
Root and Shoot Response to Nickel in Hyperaccumulator and Non-Hyperaccumulator Species





Abstract
1. Introduction
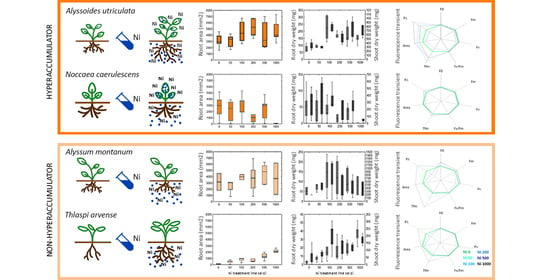
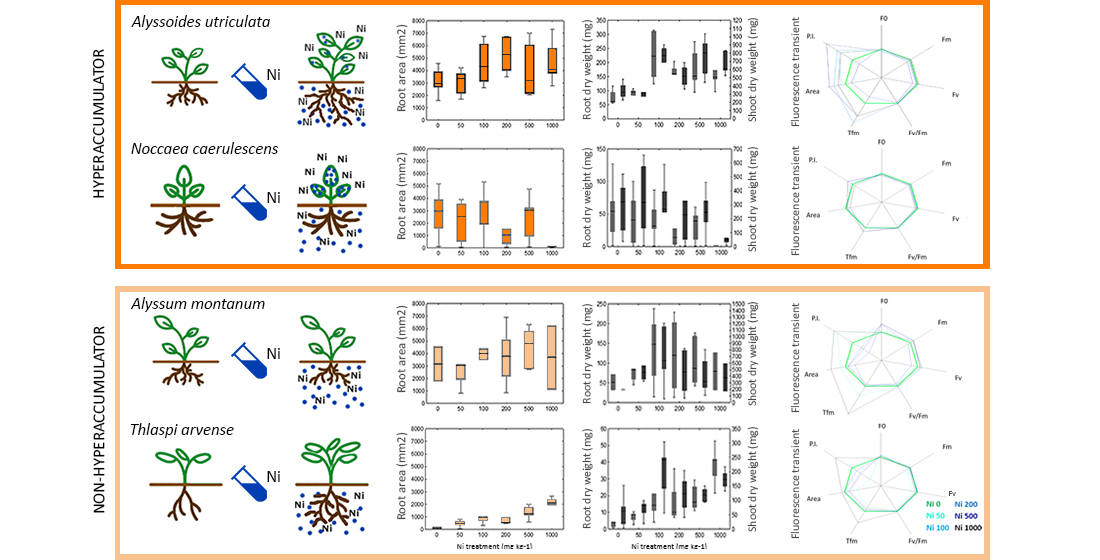
2. Results
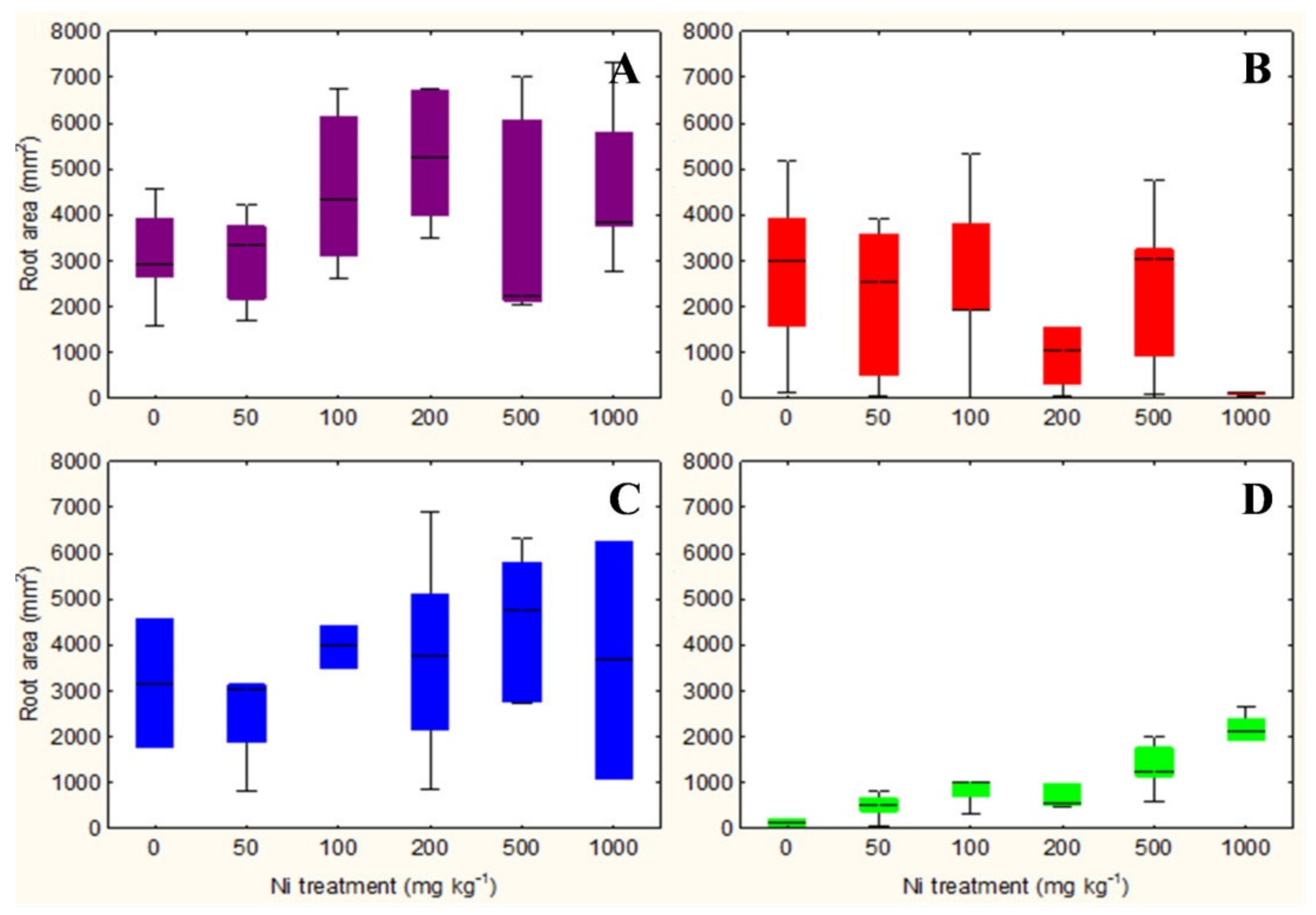
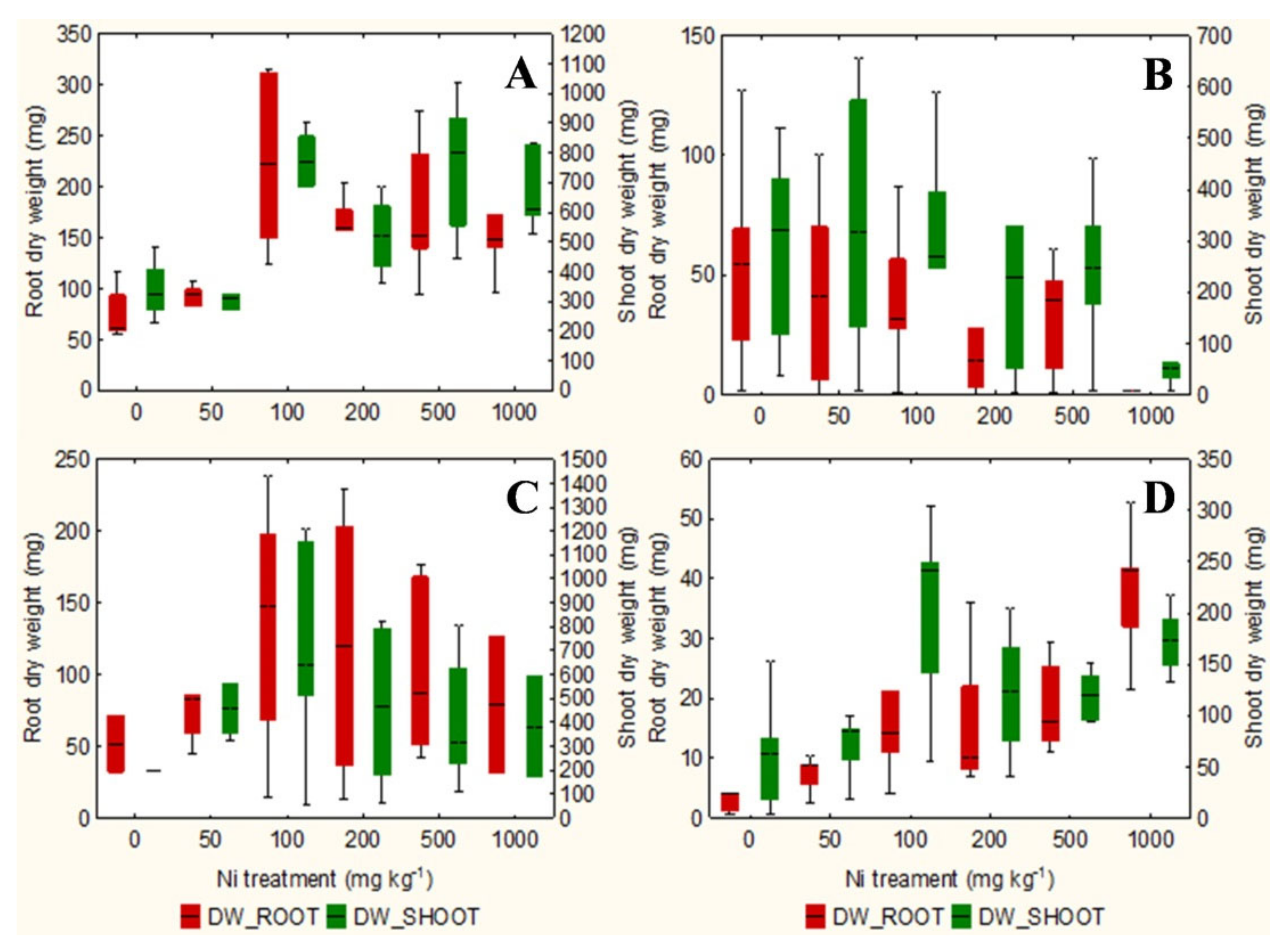
2.1. Evaluation of Root Area, Biomass, and Plant Water Content
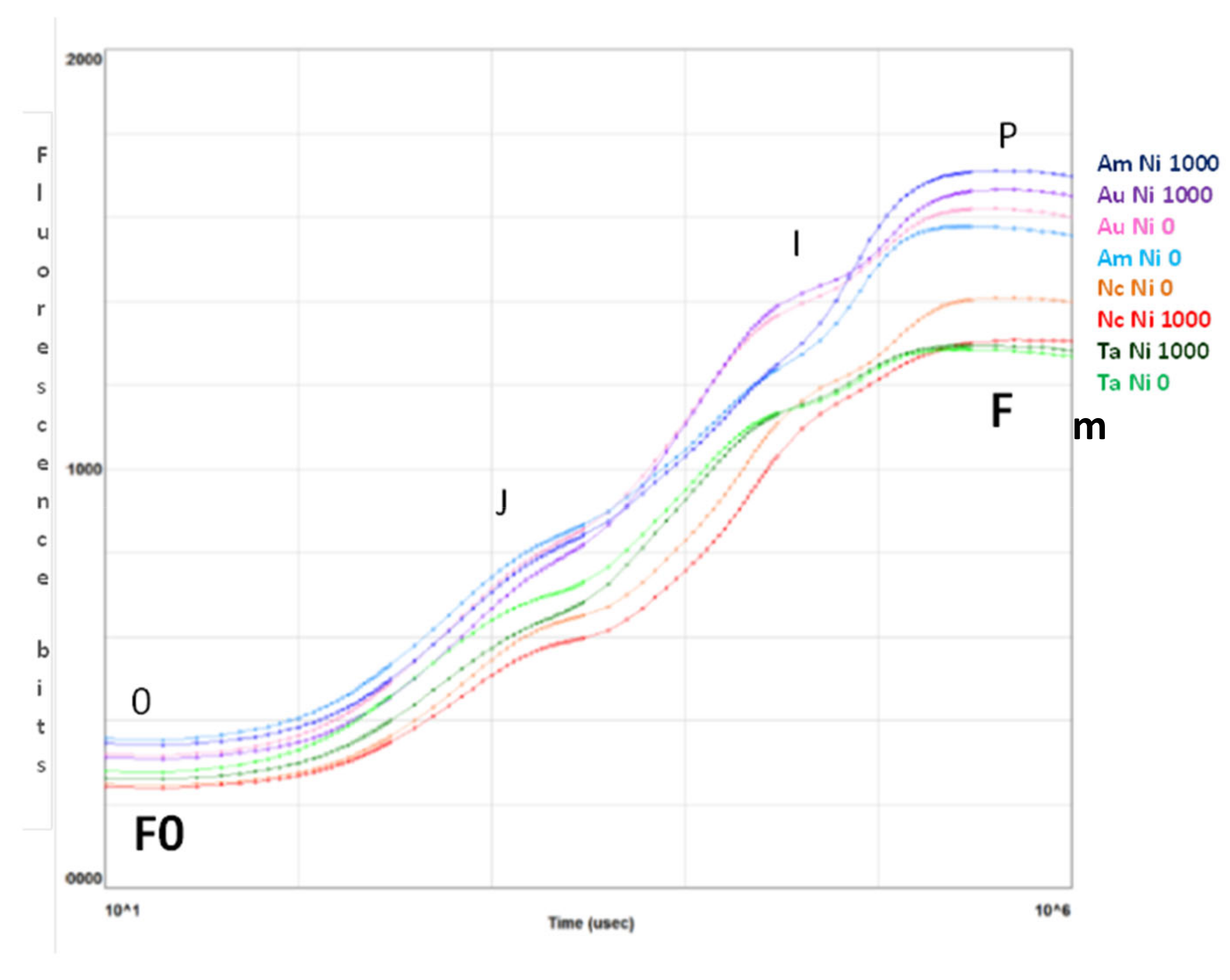
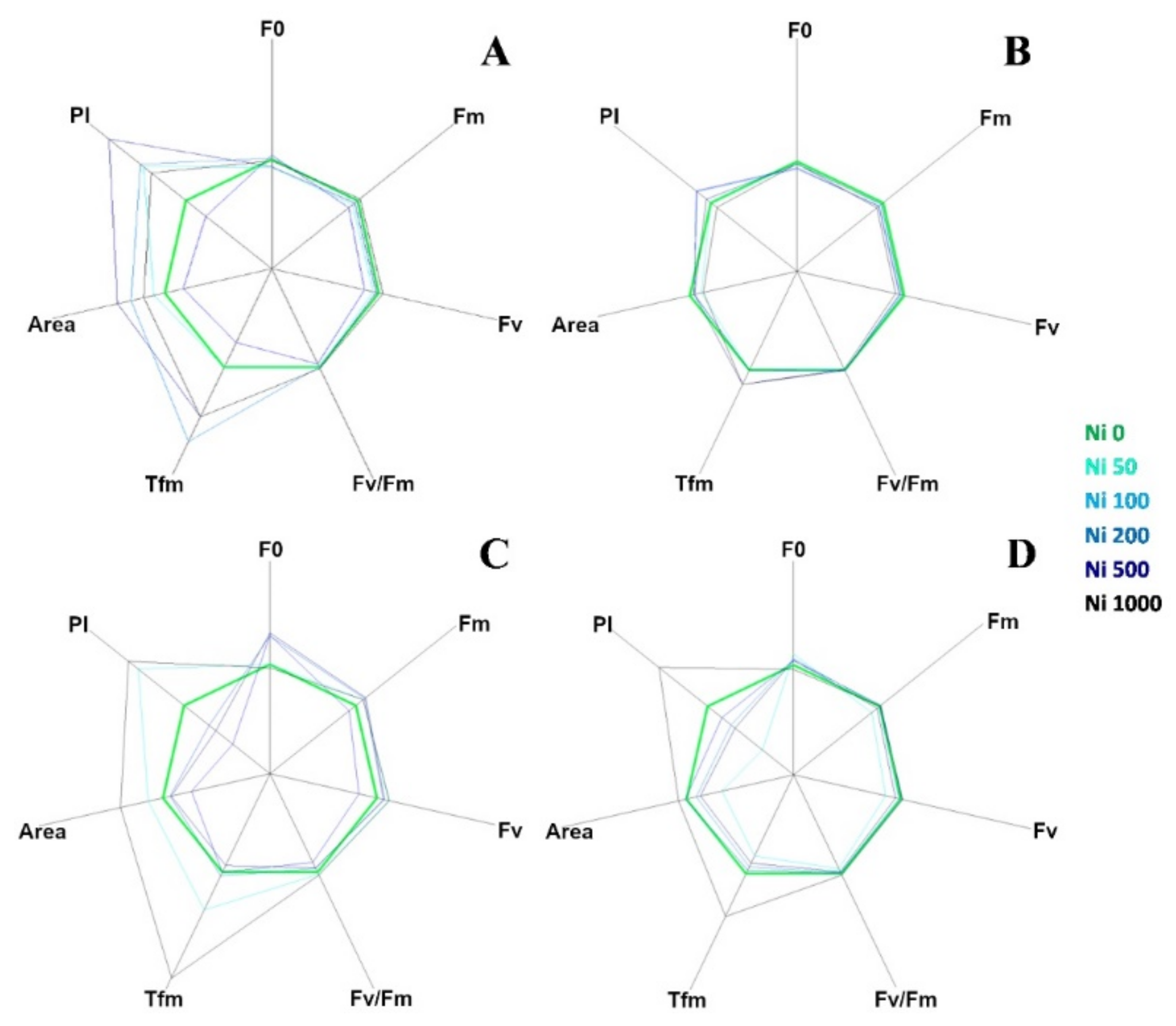
2.2. Photosynthetic Efficiency
3. Discussion
3.1. Evaluation of Root Area, Biomass, and Plant Water Content
3.2. Photosynthetic Efficiency
4. Materials and Methods
4.1. Plant Species
4.2. Seed Collection
4.3. Pot Experiment
