Circadian Variation of Root Water Status in Three Herbaceous Species Assessed by Portable NMR

 ,
,  and
and 
Abstract
1. Introduction
2. Results
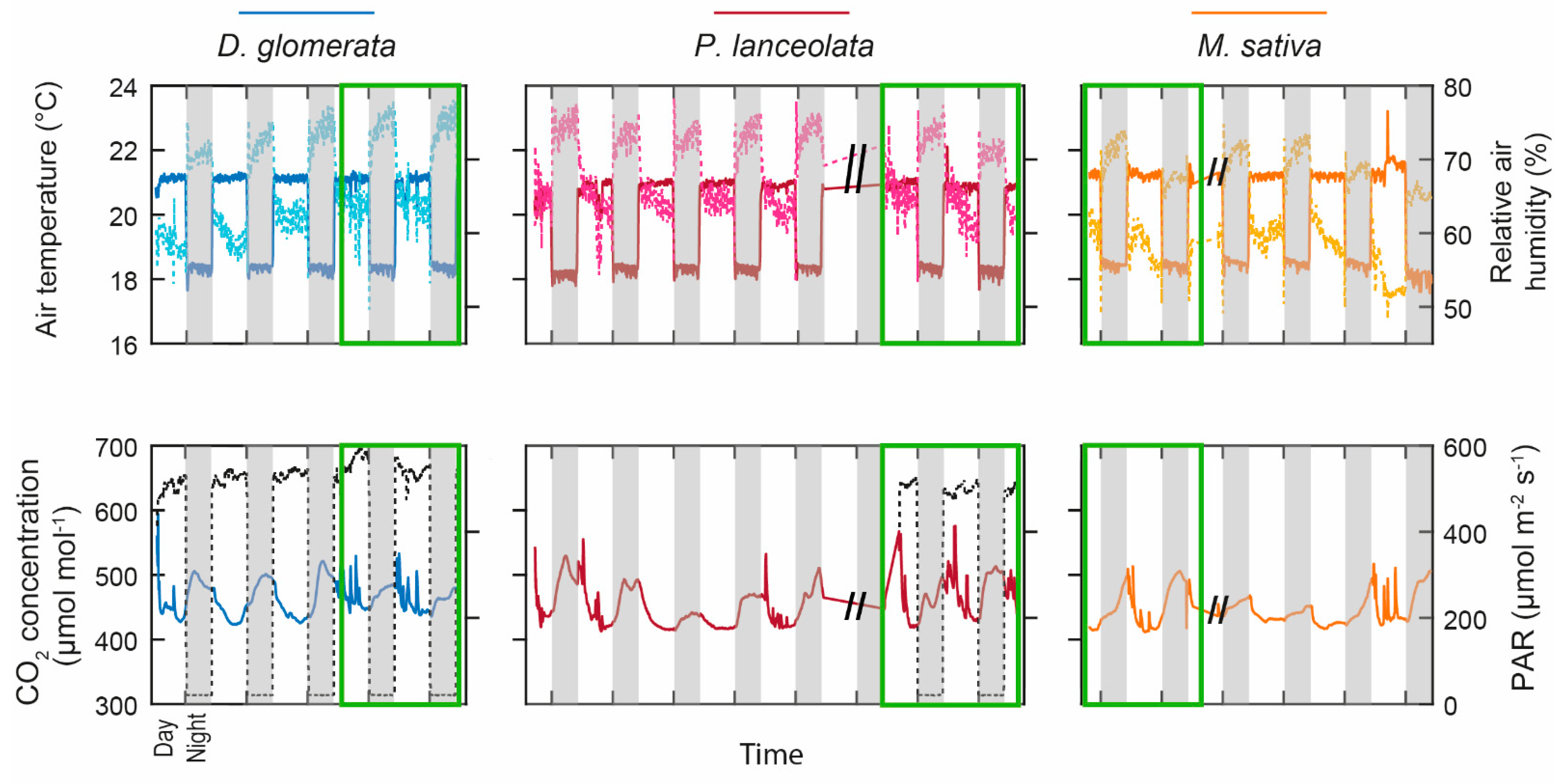
2.1. Climatic Chamber
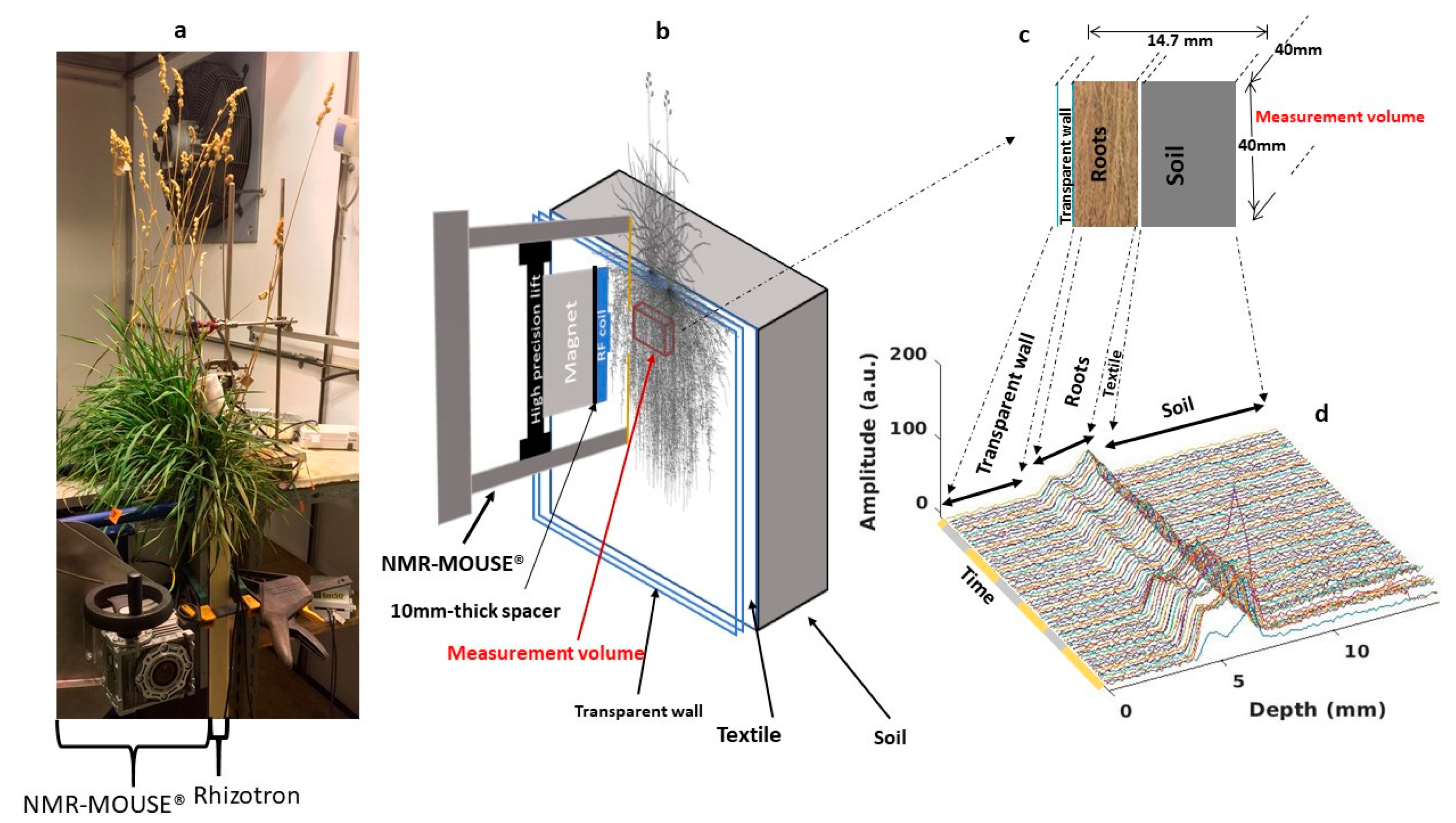
2.2. NMR Profiles within the Three Rhizotrons
2.3. Circadian Ecophysiological Measurements and NMR Signals in Roots and Soil
2.4. Root Morphological Traits and Leaf Area
2.5. T2 Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.1.1. Rhizotrons
4.1.2. Plant Material
4.1.3. Climatic Chamber
4.2. Ecophysiological Measurements
4.2.1. Leaf Water Potential
4.2.2. Soil Humidity
4.2.3. Destructive Samplings
4.2.4. Root Morphology
4.3. NMR Experiments and Signal Analysis
4.3.1. NMR-MOUSE System
4.3.2. Intensity Profile Measurements and Signal Analysis
4.3.3. T2 Measurements and Fitting
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedlingstein, P.; Jones, M.W.; O’Sullivan, M.; Andrew, R.M.; Hauck, J.; Peters, G.P.; Peters, W.; Pongratz, J.; Sitch, S.; Le Quéré, C.; et al. Global Carbon Budget 2019. Earth Syst. Sci. Data 2019, 11, 1783–1838. [Google Scholar] [CrossRef]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Dangal, S.R.S.; Tian, H.; Pan, S.; Zhang, L.; Xu, R. Greenhouse Gas Balance in Global Pasturelands and Rangelands. Environ. Res. Lett. 2020, 15, 104006. [Google Scholar] [CrossRef]
- IPCC. Land Use, Land-Use Change, and Forestry: Summary for Policymakers: A Special Report of the Intergovernmental Panel on Climate Change; WMO (World Meteorological Organization): Geneva, Switzerland; UNEP (United Nations Environment Programme): Geneva, Switzerland, 2000; ISBN 978-92-9169-114-2. [Google Scholar]
- Jones, H.G. Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology, 3rd ed.; Cambridge University Press: Cambridge, UK, 2014; ISBN 978-0-521-27959-8. [Google Scholar]
- Windt, C.W.; Vergeldt, F.J.; De Jager, P.A.; Van As, H. MRI of Long-Distance Water Transport: A Comparison of the Phloem and Xylem Flow Characteristics and Dynamics in Poplar, Castor Bean, Tomato and Tobacco. Plant Cell Environ. 2006, 29, 1715–1729. [Google Scholar] [CrossRef]
