
Optimizing Sampling and Extraction Methods for Plant-Parasitic and Entomopathogenic Nematodes


Abstract
1. Introduction
2. Sampling Goal and Conceived Scenario
3. Ecological Considerations and Concepts
4. Sampling Tools
5. Addressing Nematode Distribution Patterns
6. Indices of Nematode Dispersion
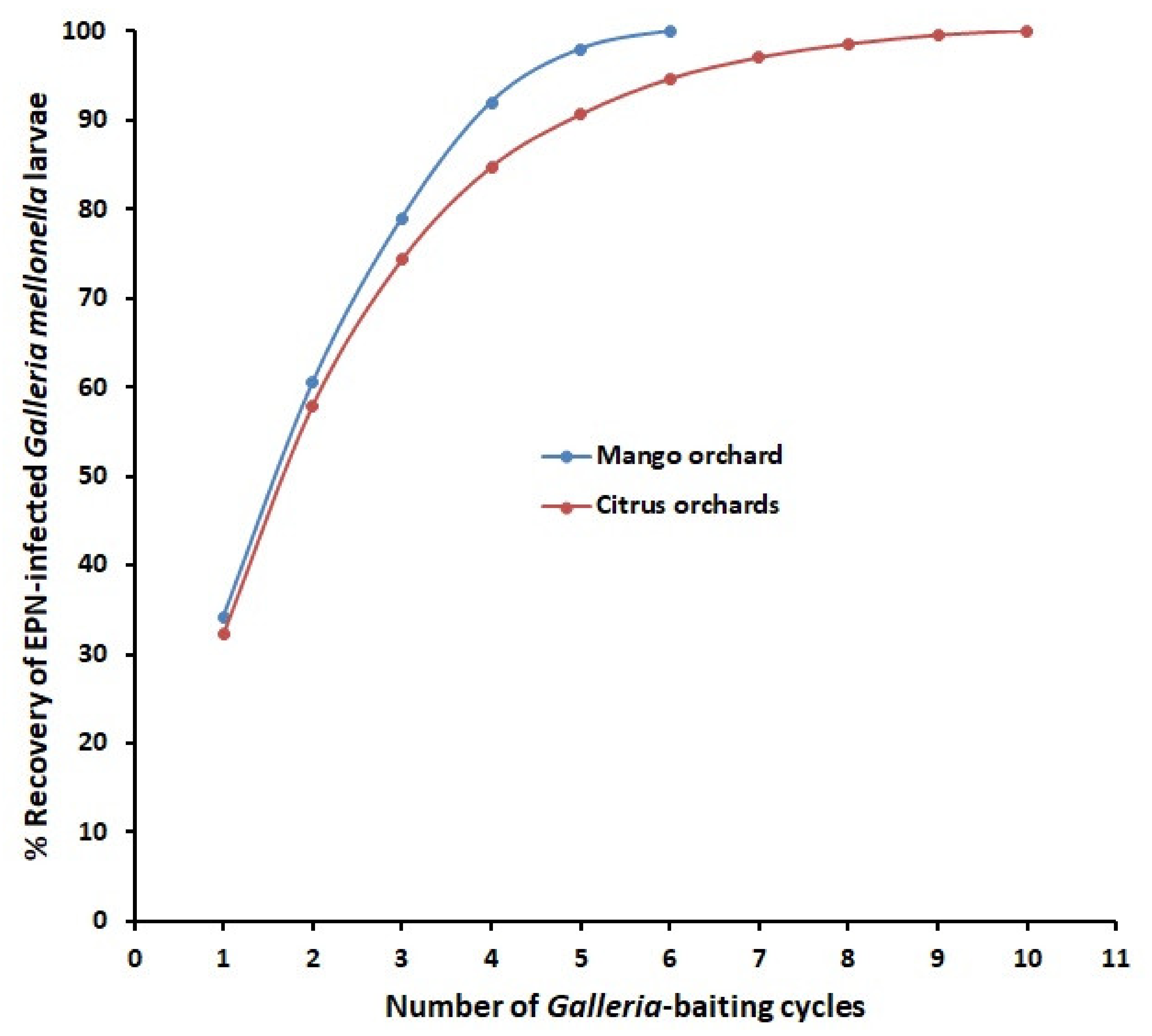
7. Other Examples to Optimize EPN Sampling and Extraction
8. Other Sampling and Extraction Methods
9. Quantifying Extraction Efficiency of EPNs with a Model Used for PPNs
10. Molecular vs. Traditional Sampling and Extraction Technology
Funding
Acknowledgments
Conflicts of Interest
References
- Sikora, D.C.; Hallmann, J.; Timper, P. (Eds.) Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; CABI: Boston, MA, USA, 2018. [Google Scholar]
- Abd-Elgawad, M.M.; Askary, T.H.; Coupland, J. Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes; CAB International: Wallingford, UK, 2017. [Google Scholar]
- Abd-Elgawad, M.M.; Askary, T.H. Impact of phytonematodes on agriculture economy. In Biocontrol Agents of Phytonematodes; Askary, T.H., Martinelli, P.R.P., Eds.; CABI: Wallingford, UK, 2015; pp. 3–49. [Google Scholar]
- Campos-Herrera, R.; Stuart, R.J.; Pathak, E.; El-Borai, F.E.; Duncan, L.W. Temporal patterns of entomopathogenic nematodes in Florida citrus orchards: Evidence of natural regulation by microorganisms and nematode competitors. Soil Biol. Biochem. 2019, 128, 193–204. [Google Scholar] [CrossRef]
- Been, T.H.; Schomaker, C.H. Distribution patterns and sampling. In Plant Nematology, 2nd ed.; Perry, R.N., Moens, M., Eds.; CABI: Wallingford, UK, 2013; pp. 331–358. [Google Scholar]
- Hallmann, J.; Subbotin, S.A. Methods for extraction, processing and detection of plant and soil nematodes. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; Sikora, R.A., Coyne, D., Hallmann, J., Timper, P., Eds.; CABI: Boston, MA, USA, 2018; pp. 87–119. [Google Scholar]
- Duncan, L.W.; Phillips, M.S. Sampling root-knot nematodes. In Root-Knot Nematodes; Perry, R.N., Moens, M., Starr, J.L., Eds.; CABI: St. Albans, UK, 2009; pp. 275–300. [Google Scholar]
- Abd-Elgawad, M.M. Accuracy and Precision of Phytonematode Sampling Plans. Agric. Eng. Int. CIGR J. 2017, 19, 6–15. [Google Scholar]
- Timper, P. Conserving and Enhancing Biological Control of Nematodes. J. Nematol. 2014, 46, 75–89. [Google Scholar]
- Duncan, L.W.; Stuart, R.J.; El-Borai, F.E.; Campos-Herrera, R.; Pathak, E.; Giurcanu, M.; Graham, J.H. Modifying orchard planting sites conserves entomopathogenic nematodes, reduces weevil herbivory and increases citrus tree growth, survival and fruit yield. Biol. Control 2013, 64, 26–36. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Spence, K.O.; Nakatani, J.; Lewis, E.E. Effect of soil salinity on entomopathogenic nematode survival and behaviour. Nematology 2011, 3, 859–867. [Google Scholar] [CrossRef]