4.3.1. Evaluation of Root Area, Biomass, and Plant Water Content
4.3.2. Photosynthethic Efficiency
- F0 is the minimum fluorescence value and represents emission by the excited chlorophyll a molecule in the antenna structure of Photosystem II.
- Fm is the maximum fluorescence value obtained after the application of a saturation pulse to the dark-adapted leaf.
- Fv is the variable fluorescence, and it denotes the variable component of the recording and relates to the maximum capacity for photochemical quenching.
- Fv/Fm is widely used to indicate the maximum quantum efficiency of Photosystem II. It is a sensitive indication of plant photosynthetic performance.
- Tfm is used to express the time at which the maximum fluorescence value (Fm) was reached.
- Area above the fluorescence curve between F0 and Fm is proportional to the pool size of electron acceptors.
- P.I. (Performance Index) is essentially an indicator of sample vitality.
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Consulted Websites
- Acta Plantarum-Flora delle Regioni italiane https://www.actaplantarum.org./ (accessed 11 December 2020).
- ENSCOBASE: the ENSCONET Virtual Seed Bank http://enscobase.maich.gr/ (accessed 8 October 2018).
- Global Hyperaccumulator Database http://hyperaccumulators.smi.uq.edu.au/collection/ (accessed 1 February 2021).
- KEW Royal Botanic Gardens http://data.kew.org/sid/ (accessed 9 November 2020).
