
Effects of Acute and Chronic Gamma Irradiation on the Cell Biology and Physiology of Rice Plants

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. DNA Damage
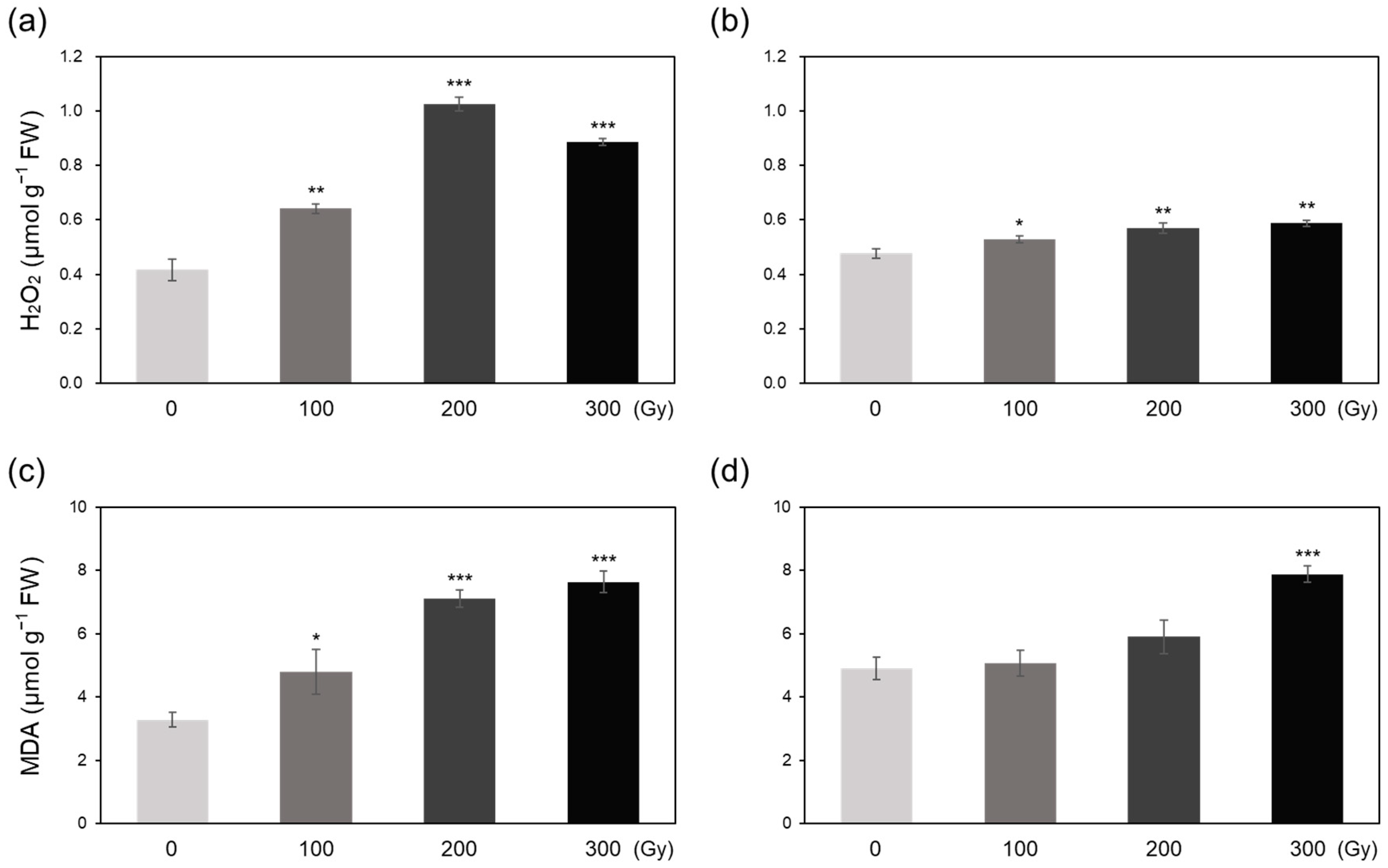
2.2. Free Radicals, H2O2, and Lipid Peroxidation
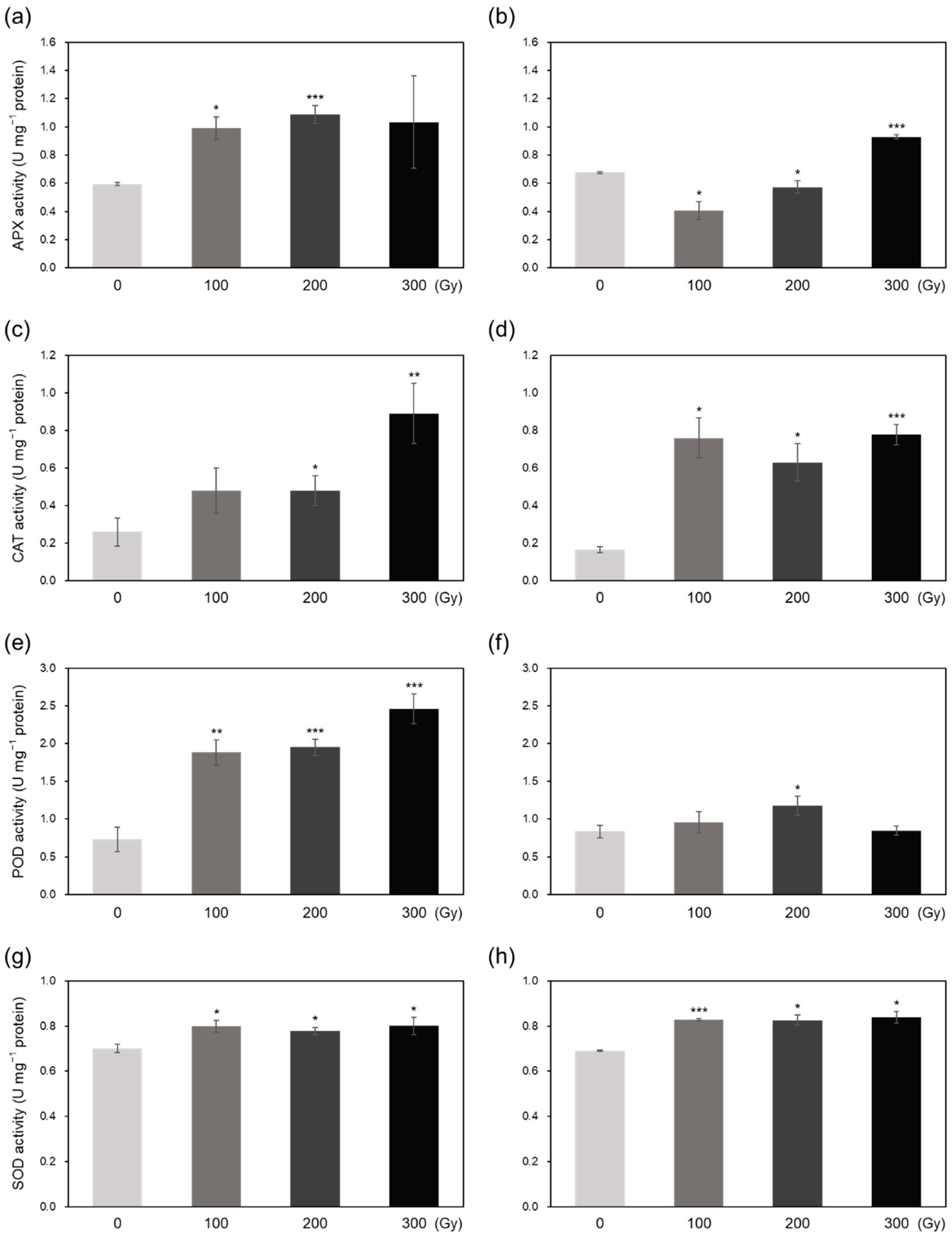
2.3. Superoxide Radical Scavenging and Antioxidant Activities
2.4. Photosynthesis Efficiency
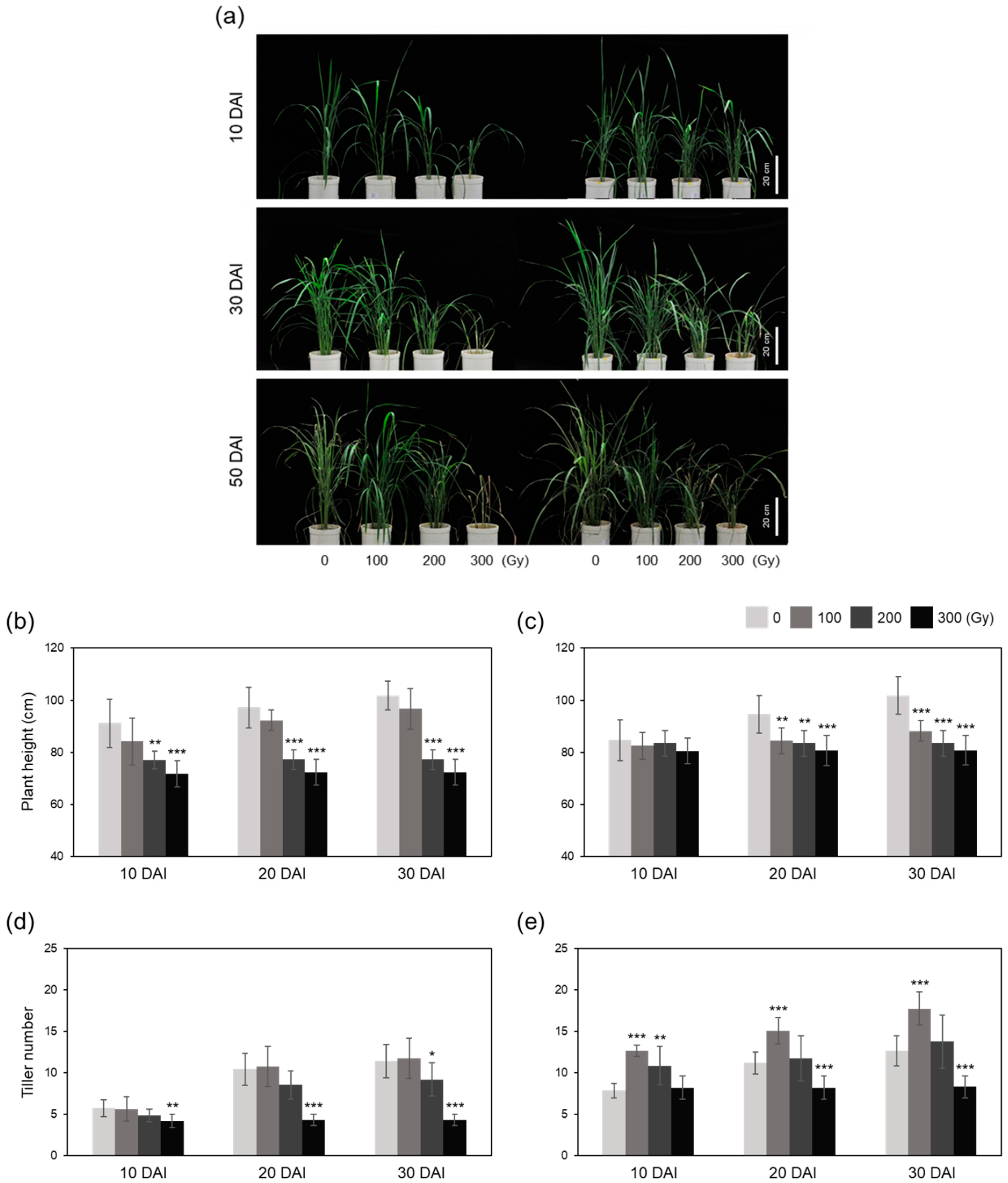
2.5. Growth and Reproduction
3. Discussion
4. Materials and Methods
4.1. Sample Preparation, Growth, and Growth Test after Irradiation
4.2. Comet Assay
4.3. Electron Spin Resonance (ESR) Analysis
4.4. H2O2 and Malonedialdehyde (MDA) Content
4.5. Superoxide Radical Scavenging Activity
4.6. Antioxidant Enzyme Activity
4.7. Phenylanine Ammonia-Lyase (PAL) Activity, Total Phenolic Content, and Ascorbic Acid (AsA) Content
4.8. Chlorophyll Fluorescence
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Caplin, N.; Willey, N. Ionizing radiation, higher plants, and radioprotection: From acute high doses to chronic low doses. Front. Plant Sci. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, L. Origin of the gamma rays*. Nature 1932, 129, 457–458. [Google Scholar] [CrossRef]
- Gerward, L. Paul Villard and his discovery of gamma rays. Phys. Perspect. 1999, 1, 367–383. [Google Scholar] [CrossRef]
- Mba, C.; Shu, Q.Y. Gamma irradiation. In Plant Mutation Breeding and Biotechnology; Shu, Q.Y., Forster, B.P., Nakagawa, H., Eds.; CABI: Oxfordshire, UK, 2012; pp. 91–98. [Google Scholar]
- Hase, Y.; Satoh, K.; Seito, H.; Oono, Y. Genetic consequences of acute/chronic gamma and carbon ion irradiation of Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 336. [Google Scholar] [CrossRef] [PubMed]