- Hussaini, S.S. Entomopathogenic nematodes: Ecology, diversity and geographical distribution. In Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes; Abd-Elgawad, M.M.M., Askary, T.H., Coupland, J., Eds.; CABI: Wallingford, UK, 2017; pp. 88–142. [Google Scholar]
- Campos-Herrera, R.; Pathak, E.; El-Borai, F.E.; Schumann, A.; Abd-Elgawad, M.M.; Duncan, L.W. New citriculture system suppresses native and augmented entomopathogenic nematodes. Biol. Control 2013, 66, 183–194. [Google Scholar] [CrossRef]
- Van Bezooijen, J. Methods and Techniques for Nematology (A Manual); Wageningen University: Wageningen, The Netherlands, 2006. [Google Scholar]
- Dritsoulas, A.; Duncan, L.W. Optimizing for taxonomic coverage: A comparison of methods to recover mesofauna from soil. J. Nematol. 2020, 52, 1–9. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Lacey, L.A. Methods for studying the ecology of invertebrate diseases and pathogens. In Ecology of Invertebrate Diseases; Hajek, A.E., Shapiro-Ilan, D.I., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2018; pp. 19–48. [Google Scholar]
- Berg, W.V.D.; Hartsema, O.; Nijs, L.J.D. Statistical analysis of nematode counts from interlaboratory proficiency tests. Nematology 2014, 16, 229–243. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M. Towards optimization of entomopathogenic nematodes for more service in the biological control of insect pests. Egypt. J. Biol. Pest Control 2019, 29, 77. [Google Scholar] [CrossRef]
- Elliott, J.M. Some Methods for the Statistical Analysis of Samples of Benthic Invertebrates; Freshwater Biological Association, Science Publication No. 15; The Ferry House: Ambleside, UK, 1971. [Google Scholar]
- Wilson, M.J.; Lewis, E.E.; Yoder, F.; Gaugler, R. Application pattern and persistence of the entomopathogenic nematode Heterorhabditis bacteriophora. Biol. Control 2003, 26, 180–188. [Google Scholar] [CrossRef]
- Spiridonov, S.E.; Moens, M.; Wilson, M.J. Fine scale spatial distributions of two entomopathogenic nematodes in a grassland soil. Appl. Soil Ecol. 2007, 37, 192–201. [Google Scholar] [CrossRef]
- Bal, H.K.; Acosta, N.; Cheng, Z.; Grewal, P.S.; Hoy, C.W. Effect of habitat and soil management on dispersal and distribution patterns of entomopathogenic nematodes. Appl. Soil Ecol. 2017, 121, 48–59. [Google Scholar] [CrossRef]
- Wu, S.-Y.; El-Borai, F.E.; Graham, J.H.; Duncan, L.W. Geospatial relationships between native entomopathogenic nematodes and Fusarium solani in a Florida citrus orchard. Appl. Soil Ecol. 2019, 140, 108–114. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M. Can rational sampling maximise isolation and fix distribution measure of entomopathogenic nematodes? Nematology 2020, 22, 907–916. [Google Scholar] [CrossRef]
- Stuart, R.; Gaugler, R. Patchiness in Populations of Entomopathogenic Nematodes. J. Invertebr. Pathol. 1994, 64, 39–45. [Google Scholar] [CrossRef]
