
Detection and Distribution of Viruses Infecting Garlic Crops in Australia





Abstract
1. Introduction
2. Results
2.1. Molecular Detection of Garlic Viruses
2.2. Field Surveys
2.3. Evaluation of Planting Propagules for Virus Distribution
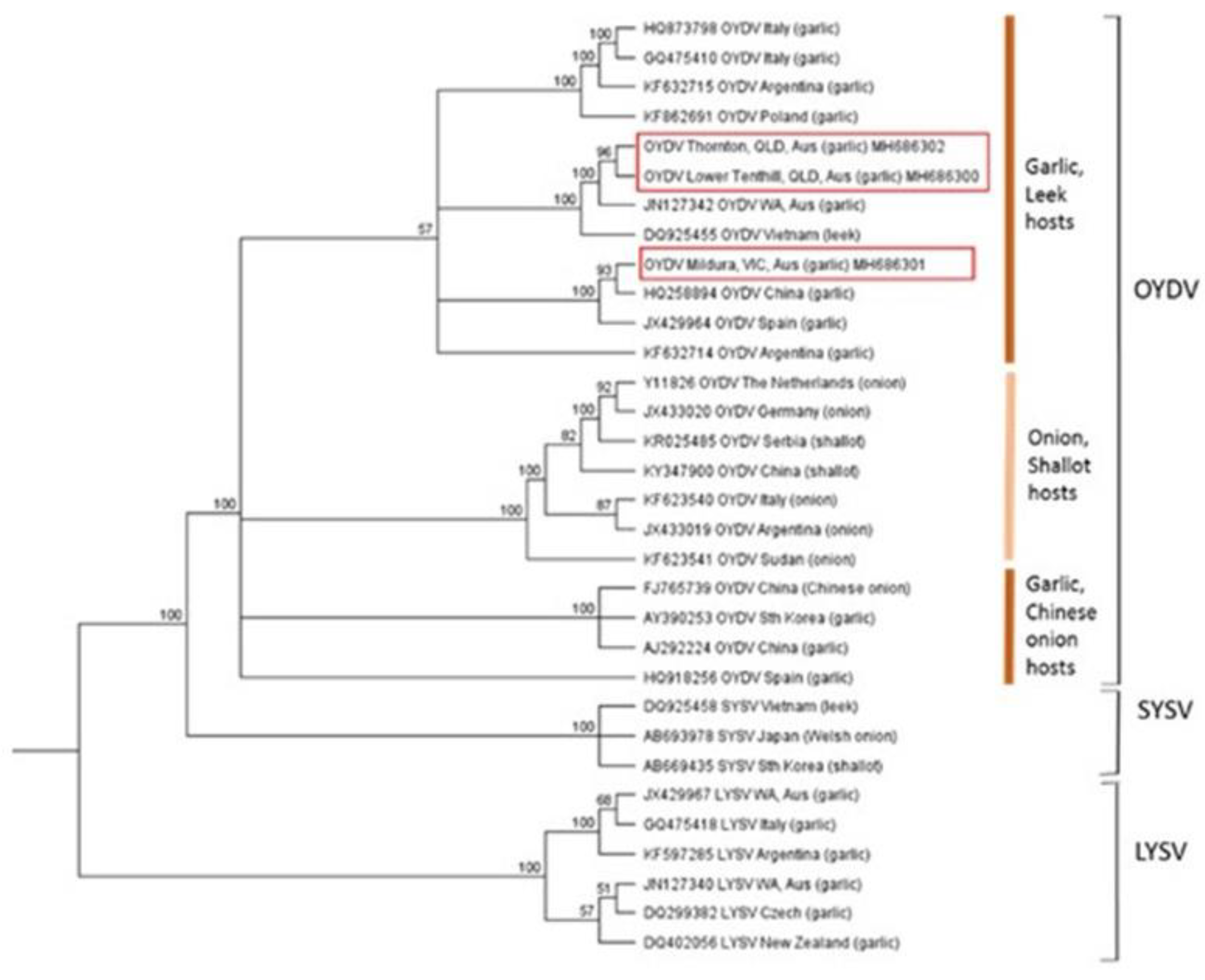
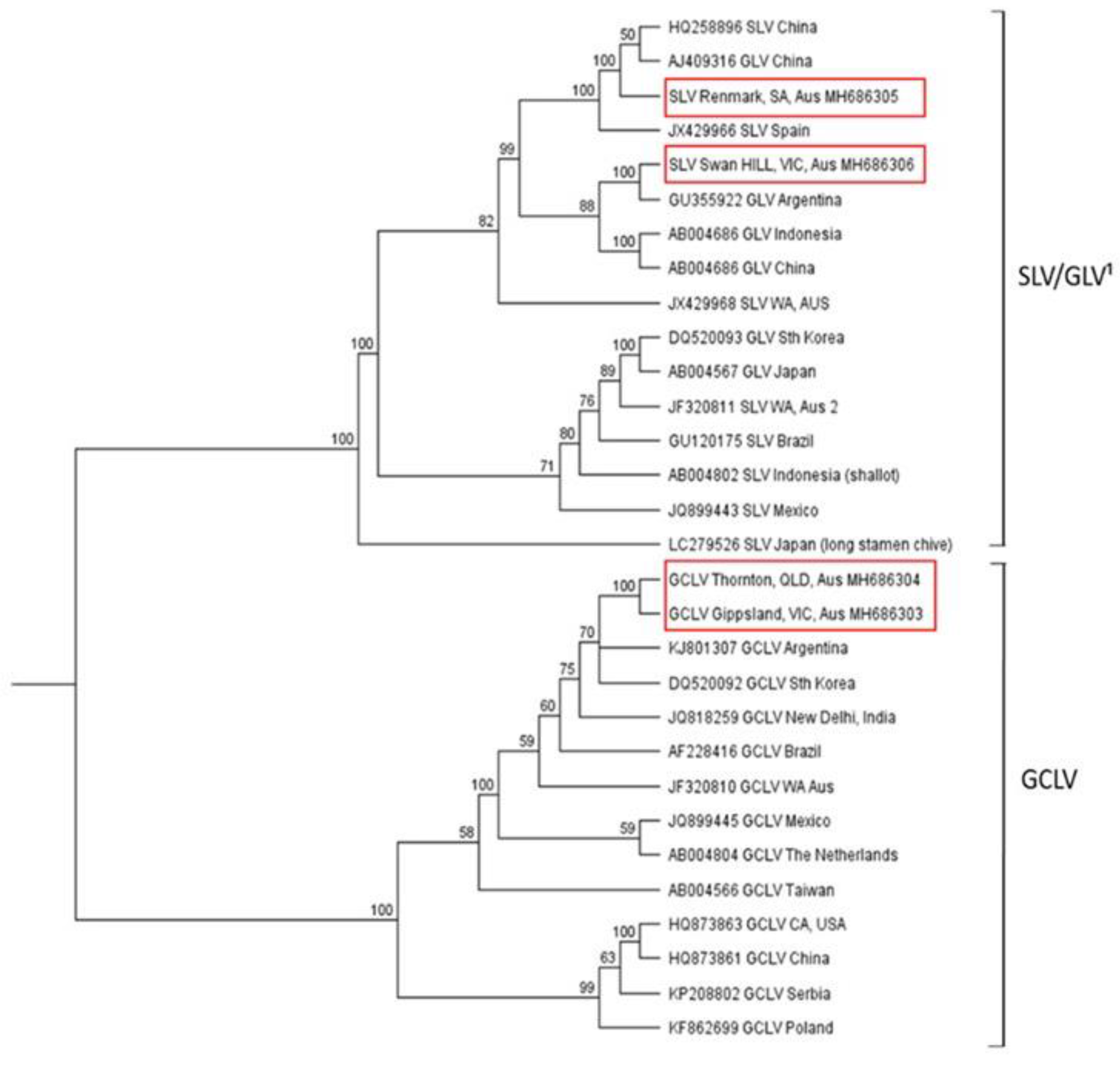