- Garrigues, E.; Doussan, C.; Pierret, A. Water Uptake by Plant Roots: I–Formation and Propagation of a Water Extraction Front in Mature Root Systems as Evidenced by 2D Light Transmission Imaging. Plant Soil 2006, 283, 83–98. [Google Scholar] [CrossRef]
- Doussan, C.; Pierret, A.; Garrigues, E. Water Uptake by Plant Roots: II–Modelling of Water Transfer in the Soil Root-System with Explicit Account of Flow within the Root System–Comparison with Experiments. Plant Soil 2006, 283, 99–117. [Google Scholar] [CrossRef]
- Zarebanadkouki, M.; Kim, Y.X.; Carminati, A. Where Do Roots Take up Water? Neutron Radiography of Water Flow into the Roots of Transpiring Plants Growing in Soil. New Phytol. 2013, 199, 1034–1044. [Google Scholar] [CrossRef]
- Tötzke, C.; Kardjilov, N.; Manke, I.; Oswald, S.E. Capturing 3D Water Flow in Rooted Soil by Ultra-Fast Neutron Tomography. Sci. Rep. 2017, 7, 6192. [Google Scholar] [CrossRef]
- Metzner, R.; Eggert, A.; van Dusschoten, D.; Pflugfelder, D.; Gerth, S.; Schurr, U.; Uhlmann, N.; Jahnke, S. Direct Comparison of MRI and X-Ray CT Technologies for 3D Imaging of Root Systems in Soil: Potential and Challenges for Root Trait Quantification. Plant Methods 2015, 11, 17. [Google Scholar] [CrossRef]
- Pohlmeier, A.; Oros-Peusquens, A.; Javaux, M.; Menzel, M.I.; Vanderborght, J.; Kaffanke, J.; Romanzetti, S.; Lindenmair, J.; Vereecken, H.; Shah, N.J. Changes in Soil Water Content Resulting from Ricinus Root Uptake Monitored by Magnetic Resonance Imaging. Vadose Zone J. 2008, 7, 1010–1017. [Google Scholar] [CrossRef]
- Pohlmeier, A.; Vergeldt, F.; Gerkema, E. MRI in Soils: Determination of Water Content Changes Due to Root Water Uptake by Means of a Multi-Slice-Multi-Echo Sequence (MSME). Open Magn. Reson. J. 2010, 3, 69–74. [Google Scholar] [CrossRef]
- Gruwel, M.L.H. In Situ Magnetic Resonance Imaging of Plant Roots. Vadose Zone J. 2014, 13, 1–8. [Google Scholar] [CrossRef]
- Pflugfelder, D.; Metzner, R.; van Dusschoten, D.; Reichel, R.; Jahnke, S.; Koller, R. Non-Invasive Imaging of Plant Roots in Different Soils Using Magnetic Resonance Imaging (MRI). Plant Methods 2017, 13, 102. [Google Scholar] [CrossRef]