- Duncan, L.W.; Ferguson, J.J.; Dunn, R.A.; Noling, J.W. Application of Taylor’s Power Law to Sample Statistics of Tylenchulus semipenetrans in Florida Citrus. J. Nematol. 1989, 21, 707–711. [Google Scholar] [PubMed]
- Abd-Elgawad, M. Spatial distribution of the phytonematode community in Egyptian citrus groves. Fundam. Appl. Nematol. 1992, 14, 367–373. [Google Scholar]
- Abd-Elgawad, M.M. Spatial patterns of Tuta absoluta and heterorhabditid nematodes. Russ. J. Nematol. 2014, 22, 89–100. [Google Scholar]
- Abd-Elgawad, M.M. Managing nematodes in Egyptian citrus orchards. Bull. Natl. Res. Centre 2020, 44, 41. [Google Scholar] [CrossRef]
- Ferris, H.; Goodell, P.B.; McKenry, M.V. Sampling for nematodes. Calif. Agric. 1981, 35, 13–15. [Google Scholar]
- Taylor, L.R. Aggregation, Variance and the Mean. Nat. Cell Biol. 1961, 189, 732–735. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Ali, J.G.; Diaz, B.M.; Duncan, L.W. Analyzing spatial patterns linked to the ecology of herbivores and their natural enemies in the soil. Front. Plant Sci. 2013, 4, 378. [Google Scholar] [CrossRef]
- Perry, J.N. Spatial Analysis by Distance Indices. J. Anim. Ecol. 1995, 64, 303. [Google Scholar] [CrossRef]
- Gorny, A.M.; Hay, F.S.; Esker, P.; Pethybridge, S.J. Spatial and spatiotemporal analysis of Meloidogyne hapla and Pratylenchus penetrans populations in commercial potato fields in New York, USA. Nematology 2020, 23, 139–151. [Google Scholar] [CrossRef]
- Stuart, R.J.; Barbercheck, M.E.; Grewal, P.S. Entomopathogenic nematodes in the soil environment: Distributions, interactions and the influence of biotic and abiotic factors. In Nematode Pathogenesis of Insects and Other Pests; Campos-Herrera, R., Ed.; Springer: Cham, Switzerland, 2015; pp. 97–138. [Google Scholar]
- Winder, L.; Alexander, C.; Griffiths, G.; Holland, J.; Wooley, C.; Perry, J. Twenty years and counting with SADIE: Spatial Analysis by Distance Indices software and review of its adoption and use. Rethink. Ecol. 2019, 4, 1–16. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; El-Borai, F.E.; Duncan, L.W. Wide interguild relationships among entomopathogenic and free-living nematodes in soil as measured by real time qPCR. J. Invertebr. Pathol. 2012, 111, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, C.; Ricci, B.; Franck, P.; Senoussi, R. Spatial analyses of ecological count data: A density map comparison approach. Basic Appl. Ecol. 2010, 11, 734–742. [Google Scholar] [CrossRef]
- Li, B.; Madden, L.V.; Xu, X. Spatial analysis by distance indices: An alternative local clustering index for studying spatial patterns. Methods Ecol. Evol. 2012, 3, 368–377. [Google Scholar] [CrossRef]
- Stock, S.P. Diversity, biology and evolutionary relationships. In Nematode Pathogenesis of Insects and Other Pests; Campos-Herrera, R., Ed.; Springer: Cham, Switzerland, 2015; pp. 3–27. [Google Scholar]
- Abdel-Razek, S.A.; Hussein, M.; Shehata, I. Isolation and identification of indigenous entomopathogenic nematode (EPN) isolate from Egyptian fauna. Arch. Phytopathol. Plant Prot. 2018, 51, 197–206. [Google Scholar] [CrossRef]