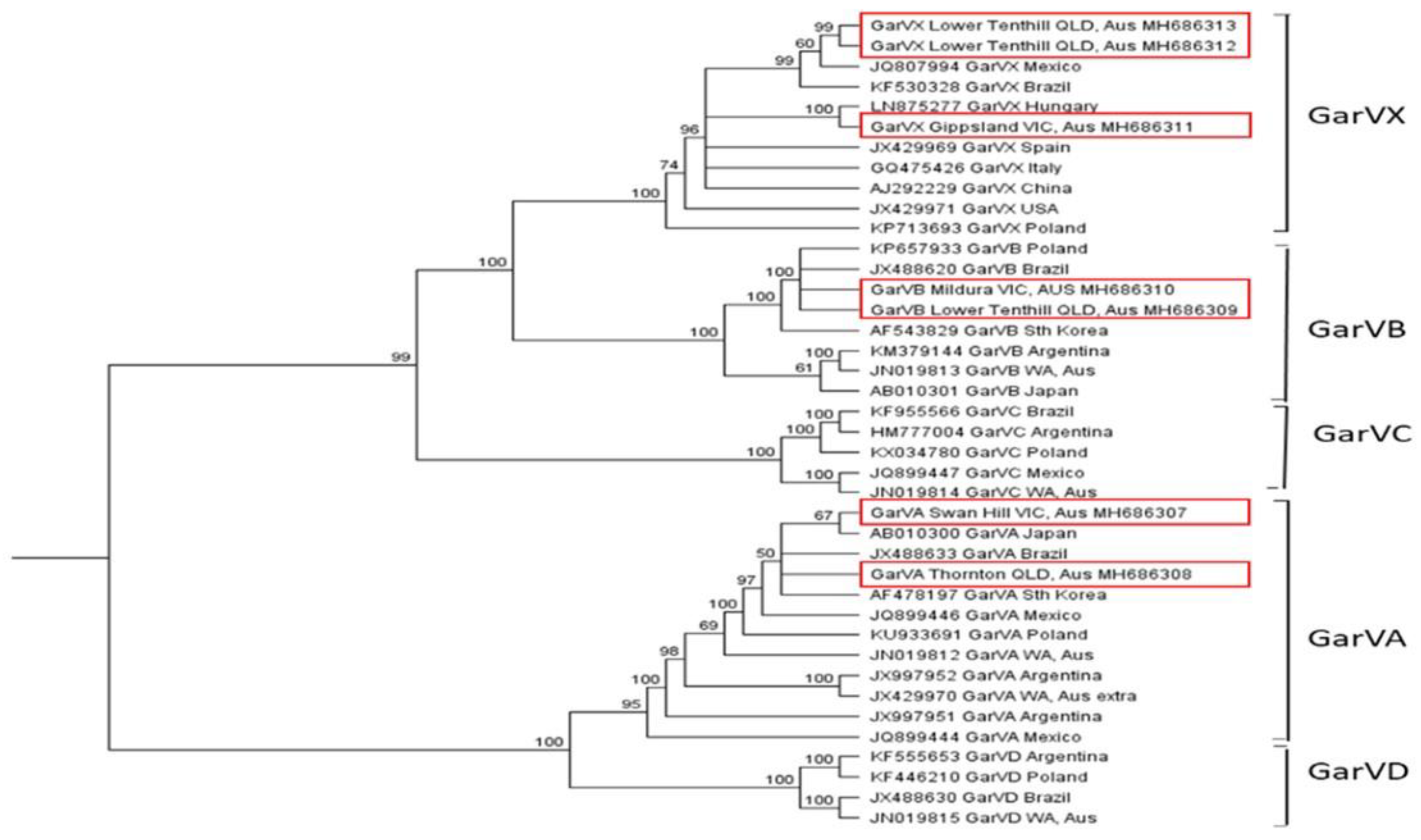
2.4. Genetic Diversity and Phylogenetic Analysis of Viruses Present in Australian Garlic Crops
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Virus Indexing
5.2. Field Surveys
5.3. Evaluation of Planting Propagules for Virus Distribution
5.4. Phylogenetic Analysis of the Virus Coat Protein Gene
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andreeva, I. The Australian Garlic Trade: Main Trends. World: Garlic. Market Report Analysis and Forecast to 2020. Available online: https://garlicaustralia.asn.au/wp-content/uploads/2020/02/IndexBox_Marketing-Garlic.pdf (accessed on 9 April 2021).