References
- Wenzel, W.W.; Lombi, E.; Adriano, D.C. Root and Rhizosphere Processes in Metal Hyperaccumulation and Phytoremediation Technology. In Heavy Metal Stress in Plants; Springer: Berlin/Heidelberg, Germany, 2004; pp. 313–344. [Google Scholar] [CrossRef]
- Alford, É.R.; Pilon-Smits, E.A.H.; Paschke, M.W. Metallophytes—A View from the Rhizosphere. Plant Soil 2010, 337, 33–50. [Google Scholar] [CrossRef]
- Hodge, A. The Plastic Plant: Root Responses to Heterogeneous Supplies of Nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Dechamps, C.; Noret, N.; Mozek, R.; Draye, X.; Meerts, P. Root Allocation in Metal-Rich Patch by Thlaspi caerulescens from Normal and Metalliferous Soil—New Insights into the Rhizobox Approach. Plant Soil 2008, 310, 211. [Google Scholar] [CrossRef]
- Himmelbauer, M.L.; Puschenreiter, M.; Schnepf, A.; Loiskandl, W.; Wenzel, W.W. Root Morphology of Thlaspi goesingense Hálácsy Grown on a Serpentine Soil. J. Plant Nutr. Soil Sci. 2005, 168, 138–144. [Google Scholar] [CrossRef]
- Liu, F.; Tang, Y.; Du, R.; Yang, H.; Wu, Q.; Qiu, R. Root Foraging for Zinc and Cadmium Requirement in the Zn/Cd Hyperaccumulator Plant Sedum alfredii. Plant Soil 2010, 327, 365–375. [Google Scholar] [CrossRef]
- Mincey, K. Ecological and Genetic Investigations of the Nickel Hyperaccumulator Streptanthus polygaloides (Brassicaceae). Ph.D. Dissertation, Auburn University, Auburn, AL, USA, 2018. [Google Scholar]
- Moradi, A.B.; Conesa, H.M.; Robinson, B.H.; Lehmann, E.; Kaestner, A.; Schulin, R. Root Responses to Soil Ni Heterogeneity in a Hyperaccumulator and a Non-Accumulator Species. Environ. Pollut. 2009, 157, 2189–2196. [Google Scholar] [CrossRef] [PubMed]
- Golestanifard, A.; Puschenreiter, M.; Aryan, A.; Santner, J.; Wenzel, W.W. Metal Accumulation and Rhizosphere Characteristics of Noccaea rotundifolia ssp. cepaeifolia. Environ. Pollut. 2020, 266, 115088. [Google Scholar] [CrossRef]
- Keller, C.; Hammer, D.; Kayser, A.; Richner, W.; Brodbeck, M.; Sennhauser, M. Root Development and Heavy Metal Phytoextraction Efficiency: Comparison of Different Plant Species in the Field. Plant Soil 2003, 249, 67–81. [Google Scholar] [CrossRef]
- Kutschera, L.; Lichtenegger, E.; Sobotik, M. Wurzelatlas Mitteleuropäischer Grünlandpflanzen. G. Fischer-Verlag: Stuttgart, Germany, 1992; Volume 1. [Google Scholar]
- Gendre, D.; Czernic, P.; Conéjéro, G.; Pianelli, K.; Briat, J.-F.; Lebrun, M.; Mari, S. TcYSL3, a Member of the YSL Gene Family from the Hyper-Accumulator Thlaspi caerulescens, Encodes a Nicotianamine-Ni/Fe Transporter. Plant J. 2007, 49, 1–15. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy Metal Hyperaccumulating Plants: How and Why Do They Do It? And What Makes Them so Interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of Metal and Metalloid Trace Elements: Facts and Fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Freitas, H.; Prasad, M.N.V.; Pratas, J. Analysis of Serpentinophytes from North–East of Portugal for Trace Metal Accumulation—Relevance to the Management of Mine Environment. Chemosphere 2004, 54, 1625–1642. [Google Scholar] [CrossRef] [PubMed]
- Ghaderian, S.M.; Mohtadi, A.; Rahiminejad, R.; Reeves, R.D.; Baker, A.J.M. Hyperaccumulation of Nickel by Two Alyssum Species from the Serpentine Soils of Iran. Plant Soil 2007, 293, 91–97. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J. Metal Accumulating Plants. Phytoremediation Toxic Met. Using Plants Clean Up environment; Ensley, R.I., Ed.; John Wiley Sons Inc.: New York, NY, USA, 2000. [Google Scholar]
- Roccotiello, E.; Serrano, H.C.; Mariotti, M.G.; Branquinho, C. Nickel Phytoremediation Potential of the Mediterranean Alyssoides utriculata (L.) Medik. Chemosphere 2015, 119, 1372–1378. [Google Scholar] [CrossRef] [PubMed]
- Turgay, O.C.; Görmez, A.; Bilen, S. Isolation and Characterization of Metal Resistant-Tolerant Rhizosphere Bacteria from the Serpentine Soils in Turkey. Environ. Monit. Assess. 2012, 184, 515–526. [Google Scholar] [CrossRef]
- Sobczyk, M.K.; Smith, J.A.C.; Pollard, A.J.; Filatov, D.A. Evolution of Nickel Hyperaccumulation and Serpentine Adaptation in the Alyssum serpyllifolium Species Complex. Heredity 2017, 118, 31–41. [Google Scholar] [CrossRef]
- Rajakaruna, N.; Harris, T.B.; Alexander, E.B. Serpentine Geoecology of Eastern North America: A Review. Rhodora 2009, 111, 21–108. [Google Scholar] [CrossRef]
- Brady, K.U.; Kruckeberg, A.R.; Jr, H.D.B. Evolutionary Ecology of Plant Adaptation to Serpentine Soils. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 243–266. [Google Scholar] [CrossRef]
- Van der Ent, A.; Rajakaruna, N.; Boyd, R.; Echevarria, G.; Repin, R.; Williams, D. Global Research on Ultramafic (Serpentine) Ecosystems (8th International Conference on Serpentine Ecology in Sabah, Malaysia): A Summary and Synthesis. Aust. J. Bot. 2015, 63, 1–16. [Google Scholar] [CrossRef]
- Mengoni, A.; Schat, H.; Vangronsveld, J. Plants as Extreme Environments? Ni-Resistant Bacteria and Ni-Hyperaccumulators of Serpentine Flora. Plant Soil 2010, 331, 5–16. [Google Scholar] [CrossRef]
- Pollard, A.J.; Reeves, R.D.; Baker, A.J.M. Facultative Hyperaccumulation of Heavy Metals and Metalloids. Plant Sci. 2014, 217-218, 8–17. [Google Scholar] [CrossRef]
- Marsili, S.; Roccotiello, E.; Carbone, C.; Marescotti, P.; Cornara, L.; Mariotti, M.G. Plant Colonization on a Contaminated Serpentine Site. Northeast. Nat. 2009, 16, 297–308. [Google Scholar] [CrossRef]
- Global Hyperaccumulator Database. Available online: http://hyperaccumulators.smi.uq.edu.au/collection/ (accessed on 29 January 2021).
- Boyd, R.S. Plant Defense Using Toxic Inorganic Ions: Conceptual Models of the Defensive Enhancement and Joint Effects Hypotheses. Plant Sci. 2012, 195, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Selvi, F.; Carrari, E.; Colzi, I.; Coppi, A.; Gonnelli, C. Responses of Serpentine Plants to Pine Invasion: Vegetation Diversity and Nickel Accumulation in Species with Contrasting Adaptive Strategies. Sci. Total Environ. 2017, 595, 72–80. [Google Scholar] [CrossRef]
- Baker, A.J.M.; Brooks, R. Terrestrial Higher Plants Which Hyperaccumulate Metallic Elements. A Review of Their Distribution, Ecology and Phytochemistry. Biorecovery 1989, 1, 81–126. [Google Scholar]