- Shehata, I.E.; Hammam, M.M.A.; El-Borai, F.E.; Duncan, L.W.; Abd-Elgawad, M.M.M. Comparison of virulence, reproductive potential, and persistence among local Heterorhabditis indica populations for the control of Temnorhynchus baal (Reiche & Saulcy) (Coleoptera: Scarabaeidae) in Egypt. Egypt. J. Biol. Pest Control. 2019, 29, 32. [Google Scholar] [CrossRef]
- Nguyen, K.B.; Smart, G.C., Jr. Steinernema scapterisci n. sp. (Steinernematidae: Nematoda). J. Nematol. 1990, 22, 187–199. [Google Scholar] [PubMed]
- Gaugler, R.; Han, R. Production technology. In Entomopathogenic Nematology; Gaugler, R., Ed.; CABI: Wallingford, UK, 2002; pp. 289–310. [Google Scholar]
- Fan, X.; Hominick, W.M. Efficiency of the Galleria (wax moth) baiting technique for recovering infective stages of entomopathogenic rhabditids (Steinernematidae and Heterorhabditidae) from sand and soil. Rev. Nematol. 1991, 14, 381–387. [Google Scholar]
- Griffin, C.T. Behaviour and population dynamics of entomopathogenic nematodes following application. In Nematode Pathogenesis of Insects and Other Pests; Campos-Herrera, R., Ed.; Springer: Cham, Switzerland, 2015; pp. 139–163. [Google Scholar]
- Lewis, E.E.; Hazir, S.; Hodson, A.; Gulcu, B. Trophic relationships of entomopathogenic nematodes in agricultural habitats. In Nematode Pathogenesis of Insects and Other Pests; Campos-Herrera, R., Ed.; Springer: Cham, Switzerland, 2015; pp. 139–163. [Google Scholar]
- White, G.F. A method for obtaining infective nematode larvae from cultures. Science 1927, 66, 302–303. [Google Scholar] [CrossRef]
- Duncan, L.W.; Graham, J.H.; Dunn, D.C.; Zellers, J.; McCoy, C.W.; Nguyen, K. Incidence of endemic entomopathogenic nematodes following application of Steinernema riobrave for control of Diaprepes abbreviatus. J. Nematol. 2003, 35, 178–186. [Google Scholar]
- Shapiro-Ilan, D.I.; Gardner, W.A.; Fuxa, J.R.; Wood, B.W.; Nguyen, K.B.; Adams, B.J.; Humber, R.A.; Hall, M.J. Survey of Entomopathogenic Nematodes and Fungi Endemic to Pecan Orchards of the Southeastern United States and Their Virulence to the Pecan Weevil (Coleoptera: Curculionidae). Environ. Entomol. 2003, 32, 187–195. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Gutierrez, C. Comparative study of entomopathogenic nematode isolation usingGalleria mellonella(Pyralidae) andSpodoptera littoralis(Noctuidae) as baits. Biocontrol Sci. Technol. 2008, 18, 621–626. [Google Scholar] [CrossRef]
- Woodring, J.L.; Kaya, H.K. Steinernematid and Heterorhabditid Nematodes: A Handbook of Techniques; Southern Cooperative Series Bulletin No. 331; Arkansas Agricultural Experiment Station: Fayetteville, AR, USA, 1988. [Google Scholar]
- Seinhorst, J.W. Extraction methods for nematodes inhabiting soil. In Progress in Soil Zoology; Murphy, P.W., Ed.; Butterworths: London, UK, 1962; pp. 243–256. [Google Scholar]