- Cafrune, E.E.; Perotto, M.C.; Conci, V.C. Effect of two Allexivirus isolates on garlic yield. Plant Dis. 2006, 90, 898–904. [Google Scholar] [CrossRef]
- Conci, V.C.; Canavelli, A.; Lunello, P.; Rienzo, J.D.; Nome, S.F.; Zumelzu, G.; Italia, R. Yield losses associated with virus-infected garlic plants during five successive years. Plant Dis. 2003, 87, 1411–1415. [Google Scholar] [CrossRef]
- Lunello, P.; Rienzo, J.D.; Conci, V.C. Yield loss in garlic caused by leek yellow stripe virus Argentinean isolate. Plant Dis. 2007, 91, 153–158. [Google Scholar] [CrossRef]
- Perotto, M.C.; Cafrune, E.E.; Conci, V.C. The effect of additional viral infections on garlic plants initially infected with Allexiviruses. Eur. J. Plant Pathol. 2010, 126, 489–495. [Google Scholar] [CrossRef]
- Nam, M.; Lee, Y.-H.; Park, C.Y.; Lee, M.-A.; Bae, Y.-S.; Lim, S.; Lee, J.H.; Moon, J.S.; Lee, S.-H. Development of multiplex RT-PCR for simultaneous detection of garlic viruses and the incidence of garlic viral disease in garlic genetic resources. Plant Pathol. J. 2015, 31, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Lot, H.; Chovelon, V.; Souche, S.; Delecolle, B. Effects of onion yellow dwarf and leek yellow stripe viruses on symptomatology and yield loss of three french garlic cultivars. Plant Dis. 1998, 82, 1381–1385. [Google Scholar] [CrossRef] [PubMed]
- Katis, N.; Maliogka, V.I.; Dovas, C.I. Viruses of the genus Allium in the Mediterranean Region. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2012; pp. 163–208. [Google Scholar]
- Chen, J.; Chen, J.; Adams, M.J. A universal PCR primer to detect members of the Potyviridae and its use to examine the taxonomic status of several members of the family. Arch. Virol. 2001, 146, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Tsuneyoshi, T.; Matsumi, T.; Deng, D.; Sako, I.; Sumi, S. Differentiation of allium carlaviruses isolated from different parts of the world based on the viral coat protein sequence. Arch. Virol. 1998, 143, 1093–1107. [Google Scholar] [CrossRef] [PubMed]
- Filho, M.E.L.O.; Nagata, T.; Dusi, A.N.; Buso, J.A.; Torres, A.C.; Eiras, M.; Resende, R.D.O. Detection of three Allexivirus species infecting garlic in Brazil. Pesqui. Agropecu. Bras. 2004, 39, 735–740. [Google Scholar] [CrossRef]
- Song, S.I.; Song, J.T.; Chang, M.U.; Lee, J.S.; Choi, Y.D. Identification of one of the major viruses infecting garlic plants, garlic virus X. Mol. Cells 1997, 7, 705–709. [Google Scholar]
- Sumi, S.; Tsuneyoshi, T.; Furutani, H. Novel rod-shaped viruses isolated from garlic, allium sativum, possessing a unique genome organization. J. Gen. Virol. 1993, 74, 1879–1885. [Google Scholar] [CrossRef]
- Kritzman, A.; Lampel, M.; Raccah, B.; Gera, A. Distribution and transmission of Iris yellow spot virus. Plant Dis. 2001, 85, 838–842. [Google Scholar] [CrossRef]