- Brooks, R.R. Serpentine and Its Vegetation: A Multidisciplinary Approach. Ecol. Phytogeogr. Physiol. Ser. 1987, 1, 52. [Google Scholar] [CrossRef]
- Chiarucci, A.; Baker, A.J.M. Advances in the Ecology of Serpentine Soils. Plant Soil 2007, 293, 1–2. [Google Scholar] [CrossRef]
- Pasquet, C.; Monna, F.; van Oort, F.; Gunkel-Grillon, P.; Laporte-Magoni, C.; Losno, R.; Chateau, C. Mobility of Ni, Co, and Mn in Ultramafic Mining Soils of New Caledonia, Assessed by Kinetic EDTA Extractions. Environ. Monit. Assess. 2018, 190, 638. [Google Scholar] [CrossRef]
- Raous, S.; Becquer, T.; Garnier, J.; de Martins, É.S.; Echevarria, G.; Sterckeman, T. Mobility of Metals in Nickel Mine Spoil Materials. Appl. Geochem. 2010, 25, 1746–1755. [Google Scholar] [CrossRef]
- Roccotiello, E.; Marescotti, P.; Di Piazza, S.; Cecchi, G.; Mariotti, M.G.; Zotti, M. Biodiversity in Metal-Contaminated Sites–Problem and Perspective—A Case Study. Significance 2015, 16, 26–28. [Google Scholar] [CrossRef]
- Sitko, K.; Rusinowski, S.; Kalaji, H.M.; Szopiński, M.; Małkowski, E. Photosynthetic Efficiency as Bioindicator of Environmental Pressure in A. halleri. Plant Physiol. 2017, 175, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, L.; Huang, X.; Zhou, Y.; Quan, Q.; Li, Y.; Zhu, X. Response of Photosynthesis to Different Concentrations of Heavy Metals in Davidia involucrata. PLoS ONE 2020, 15, e0228563. [Google Scholar] [CrossRef]
- Houri, T.; Khairallah, Y.; Zahab, A.A.; Osta, B.; Romanos, D.; Haddad, G. Heavy Metals Accumulation Effects on The Photosynthetic Performance of Geophytes in Mediterranean Reserve. J. King Saud Univ. Sci. 2020, 32, 874–880. [Google Scholar] [CrossRef]
- Chandra, R.; Kang, H. Mixed Heavy Metal Stress on Photosynthesis, Transpiration Rate, and Chlorophyll Content in Poplar Hybrids. For. Sci. Technol. 2016, 12, 55–61. [Google Scholar] [CrossRef]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of Different Metals on Photosynthesis: Cadmium and Zinc Affect Chlorophyll Fluorescence in Durum Wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [PubMed]
- Dobrikova, A.G.; Apostolova, E.L.; Hanć, A.; Yotsova, E.; Borisova, P.; Sperdouli, I.; Adamakis, I.-D.S.; Moustakas, M. Cadmium Toxicity in Salvia sclarea L.: An Integrative Response of Element Uptake, Oxidative Stress Markers, Leaf Structure and Photosynthesis. Ecotoxicol. Environ. Saf. 2021, 209, 111851. [Google Scholar] [CrossRef]
- Khator, K.; Saxena, I.; Shekhawat, G.S. Nitric Oxide Induced Cd Tolerance and Phytoremediation Potential of B. juncea by the Modulation of Antioxidant Defense System and ROS Detoxification. Biometals 2020. [Google Scholar] [CrossRef] [PubMed]
- Rosatto, S.; Roccotiello, E.; Di Piazza, S.; Cecchi, G.; Greco, G.; Zotti, M.; Vezzulli, L.; Mariotti, M. Rhizosphere Response to Nickel in a Facultative Hyperaccumulator. Chemosphere 2019. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence; Springer: Berlin/Heidelberg, Germany, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Mari, S.; Gendre, D.; Pianelli, K.; Ouerdane, L.; Lobinski, R.; Briat, J.-F.; Lebrun, M.; Czernic, P. Root-to-Shoot Long-Distance Circulation of Nicotianamine and Nicotianamine-Nickel Chelates in the Metal Hyperaccumulator Thlaspi caerulescens. J. Exp. Bot. 2006, 57, 4111–4122. [Google Scholar] [CrossRef] [PubMed]
- Milner, M.J.; Kochian, L.V. Investigating Heavy-Metal Hyperaccumulation Using Thlaspi caerulescens as a Model System. Ann. Bot. 2008, 102, 3–13. [Google Scholar] [CrossRef]
- Roccotiello, E.; Zotti, M.; Mesiti, S.; Marescotti, P.; Carbone, C.; Cornara, L.; Mariotti, M.G. Biodiversity in Metal-Polluted Soils. Fresenius Environ. Bull. 2010, 19, 2420–2425. [Google Scholar]
- Whiting, S.N.; Leake, J.R.; McGrath, S.P.; Baker, A.J.M. Positive Responses to Zn and Cd by Roots of the Zn and Cd Hyperaccumulator Thlaspi caerulescens. New Phytol. 2000, 145, 199–210. [Google Scholar] [CrossRef]
- Seregin, I.V.; Erlikh, N.T.; Kozhevnikova, A.D. Nickel and Zinc Accumulation Capacities and Tolerance to These Metals in the Excluder Thlaspi arvense and the Hyperaccumulator Noccaea caerulescens. Russ. J. Plant Physiol. 2014, 61, 204–214. [Google Scholar] [CrossRef]
- Kidd, P.; Barceló, J.; Bernal, M.P.; Navari-Izzo, F.; Poschenrieder, C.; Shilev, S.; Clemente, R.; Monterroso, C. Trace Element Behaviour at the Root–Soil Interface: Implications in Phytoremediation. Environ. Exp. Bot. 2009, 67, 243–259. [Google Scholar] [CrossRef]
- Chaney, R.L.; Angle, J.S.; Broadhurst, C.L.; Peters, C.A.; Tappero, R.V.; Sparks, D.L. Improved Understanding of Hyperaccumulation Yields Commercial Phytoextraction and Phytomining Technologies. J. Environ. Qual. 2007, 36, 1429–1443. [Google Scholar] [CrossRef]
- Ryser, P.; Sauder, W.R. Effects of Heavy-Metal-Contaminated Soil on Growth, Phenology and Biomass Turnover of Hieracium piloselloides. Environ. Pollut. 2006, 140, 52–61. [Google Scholar] [CrossRef]
- Al Agely, A.; Sylvia, D.M.; Ma, L.Q. Mycorrhizae Increase Arsenic Uptake by the Hyperaccumulator Chinese Brake Fern ( Pteris vittata L.). J. Environ. Qual. 2005, 34, 2181–2186. [Google Scholar] [CrossRef]
- Trotta, A.; Falaschi, P.; Cornara, L.; Minganti, V.; Fusconi, A.; Drava, G.; Berta, G. Arbuscular Mycorrhizae Increase the Arsenic Translocation Factor in the As Hyperaccumulating Fern Pteris vittata L. Chemosphere 2006, 65, 74–81. [Google Scholar] [CrossRef]
- Nkrumah, P.N.; Echevarria, G.; Erskine, P.D.; van der Ent, A. Nickel Hyperaccumulation in Antidesma montissilam: From Herbarium Discovery to Collection in the Native Habitat. Ecol. Res. 2018, 1–11. [Google Scholar] [CrossRef]
- Mirouze, M.; Paszkowski, J. Epigenetic Contribution to Stress Adaptation in Plants. Curr. Opin. Plant Biol. 2011, 14, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Zhang, Y.; Xu, C.; Lin, X.; Zang, Q.; Zhuang, T.; Jiang, L.; von Wettstein, D.; Liu, B. Transgenerational Inheritance of Modified DNA Methylation Patterns and Enhanced Tolerance Induced by Heavy Metal Stress in Rice (Oryza sativa L.). PLoS ONE 2012, 7, e41143. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, L.; Song, J.; Cui, W.; Zhang, Y.; Jia, C.; Francis, D.; Rogers, H.J.; Sun, L.; Tai, P. Cadmium-Induced Genomic Instability in Arabidopsis: Molecular Toxicological Biomarkers for Early Diagnosis of Cadmium Stress. Chemosphere 2016, 150, 258–265. [Google Scholar] [CrossRef]