- Haber-Pohlmeier, S.; Tötzke, C.; Oswald, S.E.; Lehmann, E.; Blümich, B.; Pohlmeier, A. Imaging of Root Zone Processes Using MRI T 1 Mapping. Microporous Mesoporous Mater. 2018, 269, 43–46. [Google Scholar] [CrossRef]
- Haber-Pohlmeier, S.; Tötzke, C.; Lehmann, E.; Kardjilov, N.; Pohlmeier, A.; Oswald, S.E. Combination of Magnetic Resonance Imaging and Neutron Computed Tomography for Three-Dimensional Rhizosphere Imaging. Vadose Zone J. 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Van As, H.; Scheenen, T.; Vergeldt, F.J. MRI of Intact Plants. Photosynth. Res. 2009, 102, 213–222. [Google Scholar] [CrossRef]
- Van As, H.; van Duynhoven, J. MRI of Plants and Foods. J. Magn. Reson. 2013, 229, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Barigah, T.S.; Bonhomme, M.; Lopez, D.; Traore, A.; Douris, M.; Venisse, J.-S.; Cochard, H.; Badel, E. Modulation of Bud Survival in Populus Nigra Sprouts in Response to Water Stress-Induced Embolism. Tree Physiol. 2013, 33, 261–274. [Google Scholar] [CrossRef]
- Capitani, D.; Brilli, F.; Mannina, L.; Proietti, N.; Loreto, F. In Situ Investigation of Leaf Water Status by Portable Unilateral Nuclear Magnetic Resonance. Plant Physiol. 2009, 149, 1638–1647. [Google Scholar] [CrossRef] [PubMed]
- Leca, A.; Clerjon, S.; Bonny, J.-M.; Renard, C.; Traoré, A. Multiscale NMR Analysis of the Degradation of Apple Structure Due to Thermal Treatment. J. Food Eng. 2021, 294, 110413. [Google Scholar] [CrossRef]
- Koizumi, M.; Naito, S.; Ishida, N.; Haishi, T.; Kano, H. A Dedicated MRI for Food Science and Agriculture. FSTR 2008, 14, 74–82. [Google Scholar] [CrossRef][Green Version]
- Meixner, M.; Tomasella, M.; Foerst, P.; Windt, C.W. A Small-scale MRI Scanner and Complementary Imaging Method to Visualize and Quantify Xylem Embolism Formation. New Phytol. 2020, 226, 1517–1529. [Google Scholar] [CrossRef]
- Van As, H.; Reinders, J.E.A.; de Jager, P.A.; van de Sanden, P.A.C.M.; Schaafsma, T.J. In Situ Plant Water Balance Studies Using a Portable NMR Spectrometer. J. Exp. Bot. 1994, 45, 61–67. [Google Scholar] [CrossRef]
- Homan, N.M.; Windt, C.W.; Vergeldt, F.J.; Gerkema, E.; Van As, H. 0.7 and 3 T MRI and Sap Flow in Intact Trees: Xylem and Phloem in Action. Appl. Magn. Reson. 2007, 32, 157–170. [Google Scholar] [CrossRef]