- Bag, S.; Schwartz, H.F.; Cramer, C.S.; Havey, M.J.; Pappu, H.R. Iris yellow spot virus (Tospovirus: Bunyaviridae): From obscurity to research priority. Mol. Plant Pathol. 2015, 16, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Bereda, M.; Paduch-Cichal, E.; Dąbrowska, E. Occurrence and phylogenetic analysis of allexiviruses identified on garlic from China, Spain and Poland commercially available on the Polish retail market. Eur. J. Plant Pathol. 2017, 149, 227–237. [Google Scholar] [CrossRef]
- Pramesh, D.; Baranwal, V.K. Molecular characterization of coat protein gene of garlic common latent virus isolates from India: An evidence for distinct phylogeny and recombination. Virus Genes 2013, 47, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Takaki, F.; Sano, T.; Yamashita, K.; Fujita, T.; Ueda, K.; Kato, T. Complete nucleotide sequences of attenuated and severe isolates of leek yellow stripe virus from garlic in northern Japan: Identification of three distinct virus types in garlic and leek world-wide. Arch. Virol. 2005, 150, 1135–1149. [Google Scholar] [CrossRef]
- Gieck, S.L.; Hamm, P.B.; David, N.L.; Pappu, H.R. First Report of garlic virus B and garlic virus D in garlic in the Pacific Northwest. Plant Dis. 2009, 93, 431. [Google Scholar] [CrossRef]
- Taglienti, A.; Tiberini, A.; Manglli, A.; Rea, R.; Paoletti, S.; Taviani, P.; Tomassoli, L. Molecular Identification of Allexiviruses in a complex mixture of garlic viruses in Latium (Central Italy. Eur. J. Plant Pathol. 2018, 150, 797–801. [Google Scholar] [CrossRef]
- Sward, R. Lettuce necrotic yellows rhabdovirus and other viruses infecting garlic. Australas. Plant Pathol. 1990, 19, 46. [Google Scholar] [CrossRef]
- Sward, R.J. Development of Methods for the Rapid Detection and Elimination of Virus Diseases of Garlic; Horticultural Research and Development Corporation: Gordon, NSW, Australia, 1996; ISBN 978-1-86423-404-6. [Google Scholar]
- Sward, R.J.; Brennan, A.P. Diagnosis and control of allium virus diseases in Victoria, Australia 1994. Acta Hortic. 1994, 358, 295–298. [Google Scholar] [CrossRef]
- Wei, T.; Pearson, M.N.; Fletcher, J.D. Molecular confirmation of New Zealand garlic yellow streak virus as leek yellow stripe virus. Australas. Plant Pathol. 2006, 35, 341–346. [Google Scholar] [CrossRef]
- Wylie, S.J.; Li, H.; Saqib, M.; Jones, M.G.K. The global trade in fresh produce and the vagility of plant viruses: A case study in garlic. PLoS ONE 2014, 9, e105044. [Google Scholar] [CrossRef] [PubMed]
- Wylie, S.; Luo, H.; Li, H.; Jones, M.K. Multiple polyadenylated RNA viruses detected in pooled cultivated and wild plant samples. Arch. Virol. 2012, 157, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Coutts, B.A.; McMichael, L.A.; Tesoriero, L.; Rodoni, B.C.; Wilson, C.R.; Wilson, A.J.; Persley, D.M.; Jones, R.A.C. Iris yellow spot virus found infecting onions in three Australian states. Australas. Plant Pathol. 2003, 32, 555–557. [Google Scholar] [CrossRef]
- Celli, M.G.; Torrico, A.K.; Kiehr, M.; Conci, V.C. Striking differences in the biological and molecular properties of onion and garlic isolates of onion yellow dwarf virus. Arch. Virol. 2013, 158, 1377–1382. [Google Scholar] [CrossRef]