- Esnault, M.A.; Legue, F.; Chenal, C. Ionizing radiation: Advances in plant response. Environ. Exp. Bot. 2010, 68, 231–237. [Google Scholar] [CrossRef]
- Desouky, O.; Ding, N.; Zhou, G. Targeted and non-targeted effects of ionizing radiation. J. Radiat. Res. Appl. Sci. 2015, 8, 247–254. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Roldán-Arjona, T.; Ariza, R.R. Repair and tolerance of oxidative DNA damage in plants. Mutat. Res. 2009, 681, 169–179. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, D.Y.; Ahn, J.W.; Kang, S.Y.; Seo, Y.W.; Kim, J.B. Comparison of radiosensitivity response to acute and chronic gamma irradiation in colored wheat. Genet. Mol. Biol. 2018, 41, 611–623. [Google Scholar] [CrossRef]
- Baldwin, J.; Grantham, V. Radiation hormesis: Historical and current perspectives. J. Nucl. Med. Technol. 2015, 43, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Jan, S.; Parween, T.; Siddiqi, T.O.; Mahmooduzzafar, X. Effect of gamma radiation on morphological, biochemical, and physiological aspects of plants and plant products. Environ. Rev. 2012, 20, 17–39. [Google Scholar] [CrossRef]
- Marcu, D.; Damian, G.; Cosma, C.; Cristea, V. Gamma radiation effects on seed germination, growth and pigment content, and ESR Study of induced free radicals in maize (Zea Mays). J. Biol. Phys. 2013, 39, 625–634. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Arena, C.; Pignalosa, D.; Durante, M. Effects of sparsely and densely ionizing radiation on plants. Radiat. Environ. Biophys. 