- Windt, C.W. A Portable Halbach Magnet That Can Be Opened and Closed without Force: The NMR-CUFF. J. Magn. Reson. 2011, 208, 27–33. [Google Scholar] [CrossRef]
- Windt, C.W.; Blümler, P. A Portable NMR Sensor to Measure Dynamic Changes in the Amount of Water in Living Stems or Fruit and Its Potential to Measure Sap Flow. Tree Physiol. 2015, 35, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Geya, Y.; Terada, Y.; Kose, K.; Haishi, T.; Gemma, H.; Sekozawa, Y. Development of a Mobile Magnetic Resonance Imaging System for Outdoor Tree Measurements. Rev. Sci. Instrum. 2011, 82, 053704. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Aptaker, P.S.; Cox, J.; Gardiner, B.A.; McDonald, P.J. A Transportable Magnetic Resonance Imaging System for in Situ Measurements of Living Trees: The Tree Hugger. J. Magn. Reson. 2012, 218, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Nagata, A.; Kose, K.; Terada, Y. Development of an Outdoor MRI System for Measuring Flow in a Living Tree. J. Magn. Reson. 2016, 265, 129–138. [Google Scholar] [CrossRef]
- Eidmann, G.; Savelsberg, R.; Blümler, P.; Blümich, B. The NMR MOUSE, a Mobile Universal Surface Explorer. J. Magn. Reson. Ser. A 1996, 122, 104–109. [Google Scholar] [CrossRef]
- Bagnall, G.C.; Koonjoo, N.; Altobelli, S.A.; Conradi, M.S.; Fukushima, E.; Kuethe, D.O.; Mullet, J.E.; Neely, H.; Rooney, W.L.; Stupic, K.F.; et al. Low-Field Magnetic Resonance Imaging of Roots in Intact Clayey and Silty Soils. Geoderma 2020, 370, 114356. [Google Scholar] [CrossRef]
- Bonny, J.; Traore, A.; Bouhrara, M.; Spencer, R.G.; Pages, G. Parsimonious Discretization for Characterizing Multi-exponential Decay in Magnetic Resonance. NMR Biomed. 2020, 33. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, J.A.; Pound, M.P.; Bennett, M.J.; Wells, D.M. Uncovering the Hidden Half of Plants Using New Advances in Root Phenotyping. Curr. Opin. Biotechnol. 2019, 55, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rogers, H.H.; Bottomley, P.A. In Situ Nuclear Magnetic Resonance Imaging of Roots: Influence of Soil Type, Ferromagnetic Particle Content, and Soil Water. Agron. J. 1987, 79, 957–965. [Google Scholar] [CrossRef]