- Ferris, H. Extraction efficiencies and population estimation. In Vistas on Nematology; Veech, J.A., Dickson, D.W., Eds.; Society of Nematologists: Hyattsville, MD, USA, 1987; pp. 59–63. [Google Scholar]
- Adams, J.B.; Nguyen, K.B. Taxonomy and systematics. In Entomopathogenic Nematology; Gaugler, R., Ed.; CABI: Wallingford, UK, 2002; pp. 1–33. [Google Scholar]
- Molinari, S.; Leonetti, P. Bio-control agents activate plant immune response and prime susceptible tomato against root-knot nematodes. PLoS ONE 2019, 14, e0213230. [Google Scholar] [CrossRef]
- Carneiro, R.M.D.G.; Silva, F.S.O.; Correia, V.R. Methods and tools currently used for the identification of plant parasitic nematodes. In Nematology-Concepts, Diagnosis and Control; Shah, M.M., Mahamood, M., Eds.; InTech Open: Rijeka, Croatia, 2017; pp. 19–35. [Google Scholar] [CrossRef]
- Dritsoulas, A.; Campos-Herrera, R.; Blanco-Pérez, R.; Duncan, L.W. Comparing high throughput sequencing and real time qPCR for characterizing entomopathogenic nematode biogeography. Soil Biol. Biochem. 2020, 145, 107793. [Google Scholar] [CrossRef]
- Torr, P.; Spiridonov, S.E.; Heritage, S.; Wilson, M.J. Habitat associations of two entomopathogenic nematodes: A quantitative study using real-time quantitative polymerase chain reactions. J. Anim. Ecol. 2007, 76, 238–245. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Johnson, E.; El-Borai, F.; Stuart, R.; Graham, J.; Duncan, L. Long-term stability of entomopathogenic nematode spatial patterns in soil as measured by sentinel insects and real-time PCR assays. Ann. Appl. Biol. 2011, 158, 55–68. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Martín, J.A.; Escuer, M.; García-González, M.T.; Duncan, L.W.; Gutiérrez, C. Entomopathogenic nematode food webs in an ancient, mining pollution gradient in Spain. Sci. Total Environ. 2016, 572, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Porazinska, D.L.; Giblin-Davis, R.M.; Faller, L.; Farmerie, W.; Kanzaki, N.; Morris, K.; Powers, T.O.; Tucker, A.E.; Sung, W.; Thomas, W.K. Evaluating high-throughput sequencing as a method for metagenomic analysis of nematode diversity. Mol. Ecol. Resour. 2009, 9, 1439–1450. [Google Scholar] [CrossRef]
- Jaffuel, G.; Mader, P.; Blanco-Perez, R.; Chiriboga, X.; Fliessbach, A.; Turlings, T.C.J.; Campos-Herrera, R. Prevalence and activity of entomopathogenic nematodes and their antagonists in soils that are subject to different agricultural practices. Agric. Ecosyst. Environ. 2016, 230, 329–340. [Google Scholar] [CrossRef]
- Regaieg, H.; Ciancio, A.; Raouani, N.H.; Rosso, L. Detection and biocontrol potential of Verticillium leptobactrum parasitizing Meloidogyne spp. World J. Microbiol. Biotechnol. 2010, 27, 1615–1623. [Google Scholar] [CrossRef]
- Ciancio, A.; Loffredo, A.; Paradies, F.; Turturo, C.; Sialer, M.F. Detection of Meloidogyne incognita and Pochonia chlamydosporia by fluorogenic molecular probes. EPPO Bull. 2005, 35, 157–164. [Google Scholar] [CrossRef]