- Adamidis, G.C.; Aloupi, M.; Kazakou, E.; Dimitrakopoulos, P.G. Intra-Specific Variation in Ni Tolerance, Accumulation and Translocation Patterns in the Ni-Hyperaccumulator Alyssum lesbiacum. Chemosphere 2014, 95, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Roccotiello, E.; Serrano, H.C.; Mariotti, M.G.; Branquinho, C. The Impact of Ni on the Physiology of a Mediterranean Ni-Hyperaccumulating Plant. Environ. Sci. Pollut. Res. 2016, 23, 12414–12422. [Google Scholar] [CrossRef]
- Pietrini, F.; Passatore, L.; Patti, V.; Francocci, F.; Giovannozzi, A.; Zacchini, M. Morpho-Physiological and Metal Accumulation Responses of Hemp Plants (Cannabis sativa L.) Grown on Soil from an Agro-Industrial Contaminated Area. Water 2019, 11, 808. [Google Scholar] [CrossRef]
- Wu, J.; Hu, J.; Wang, L.; Zhao, L.; Ma, F. Responses of Phragmites australis to Copper Stress: A Combined Analysis of Plant Morphology, Physiology and Proteomics. Plant Biol. (Stuttg) 2020. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 445–483. [Google Scholar]
- Živčák, M.; Olšovská, K.; Slamka, P.; Galambošová, J.; Rataj, V.; Shao, H.B.; Brestič, M. Application of Chlorophyll Fluorescence Performance Indices to Assess the Wheat Photosynthetic Functions Influenced by Nitrogen Deficiency. Plant Soil Environ. 2015, 60, 210–215. [Google Scholar] [CrossRef]
- Marchand, L.; Lamy, P.; Bert, V.; Quintela-Sabaris, C.; Mench, M. Potential of Ranunculus acris L. for Biomonitoring Trace Element Contamination of Riverbank Soils: Photosystem II Activity and Phenotypic Responses for Two Soil Series. Environ. Sci. Pollut. Res. 2016, 23, 3104–3119. [Google Scholar] [CrossRef] [PubMed]
- Quintela-Sabaris, C.; Marchand, L.; Kidd, P.S.; Friesl-Hanl, W.; Puschenreiter, M.; Kumpiene, J.; Mueller, I.; Neu, S.; Janssen, J.; Vangronsveld, J. Assessing Phytotoxicity of Trace Element-Contaminated Soils Phytomanaged with Gentle Remediation Options at Ten European Field Trials. Sci. Total Environ. 2017, 599, 1388–1398. [Google Scholar] [CrossRef]
- Visioli, G.; Vincenzi, S.; Marmiroli, M.; Marmiroli, N. Correlation between Phenotype and Proteome in the Ni Hyperaccumulator Noccaea caerulescens subsp. caerulescens. Environ. Exp. Bot. 2012, 77, 156–164. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular Mechanisms of Metal Hyperaccumulation in Plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef]
- Bakker, P.A.H.M.; Berendsen, R.L.; Doornbos, R.F.; Wintermans, P.C.A.; Pieterse, C.M.J. The Rhizosphere Revisited: Root Microbiomics. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chiarucci, A.; Fang, H.; Chen, M. An Interspecific Variation in Rhizosphere Effects on Soil Anti-Erodibility. Sci. Rep. 2020, 10, 2411. [Google Scholar] [CrossRef]
- Li, H.; Su, J.-Q.; Yang, X.-R.; Zhu, Y.-G. Distinct Rhizosphere Effect on Active and Total Bacterial Communities in Paddy Soils. Sci. Total Environ. 2019, 649, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.M. Introduction: Some Consequences of Microbial Rhizosphere Competence for Plant and Soil. Rhizosphere 1990, 1–10. [Google Scholar]
- Sørensen, J. The Rhizosphere as a Habitat for Soil Microorganisms. Mod. Soil Microbiol. 1997, 21–45. [Google Scholar]
- Foster, R.C.; Rovira, A.D.; Cock, T.W. Ultrastructure of the Root-Soil Interface; American Phytopathological Society: Saint Paul, MN, USA, 1983. [Google Scholar]
- Xie, Y.; Fan, J.; Zhu, W.; Amombo, E.; Lou, Y.; Chen, L.; Fu, J. Effect of Heavy Metals Pollution on Soil Microbial Diversity and Bermudagrass Genetic Variation. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Liao, M.; Xie, X.M. Effect of Heavy Metals on Substrate Utilization Pattern, Biomass, and Activity of Microbial Communities in a Reclaimed Mining Wasteland of Red Soil Area. Ecotox. Environ. Saf. 2007, 66, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Meng, D.; Li, J.; Yin, H.; Liu, H.; Liu, X.; Cheng, C.; Xiao, Y.; Liu, Z.; Yan, M. Response of Soil Microbial Communities and Microbial Interactions to Long-Term Heavy Metal Contamination. Environ. Pollut. 2017, 231, 908–917. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, J.; Wang, H.; Lin, Q.; Chen, X.; Chen, Y. The Influence of Soil Heavy Metals Pollution on Soil Microbial Biomass, Enzyme Activity, and Community Composition near a Copper Smelter. Ecotox. Environ. Saf. 2007, 67, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Azarbad, H.; Niklińska, M.; van Gestel, C.A.; van Straalen, N.M.; Röling, W.F.; Laskowski, R. Microbial Community Structure and Functioning along Metal Pollution Gradients. Environ. Toxicol. Chem. 2013, 32, 1992–2002. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Meng, Q.; Luo, S.; Shen, J.; Chen, B.; Khan, K.Y.; Japenga, J.; Ma, X.; Yang, X.; Feng, Y. Enhanced Cd Extraction of Oilseed Rape (Brassica napus) by Plant Growth-Promoting Bacteria Isolated from Cd Hyperaccumulator Sedum Alfredii Hance. Int. J. Phytoremediat. 2017, 19, 281–289. [Google Scholar] [CrossRef]
- Sessitsch, A.; Kuffner, M.; Kidd, P.; Vangronsveld, J.; Wenzel, W.W.; Fallmann, K.; Puschenreiter, M. The Role of Plant-Associated Bacteria in the Mobilization and Phytoextraction of Trace Elements in Contaminated Soils. Soil Biol. Biochem. 2013, 60, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Vara Prasad, M.N.; Freitas, H.; Ae, N. Biotechnological Applications of Serpentine Soil Bacteria for Phytoremediation of Trace Metals. Crit. Rev. Biotechnol. 2009, 29, 120–130. [Google Scholar] [CrossRef]
- Khan, A.G. Role of Soil Microbes in the Rhizospheres of Plants Growing on Trace Metal Contaminated Soils in Phytoremediation. J. Trace Elem. Med. Biol. 2005, 18, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Castro, C.; Prieto-Fernández, A.; Alvarez-Lopez, V.; Monterroso, C.; Cabello-Conejo, M.I.; Acea, M.J.; Kidd, P.S. Nickel Solubilizing Capacity and Characterization of Rhizobacteria Isolated from Hyperaccumulating and Non-Hyperaccumulating Subspecies of Alyssum serpyllifolium. Int. J. Phytoremediat. 2011, 13 (Suppl. 1), 229–244. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Sultana, F.; Islam, S. Plant Growth-Promoting Fungi (PGPF): Phytostimulation and Induced Systemic Resistance. In Plant-Microbe Interactions in Agro-Ecological Perspectives; Springer: Singapore, 2017; pp. 135–191. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F. Application and Mechanisms of Plant Growth Promoting Fungi (PGPF) for Phytostimulation. Org. Agric. 2020. [Google Scholar] [CrossRef]