- Van Dusschoten, D.; Metzner, R.; Kochs, J.; Postma, J.A.; Pflugfelder, D.; Bühler, J.; Schurr, U.; Jahnke, S. Quantitative 3D Analysis of Plant Roots Growing in Soil Using Magnetic Resonance Imaging1 [OPEN]. Plant Physiol. 2016, 170, 1176–1188. [Google Scholar] [CrossRef] [PubMed]
- Picon-Cochard, C.; Pilon, R.; Tarroux, E.; Pagès, L.; Robertson, J.; Dawson, L. Effect of Species, Root Branching Order and Season on the Root Traits of 13 Perennial Grass Species. Plant Soil 2012, 353, 47–57. [Google Scholar] [CrossRef]
- Prieto, I.; Roumet, C.; Cardinael, R.; Dupraz, C.; Jourdan, C.; Kim, J.H.; Maeght, J.L.; Mao, Z.; Pierret, A.; Portillo, N.; et al. Root Functional Parameters along a Land-Use Gradient: Evidence of a Community-Level Economics Spectrum. J. Ecol. 2015, 103, 361–373. [Google Scholar] [CrossRef]
- Blümich, B.; Perlo, J.; Casanova, F. Mobile Single-Sided NMR. Prog. Nucl. Magn. Reson. Spectrosc. 2008, 52, 197–269. [Google Scholar] [CrossRef]
- Traoré, A.; Aliouissi, R.; Benmoussa, A.; Pagès, G.; Bonny, J.-M. Profiling the Temperature Dependent Frequency of an Open-Magnet for Outdoor Applications. In Proceedings of the 15th International Conference on Magnetic Resonance Microscopy (ICMRM), Paris, France, 18–22 August 2019. [Google Scholar]
- Huck, M.G.; Klepper, B.; Taylor, H.M. Diurnal Variations in Root Diameter. Plant Physiol. 1970, 45, 529–530. [Google Scholar] [CrossRef]
- Blümich, B.; Blümler, P.; Eidmann, G.; Guthausen, A.; Haken, R.; Schmitz, U.; Saito, K.; Zimmer, G. The NMR-Mouse: Construction, Excitation, and Applications. Magn. Reson. Imaging 1998, 16, 479–484. [Google Scholar] [CrossRef]
- Lawson, C.L.; Hanson, R.J. Solving Least Squares Problems; Classics in Applied Mathematics: 15; SIAM: Philadelphia, PA, USA, 1995; ISBN 0-89871-356-0. [Google Scholar]
- Bjarnason, T.A.; McCreary, C.R.; Dunn, J.F.; Mitchell, J.R. Quantitative T2 Analysis: The Effects of Noise, Regularization, and Multivoxel Approaches. Magn. Reson. Med. 2010, 63, 212–217. [Google Scholar]
- Whittall, K.P.; MacKay, A.L. Quantitative Interpretation of NMR Relaxation Data. J. Magn. Reson. 1989, 84, 134–152. [Google Scholar] [CrossRef]