- Stirling, G.R. Biological Control of Plant-Parasitic Nematodes, 2nd ed.; CABI: Wallingford, UK, 2014. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| EPN Studied Population | Form of the Measured EPN | Ia Value | Comments (Location) | Reference |
|---|---|---|---|---|
| Heterorhabditis bacteriophora-infective juveniles (IJ) applied uniformly, in one, or nine patches on Kentucky bluegrass | EPN-infected Galleria mellonella larvae over time | All mean values were less than one but differed (p ≤ 0.05) until 20 weeks, no more, after EPN application | The values suggest a more even distribution than a random one (NJ/USA) | [20] |
| Natural populations of Steinernema feltiae and S. affine in grassland plots | IJ assigned to one of 4 groups of increasing physiological age | The values ranged 1.27–1.45 | All values indicate aggregated distribution (Merelbeke/Belgium) | [21] |
| H. bacteriophora or S. carpocapsae-infected G. mellonella larvae applied within 24 h of initial IJ emergence to cultivated fields and adjoining grassy border plots | H. bacteriophora and S. carpocapsae-IJ recovered from G. mellonella larvae baits applied several times after the cadavers | Range <1 to >2. Mean values differed between EPN species in bait traps and between soil management regimes at 48 h and 16 days after placing the cadavers, respectively | Spatial distributions dispersed from a grassy border to the adjacent cultivated field plots were more aggregated for H. bacteriophora than for S. carpocapsae (OH/USA) | [22] |
| Steinernema diaprepesi, Heterorhabditis indica, and Heterorhabditis zealandica | EPN were measured using quantitative qPCR during a 6-month citrus orchard survey | The values ranged 0.8–1.3 over 6 months and could be compared with those of the fungus and Diaprepes root weevil | Highly significant spatial associations between Fusarium solani and EPN communities of up to three EPN species (FL, USA) | [23] |
| Natural populations of H. indica in citrus groves | EPN-infected G. mellonella larvae | 0.913 | Ia refers to uniform distribution pattern (Giza, Egypt) | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-Elgawad, M.M.M. Optimizing Sampling and Extraction Methods for Plant-Parasitic and Entomopathogenic Nematodes. Plants 2021, 10, 629. https://doi.org/10.3390/plants10040629
Abd-Elgawad MMM. Optimizing Sampling and Extraction Methods for Plant-Parasitic and Entomopathogenic Nematodes. Plants. 2021; 10(4):629. https://doi.org/10.3390/plants10040629
Chicago/Turabian StyleAbd-Elgawad, Mahfouz M. M. 2021. "Optimizing Sampling and Extraction Methods for Plant-Parasitic and Entomopathogenic Nematodes" Plants 10, no. 4: 629. https://doi.org/10.3390/plants10040629
APA StyleAbd-Elgawad, M. M. M. (2021). Optimizing Sampling and Extraction Methods for Plant-Parasitic and Entomopathogenic Nematodes. Plants, 10(4), 629. https://doi.org/10.3390/plants10040629