- Pozzer, L.; Bezerra, I.C.; Kormelink, R.; Prins, M.; Peterse, D.; Resende, R.D.O.; de Ávila, A.C. Characterization of a tospovirus isolate of Iris yellow spot virus associated with a disease in onion fields in Brazil. Plant Dis. 1999, 83, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Moran, J.; Garrett, R.G.; Fairweather, J.V. Strategy for detecting low levels of potato viruses X and S in crops and its application to the Victorian certified potato scheme. Plant Dis. 1983, 67, 1325–1328. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survey Site | Location | Variety | Number of Samples | Allexiviruses | GarVA | GarVB | GarVC | GarVX | ShVX | Carlaviruses | GCLV | SLV | Potyviruses | OYDV | SYSV | LYSV |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WP285 | Thornton, QLD | Glenlarge | 238 | 100.0 | 66.4 | 62.6 | 2.9 | 80.3 | 0.0 | 99.6 | 2.9 | 100.0 | 99.6 | 97.9 | 0.0 | 97.5 |
| WP286 | Lower Tent Hill, QLD | Glenlarge | 94 | 98.9 | 65.6 | 93.8 | 30.2 | 62.5 | 0.0 | 98.9 | 0.0 | 89.6 | 98.9 | 94.8 | 0.0 | 87.5 |
| WP287 | Kalbar, QLD | Glenlarge | 90 | 99.0 | 52.2 | 91.1 | 0.0 | 30.0 | 0.0 | 100.0 | 98.9 | 100.0 | 91.1 | 98.9 | 0.0 | 100.0 |
| WP288 | Mt Sylvia, QLD | Glenlarge | 94 | 94.0 | 84.0 | 92.6 | 3.2 | 61.7 | 0.0 | 98.9 | 89.4 | 80.9 | 91.5 | 67 | 0.0 | 77.7 |
| Site 1 | East Gippsland, VIC | Purple stripe | 30 | 100.0 (19.4–100) | 100.0 (19.4–100) | 0.0 (0–1.2) | 0.7 (0.08–1.9) | 28.8 (16.1–51) | 0.0 (0–1.2) | 100.0 (19.4–100) | 13.5 (8.2–20.6) | 100.0 (19.4–100) | 20.6 (12.4–30.0) | 20.6 (12.4–32) | 0.0 (0–1.2) | 4.0 (1.9–7.2) |
| Site 2 | East Gippsland, VIC | not known | 30 | 23.7 (14.0–38.1) | 0.0 (0–1.2) | 20.6 (12.4–32) | 2.6 (1.04–5.3) | 0.0 (0–1.2) | 0.0 (0–1.2) | 23.7 (14.0–38.1) | 11.3 (6.8–17.4) | 16.4 (10–25) | 100.0 (19.4–100) | 100.0 (19.4–100) | 0 (0–1.2) | 14.9 (9.1–22.6) |
| Site 3 | Swan Hill, VIC | Chinese white | 30 | 100.0 (19.4–100) | 0.0 (0–1.2) | 14.9 (9.1–22.6) | 0.7 (0.1–1.9) | 100.0 (19.4–100) | 0.0 (0–1.2) | 100.0 (19.4–100) | 100.0 (19.4–100) | 100.0 (19.4–100) | 100.0 (19.4–100) | 100.0 (19.4–100) | 0.0 (0–1.2) | 3.8 (5.1–13.8) |
| Site 4 | Merbein, VIC | Red | 30 | 100.0 (19.4–100) | 0.0 (0–1.2) | 18.2 (11.1–28) | 1.05 (0.2–3.03) | 100.0 (19.4–100) | 0.0 (0–1.2) | 100.0 (19.4–100) | 100.0 (19.4–100) | 100.0 (19.4–100) | 28.8 (16.1–51.0) | 16.4 (10–25) | 0.0 (0–1.2) | 0.0 (0–1.2) |
| Site 5 | Merbein, VIC | not known | 30 | 28.8 (16.1–51.0) | 0.0 (0–1.2) | 20.6 (12.4–32) | 0.7 (0.08–1.9) | 28.8 (16.1–51) | 0.0 (0–1.2) | 100.0 (19.4–100) | 3.5 (1.6–6.6) | 100.0 (19.4–100) | 28.8 (16.1–51.0) | 23.7 (14–38.1) | 0.0 (0–1.2) | 4.0 (1.9–7.2) |
| Site 6 | Mildura, VIC | Red | 30 | 16.4 (10.0–25.0) | 0.0 (0–1.2) | 14.9 (9.1–22.6) | 0.3 (0.01–1.9) | 13.5 (8.2–20.6) | 0.0 (0–1.2) | 100.0 (19.4–100) | 28.8 (16.1–51) | 100.0 (19.4–100) | 100.0 (19.4–100) | 28.8 (16.1–51) | 0.0 (0–1.2) | 5.0 (2.5–8.6) |