- Hahn, E.L. Spin Echoes. Phys. Rev. 1950, 80, 580–594. [Google Scholar] [CrossRef]
- Carr, H.Y.; Purcell, E.M. Effects of Diffusion on Free Precession in Nuclear Magnetic Resonance Experiments. Phys. Rev. 1954, 94, 630–638. [Google Scholar] [CrossRef]
- Hahn, E.L. Detection of Sea-Water Motion by Nuclear Precession. J. Geophys. Res. 1960, 65, 776–777. [Google Scholar] [CrossRef]
- Packer, K.J.; Rees, C.; Tomlinson, D.J. Studies of Diffusion and Flow by Pulsed NMR Techniques. Adv. Mol. Relax. Process. 1972, 3, 119–131. [Google Scholar] [CrossRef]
- Ryser, P. The Mysterious Root Length. Plant Soil 2006, 286, 1–6. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | D. glomerata | P. lanceolata | M. sativa |
|---|---|---|---|
| Total root length (m) | 56.237 | 17.498 | 46.137 |
| Total root volume (cm3) | 2.431 | 0.780 | 5.647 |
| Total root dry mass (g) | 0.507 | 0.132 | 1.634 |
| Mean root diameter (mm) | 0.223 | 0.270 | 0.432 |
| Mean root water content (g g−1) * | 0.772 | 0.808 | 0.675 |
| Total leaf area (cm2) | 6055.8 | 2166.6 | 4976.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuixe, M.; Traoré, A.S.; Blystone, S.; Bonny, J.-M.; Falcimagne, R.; Pagès, G.; Picon-Cochard, C. Circadian Variation of Root Water Status in Three Herbaceous Species Assessed by Portable NMR. Plants 2021, 10, 782. https://doi.org/10.3390/plants10040782
Nuixe M, Traoré AS, Blystone S, Bonny J-M, Falcimagne R, Pagès G, Picon-Cochard C. Circadian Variation of Root Water Status in Three Herbaceous Species Assessed by Portable NMR. Plants. 2021; 10(4):782. https://doi.org/10.3390/plants10040782
Chicago/Turabian StyleNuixe, Magali, Amidou Sissou Traoré, Shannan Blystone, Jean-Marie Bonny, Robert Falcimagne, Guilhem Pagès, and Catherine Picon-Cochard. 2021. "Circadian Variation of Root Water Status in Three Herbaceous Species Assessed by Portable NMR" Plants 10, no. 4: 782. https://doi.org/10.3390/plants10040782
APA StyleNuixe, M., Traoré, A. S., Blystone, S., Bonny, J.-M., Falcimagne, R., Pagès, G., & Picon-Cochard, C. (2021). Circadian Variation of Root Water Status in Three Herbaceous Species Assessed by Portable NMR. Plants, 10(4), 782. https://doi.org/10.3390/plants10040782