- Thijs, S.; Langill, T.; Vangronsveld, J. Chapter Two—The Bacterial and Fungal Microbiota of Hyperaccumulator Plants: Small Organisms, Large Influence. In Advances in Botanical Research; Cuypers, A., Vangronsveld, J., Eds.; Phytoremediation: Oxford, UK; Elsevier Ltd.: Oxford, UK, 2017; Volume 83, pp. 43–86. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia. Edagricole: Bologna, Italy, 1982; 3 Vols. [Google Scholar]
- Assunção, A.G.L.; Bleeker, P.; ten Bookum, W.M.; Vooijs, R.; Schat, H. Intraspecific Variation of Metal Preference Patterns for Hyperaccumulation in Thlaspi caerulescens: Evidence from Binary Metal Exposures. Plant Soil 2008, 303, 289–299. [Google Scholar] [CrossRef]
- Macnair, M.R. The Hyperaccumulation of Metals by Plants. Adv. Bot. Res. 2003, 40, 63–105. [Google Scholar] [CrossRef]
- Fones, H.N.; Preston, G.M.; Smith, J.A.C. Variation in Defence Strategies in the Metal Hyperaccumulator Plant Noccaea caerulescens Is Indicative of Synergies and Trade-Offs between Forms of Defence. R. Soc. Open Sci. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Halimaa, P.; Blande, D.; Aarts, M.G.M.; Tuomainen, M.; Tervahauta, A.; Kärenlampi, S. Comparative Transcriptome Analysis of the Metal Hyperaccumulator Noccaea caerulescens. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Visioli, G.; Gullì, M.; Marmiroli, N. Noccaea caerulescens Populations Adapted to Grow in Metalliferous and Non-Metalliferous Soils: Ni Tolerance, Accumulation and Expression Analysis of Genes Involved in Metal Homeostasis. Environ. Exp. Bot. 2014, 105, 10–17. [Google Scholar] [CrossRef]
- Gonneau, C. Distribution, Écologie et Évolution de l’hyperaccumulation Des Éléments En Traces Par Noccaea caerulescens. Ph.D. Thesis, Université de Lorraine, Metz, France, 2014. [Google Scholar]
- Sterckeman, T.; Cazes, Y.; Gonneau, C.; Sirguey, C. Phenotyping 60 Populations of Noccaea caerulescens Provides a Broader Knowledge of Variation in Traits of Interest for Phytoextraction. Plant Soil 2017, 418, 523–540. [Google Scholar] [CrossRef]
- Agrawal, B.; Czymmek, K.J.; Sparks, D.L.; Bais, H.P. Transient Influx of Nickel in Root Mitochondria Modulates Organic Acid and Reactive Oxygen Species Production in Nickel Hyperaccumulator Alyssum Murale. J. Biol. Chem. 2013, 288, 7351–7362. [Google Scholar] [CrossRef]
- Broadhurst, C.L.; Chaney, R.L. Growth and Metal Accumulation of an Alyssum murale Nickel Hyperaccumulator Ecotype Co-Cropped with Alyssum montanum and Perennial Ryegrass in Serpentine Soil. Front. Plant Sci. 2016, 7, 451. [Google Scholar] [CrossRef] [PubMed]
- Kozhevnikova, A.D.; Seregin, I.V.; Erlikh, N.T.; Shevyreva, T.A.; Andreev, I.M.; Verweij, R.; Schat, H. Histidine-Mediated Xylem Loading of Zinc Is a Species-Wide Character in Noccaea caerulescens. New Phytol. 2014, 203, 508–519. [Google Scholar] [CrossRef]
- Arrigo, N.; de Harpe, M.L.; Litsios, G.; Zozomová-Lihová, J.; Španiel, S.; Marhold, K.; Barker, M.S.; Alvarez, N. Is Hybridization Driving the Evolution of Climatic Niche in Alyssum montanum. Am. J. Bot. 2016, 103, 1348–1357. [Google Scholar] [CrossRef]
- Rešetnik, I.; Satovic, Z.; Schneeweiss, G.M.; Liber, Z. Phylogenetic Relationships in Brassicaceae Tribe Alysseae Inferred from Nuclear Ribosomal and Chloroplast DNA Sequence Data. Mol. Phylogenet. Evol. 2013, 69, 772–786. [Google Scholar] [CrossRef]
- Li, Y.; Kong, Y.; Zhang, Z.; Yin, Y.; Liu, B.; Lv, G.; Wang, X. Phylogeny and Biogeography of Alyssum (Brassicaceae) Based on Nuclear Ribosomal ITS DNA Sequences. J. Genet. 2014, 93, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Jalas, J. Atlas Florae Europaeae Notes. 13. Suggestions on Alyssum and Lepidium (Cruciferae). Ann. Bot. Fennici 1996, 33, 283–284. [Google Scholar]
- Španiel, S.; Marhold, K.; Filová, B.; Zozomová-Lihová, J. Genetic and Morphological Variation in the Diploid–Polyploid Alyssum montanum in Central Europe: Taxonomic and Evolutionary Considerations. Plant Syst. Evol. 2011, 294, 1. [Google Scholar] [CrossRef]
- Španiel, S.; Marhold, K.; Passalacqua, N.G.; Zozomová-Lihová, J. Intricate Variation Patterns in the Diploid-Polyploid Complex of Alyssum montanum—A. repens (Brassicaceae) in the Apennine Peninsula: Evidence for Long-Term Persistence and Diversification. Am. J. Bot. 2011, 98, 1887–1904. [Google Scholar] [CrossRef]
- Zozomová-Lihová, J.; Marhold, K.; Španiel, S. Taxonomy and Evolutionary History of Alyssum montanum (Brassicaceae) and Related Taxa in Southwestern Europe and Morocco: Diversification Driven by Polyploidy, Geographic and Ecological Isolation. Taxon 2014, 63, 562–591. [Google Scholar] [CrossRef]
- Dorn, K.M.; Johnson, E.B.; Daniels, E.C.; Wyse, D.L.; Marks, M.D. Spring Flowering Habit in Field Pennycress (Thlaspi arvense) Has Arisen Multiple Independent Times. Plant Direct. 2018, 2. [Google Scholar] [CrossRef] [PubMed]
- McGinn, M.; Phippen, W.B.; Chopra, R.; Bansal, S.; Jarvis, B.A.; Phippen, M.E.; Dorn, K.M.; Esfahanian, M.; Nazarenus, T.J.; Cahoon, E.B.; et al. Molecular Tools Enabling Pennycress (Thlaspi arvense) as a Model Plant and Oilseed Cash Cover Crop. Plant Biotechnol. J. 2019, 17, 776–788. [Google Scholar] [CrossRef]
- Beilstein, M.A.; Al-Shehbaz, I.A.; Mathews, S.; Kellogg, E.A. Brassicaceae Phylogeny Inferred from Phytochrome A and NdhF Sequence Data: Tribes and Trichomes Revisited. Am. J. Bot. 2008, 95, 1307–1327. [Google Scholar] [CrossRef]
- Thomas, J.B.; Hampton, M.E.; Dorn, K.M.; David Marks, M.; Carter, C.J. The Pennycress (Thlaspi arvense L.) Nectary: Structural and Transcriptomic Characterization. BMC Plant Biol. 2017, 17, 201. [Google Scholar] [CrossRef]
- Warwick, S.I.; Francis, A.; Susko, D.J. The Biology of Canadian Weeds. 9. Thlaspi arvense L.(Updated). Can. J. Plant Sci. 2002, 82, 803–823. [Google Scholar] [CrossRef]