| Site 7 | Mildura, VIC | not known | 30 | 28.8 (16.1–51.0) | 0.0 (0–1.2) | 20.6 (12.4–32) | 0.0 (0–1.2) | 28.8 (16.1–51) | 0.0 (0–1.2) | 100.0 (19.4–100) | 100.0 (19.4–100) | 100.0 (19.4–100) | 28.8 (16.1–51.0) | 28.8 (16.1–51) | 0.0 (0–1.2) | 2.2 (0.8–4.8) |
| Site 8 | Daylesford, VIC | Purple stripe | 30 | 28.8 (16.1–51.0) | 2.0 (0.8–4.8) | 0.0 (0–1.2) | 14.9 (9.1–22.6) | 18.2 (11.1–28) | 0.0 (0–1.2) | 100.0 (19.4–100) | 3.1 (1.3–6) | 100.0 (19.4–100) | 100.0 (19.4–100) | 28.8 (16.1–51) | 0.0 (0–1.2) | 8.2 (11.1–28) |
| Site 9 | Buronga, NSW | Various | 30 | 100.0 (19.4–100) | 0.0 (0–1.2) | 6.4 (10–25) | 0.0 (0–1.2) | 0.3 (0.01–1.9) | 0.0 (0–1.2) | 100.0 (19.4–100) | 18.2 (11.1–28) | 100.0 (19.4–100) | 100.0 (19.4–100) | 100.0 (19.4–100) | 0.0 (0–1.2) | 3.5 (1.6–6.6) |
| Site 10 | Renmark, SA | Purple | 30 | 5.0 (2.5–8.6) | 0.0 (0–1.2) | 0.0 (0–1.2) | 0.0 (0–1.2) | 4.45 (2.2–7.9) | 0.0 (0–1.2) | 100.0 (19.4–100) | 0.0 (0–1.2) | 100.0 (19.4–100) | 100.0 (19.4–100) | 28.8 (16.1–51) | 0.0 (0–1.2) | 1.8 (0.6–4.2) |
| Bulb Quality | Bulb Reference | Number of Cloves Evaluated | GarVA | GarVB | GarVX | SLV | OYDV | LYSV |
|---|---|---|---|---|---|---|---|---|
| Small | 1 | 9 | 71 | 86 | 71 | 86 | 86 | 86 |
| 2 | 10 | 100 | 70 | 70 | 90 | 90 | 80 | |
| 3 | 4 | 100 | 100 | 100 | 100 | 100 | 100 | |
| 4 | 6 | 100 | 100 | 100 | 100 | 100 | 100 | |
| 5 | 4 | 100 | 100 | 100 | 100 | 100 | 100 | |
| 6 | 12 | 100 | 83 | 100 | 92 | 83 | 83 | |
| 7 | 10 | 90 | 100 | 100 | 100 | 100 | 100 | |
| 8 | 3 | 100 | 100 | 100 | 100 | 100 | 100 | |
| 9 | 4 | 100 | 100 | 100 | 100 | 100 | 100 | |
| 10 | 11 | 64 | 73 | 73 | 100 | 100 | 100 | |
| 11 | 4 | 50 | 25 | 75 | 75 | 100 | 100 | |
| 12 | 6 | 67 | 67 | 83 | 100 | 100 | 83 | |
| 13 | 4 | 100 | 100 | 100 | 100 | 100 | 100 | |
| 14 | 5 | 60 | 60 | 60 | 80 | 100 | 100 | |
| Large | 1 | 17 | 100 | 100 | 82 | 100 | 100 | 100 |
| 2 | 15 | 100 | 100 | 93 | 100 | 93 | 100 | |
| 3 | 16 | 100 | 100 | 100 | 100 | 100 | 100 | |
| 4 | 18 | 94 | 94 | 94 | 94 | 100 | 94 | |
| 5 | 12 | 92 | 92 | 92 | 92 | 100 | 92 | |
| 6 | 15 | 100 | 100 | 80 | 87 | 100 | 100 | |
| 7 | 17 | 94 | 94 | 94 | 94 | 100 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cremer, J.; Campbell, P.; Steele, V.; Persley, D.; Thomas, J.; Harper, S.; Gambley, C. Detection and Distribution of Viruses Infecting Garlic Crops in Australia. Plants 2021, 10, 1013. https://doi.org/10.3390/plants10051013
Cremer J, Campbell P, Steele V, Persley D, Thomas J, Harper S, Gambley C. Detection and Distribution of Viruses Infecting Garlic Crops in Australia. Plants. 2021; 10(5):1013. https://doi.org/10.3390/plants10051013
Chicago/Turabian StyleCremer, Julia, Paul Campbell, Visnja Steele, Denis Persley, John Thomas, Stephen Harper, and Cherie Gambley. 2021. "Detection and Distribution of Viruses Infecting Garlic Crops in Australia" Plants 10, no. 5: 1013. https://doi.org/10.3390/plants10051013
APA StyleCremer, J., Campbell, P., Steele, V., Persley, D., Thomas, J., Harper, S., & Gambley, C. (2021). Detection and Distribution of Viruses Infecting Garlic Crops in Australia. Plants, 10(5), 1013. https://doi.org/10.3390/plants10051013