. anomala, A. pullulans and P. kluyveri against B. cinerea | [38,118,119,140,143] |
| VOCs production | Biofungicide | A. pullulans, Hanseniaspora sp., P. anomala, M. pulcherrima, P. membranifaciens, M. guilliermondii, H. uvarum, Starmerella bacillaris, Candida pyralidae and P. kluyveri against B. cinerea | [38,112,113,140,143,144,145,146,147] |
| Killer activity | Biofungicide | P. membranifaciens, P. anomala, K. lactis | [115,116,117] |
| Induced systemic resistance | Biofungicide | R. paludigenum against P. digitatum; P. anomala against P. expansum | [123,124] |
| Insecticide biodegradation | Biodegrader | R. glutinis and R. rubra degrades chlorpyrifos; R. mucilaginosa degrades neonicotinoid insecticide; S. terricola degrades glyphosate insecticide | [31,126,135] |
| Herbicide biodegradation | Biodegrader | C. lusitaniae degrades dinitroaniline herbicide; P. kudriavzevii degrades s-triazine group herbicides | [136,137] |
| Insecticidal activity | Insecticide | Y. lipolytica, A. pullulans, H. uvarum, M. pulcherrima, C. tephrensis | [128,129,131,132] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Fernández, M.; Cordero-Bueso, G.; Ruiz-Muñoz, M.; Cantoral, J.M. Culturable Yeasts as Biofertilizers and Biopesticides for a Sustainable Agriculture: A Comprehensive Review. Plants 2021, 10, 822. https://doi.org/10.3390/plants10050822
Hernández-Fernández M, Cordero-Bueso G, Ruiz-Muñoz M, Cantoral JM. Culturable Yeasts as Biofertilizers and Biopesticides for a Sustainable Agriculture: A Comprehensive Review. Plants. 2021; 10(5):822. https://doi.org/10.3390/plants10050822
Chicago/Turabian StyleHernández-Fernández, María, Gustavo Cordero-Bueso, Marina Ruiz-Muñoz, and Jesús M. Cantoral. 2021. "Culturable Yeasts as Biofertilizers and Biopesticides for a Sustainable Agriculture: A Comprehensive Review" Plants 10, no. 5: 822. https://doi.org/10.3390/plants10050822
APA StyleHernández-Fernández, M., Cordero-Bueso, G., Ruiz-Muñoz, M., & Cantoral, J. M. (2021). Culturable Yeasts as Biofertilizers and Biopesticides for a Sustainable Agriculture: A Comprehensive Review. Plants, 10(5), 822. https://doi.org/10.3390/plants10050822