- Levy, A.; Gonzalez, I.S.; Mittelviefhaus, M.; Clingenpeel, S.; Paredes, S.H.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F. Genomic Features of Bacterial Adaptation to Plants. Nat. Genet. 2018, 50, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Isbell, T. Thlaspi arvense (Pennycress) as a Biodiesel in a One Year-Two Crop Rotation with Soybean. In Association for the Advancement of Industrial Crops Conference; Agricultural Research Service U.S. Department of Agriculture: Washington, DC, USA, 2008; Volume 6. [Google Scholar]
- Eberle, C.A.; Thom, M.D.; Nemec, K.T.; Forcella, F.; Lundgren, J.G.; Gesch, R.W.; Riedell, W.E.; Papiernik, S.K.; Wagner, A.; Peterson, D.H. Using Pennycress, Camelina, and Canola Cash Cover Crops to Provision Pollinators. Ind. Crop. Prod. 2015, 75, 20–25. [Google Scholar] [CrossRef]
- Groeneveld, J.H.; Klein, A.-M. Pennycress-Corn Double-Cropping Increases Ground Beetle Diversity. Biomass Bioenerg. 2015, 77, 16–25. [Google Scholar] [CrossRef]
- Johnson, G.A.; Kantar, M.B.; Betts, K.J.; Wyse, D.L. Field Pennycress Production and Weed Control in a Double Crop System with Soybean in Minnesota. Agron. J. 2015, 107, 532–540. [Google Scholar] [CrossRef]
- Thom, M.D.; Forcella, F.; Eberle, C.A.; Matthees, H.L.; Weyers, S.L.; Gesch, R.W.; Ott, M.A.; Feyereisen, G.W.; Strock, J.S.; Wyse, D. Reduced-Nutrient Leachates in Cash Cover Crop-Soybean Systems. BioRxiv 2018, 254169. [Google Scholar] [CrossRef]
- Charlot, G. Colorimetric Determination of Elements, Principles and Methods: By G. Charlot; Elsevier: Amsterdam, The Netherlands, 1964. [Google Scholar]
- Küpper, H.; Lombi, E.; Zhao, F.-J.; Wieshammer, G.; McGrath, S.P. Cellular Compartmentation of Nickel in the Hyperaccumulators Alyssum lesbiacum, Alyssum bertolonii and Thlaspi goesingense. J. Exp. Bot. 2001, 52, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, K.; Aggarwal, P.; Chakraborty, D.; Pradhan, S.; Narayan Garg, R.; Singh, R. Practical Manual on Measurement of Soil Physical Properties Practical; Division of Agricultural Physics, Indian Agricultural Research Institute: New Delhi, India, 2012. [Google Scholar]
- Baiyeri, K.P.; Mbah, B.N. Surface Sterilization and Duration of Seed Storage Influenced Emergence and Seedling Quality of African Breadfruit (Treculia africana Decne). Afr. J. Biotechnol. 2006, 5, 1393–1396. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Method 6200: Field Portable X-ray Fluorescence Spectrometry for the Determination of Elemental Concentrations in Soil and Sediment; Environmental Protection Agency: Washington, DC, USA.
- Murchie, E.H.; Lawson, T. Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M. PSII Fluorescence Techniques for Measurement of Drought and High Temperature Stress Signal in Crop Plants: Protocols and Applications. In Molecular Stress Physiology of Plants; Rout, G.R., Das, A.B., Eds.; Springer India: New Delhi, India, 2013; pp. 87–131. ISBN 978-81-322-0807-5. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in Vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Bosa, K.; Kościelniak, J.; Żuk-Gołaszewska, K. Effects of Salt Stress on Photosystem II Efficiency and CO2 Assimilation of Two Syrian Barley Landraces. Environ. Exp. Bot. 2011, 73, 64–72. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Strasser, R.J. Experimental in Vivo Measurements of Light Emission in Plants: A Perspective Dedicated to David Walker. Photosynth. Res. 2012, 114, 69–96. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P. Frequently Asked Questions about in Vivo Chlorophyll Fluorescence: Practical Issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [PubMed]
- Furbank, R.T.; Caemmerer, S.; von Sheehy, J.; Edwards, G. C4 Rice: A Challenge for Plant Phenomics. Funct. Plant Biol. 2009, 36, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Montes, J.M.; Melchinger, A.E.; Reif, J.C. Novel Throughput Phenotyping Platforms in Plant Genetic Studies. Trends Plant Sci. 2007, 12, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W.S. 2011. ImageJ; US National Institutes of Health: Bethesda, MD, USA, 1997. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Parameters | Ni Treatment | |||
|---|---|---|---|---|
| Au++ | Nc++ | Am+ | Ta+ | |
| Root surface area | NS | NS | NS | 0.74 *** |
| FW_root | NS | NS | NS | 0.66 *** |
| DW_root | 0.53 ** | NS | NS | 0.78 *** |
| FW_shoot | 0.61 *** | NS | NS | NS |
| DW_shoot | 0.61 *** | NS | NS | 0.48 ** |
| R/S FW | NS | −0.42 * | NS | 0.68 *** |
| R/S DW | NS | NS | NS | 0.64 *** |
| % water R | NS | 0.54 ** | NS | NS |
| % water S | NS | NS | NS | 0.60 *** |
| Species | Ni Treatment (mg kg−1) | Ni Concentration (mg kg−1) |
|---|---|---|
| A. utriculata ++ | 0 | 18.33 ± 2.00 |
| 1000 | 999.67 ± 8.00 | |
| N. caerulescens ++ | 0 | 76.67 ± 3.00 |
| 1000 | 6798 ± 24.33 | |
| A. montanum + | 0 | 4.00 ± 1.33 |
| 1000 | 127.33 ± 4.00 | |
| T. arvense + | 0 | 0.00 |
| 1000 | 177.00 ± 5.00 |
| Parameters | Ni Treatment | |||
|---|---|---|---|---|
| Au++ | Nc++ | Am+ | Ta+ | |
| F0 | NS | NS | 0.25 *** | −0.16 ** |
| Fm | NS | −0.17 ** | 0.15 * | NS |
| Fv/Fm | NS | NS | −0.22 ** | 0.24 *** |
| Tfm (ms) | 0.38 *** | 0.37 *** | NS | 0.25 *** |
| Area | 0.30 *** | NS | NS | 0.19 *** |
| P.I. | 0.13 * | NS | −0.21 ** | 0.31 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosatto, S.; Mariotti, M.; Romeo, S.; Roccotiello, E. Root and Shoot Response to Nickel in Hyperaccumulator and Non-Hyperaccumulator Species. Plants 2021, 10, 508. https://doi.org/10.3390/plants10030508
Rosatto S, Mariotti M, Romeo S, Roccotiello E. Root and Shoot Response to Nickel in Hyperaccumulator and Non-Hyperaccumulator Species. Plants. 2021; 10(3):508. https://doi.org/10.3390/plants10030508
Chicago/Turabian StyleRosatto, Stefano, Mauro Mariotti, Sara Romeo, and Enrica Roccotiello. 2021. "Root and Shoot Response to Nickel in Hyperaccumulator and Non-Hyperaccumulator Species" Plants 10, no. 3: 508. https://doi.org/10.3390/plants10030508
APA StyleRosatto, S., Mariotti, M., Romeo, S., & Roccotiello, E. (2021). Root and Shoot Response to Nickel in Hyperaccumulator and Non-Hyperaccumulator Species. Plants, 10(3), 508. https://doi.org/10.3390/plants10030508