2011, 50, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Wi, S.G.; Chung, B.Y.; Kim, J.H.; Baek, M.H.; Yang, D.H.; Lee, J.W.; Kim, J.S. Ultrastructural changes of cell organelles in Arabidopsis stems after gamma irradiation. J. Plant Biol. 2005, 48, 195–200. [Google Scholar] [CrossRef]
- Wi, S.G.; Chung, B.Y.; Kim, J.S.; Kim, J.H.; Baek, M.H.; Lee, J.W.; Kim, Y.S. Effects of gamma irradiation on morphological changes and biological responses in plants. Micron 2007, 38, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Nandhakumar, S.; Parasuraman, S.; Shanmugam, M.M.; Ramachandra, K.; Chand, R.P.; Bhat, B.V. Evaluation of DNA damage using single-cell gel electrophoresis (Comet Assay). J. Pharmacol. Pharmacother. 2011, 2, 107–111. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, Y.S.; Jo, Y.D.; Ha, B.K.; Kim, D.S.; Kim, J.B.; Kang, S.Y.; Kim, S.H. Oxidative stress and DNA damage induced by gamma irradiation in Korean lawngrass (Zoysia Japonica Steud.). Eur. J. Hortic. Sci. 2016, 81, 303–309. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Goh, E.J.; Kim, J.B.; Kim, W.J.; Ha, B.K.; Kim, S.H.; Kang, S.Y.; Seo, Y.W.; Kim, D.S. Physiological changes and anti-oxidative responses of Arabidopsis plants after acute and chronic γ-irradiation. Radiat. Environ. Biophys. 2014, 53, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.Y.; Sattler, S.A.; Cortez, G.S.; Vermerris, W.; Sattler, S.E.; Kang, C.H. Biochemical and structural analysis of substrate specificity of a phenylalanine ammonia-lyase. Plant Physiol. 2018, 176, 1452–1468. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Hussain, P.R.; Wani, A.M.; Meena, R.S.; Dar, M.A. Gamma irradiation induced enhancement of phenylalanine ammonia-lyase (PAL) and antioxidant activity in peach (Prunus Persica Bausch, Cv. Elberta). Radiat. Phys. Chem. 2010, 79, 982–989. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Photosynthesis. In Plant Physiological Ecology; Springer: New York, NY, USA, 2008; pp. 11–99. [Google Scholar]
- Shcherbakov, V.P. Biological species is the only possible form of existence for higher organisms: The evolutionary meaning of sexual reproduction. Biol. Direct 2010, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Reznick, D. Costs of reproduction: An evaluation of the empirical evidence. Oikos 1985, 44, 257–267. [Google Scholar] [CrossRef]
- Obeso, J.R. The costs of reproduction in plants. New Phytol. 2002, 155, 321–348. [Google Scholar] [CrossRef]
- Tsai, C.H.; Chang, R.C.; Chiou, J.F.; Liu, T.Z. Improved superoxide-generating system suitable for the assessment of the superoxide-scavenging ability of aqueous extracts of food constituents using ultraweak chemiluminescence. J. Agric. Food Chem. 2003, 51, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Goldson, A.; Lam, M.; Scaman, C.H.; Clemens, S.; Kermode, A. Screening of phenylalanine ammonia lyase in plant tissues, and retention of activity during dehydration. J. Sci. Food Agric. 2008, 88, 619–625. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using folin–ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode | Dose (Gy) | PN 1 | PL (cm) 2 | SN 3 | FR (%) 4 |
|---|---|---|---|---|---|
| Acute irradiation | 0 | 4.6 ± 0.5 | 18.4 ± 1.1 | 98.6 ± 20.7 | 71.1 ± 16.3 |
| 100 | 4.1 ± 1.0 | 17.1 ± 2.0 ** | 63.6 ± 21.4 *** | 42.6 ± 19.4 *** | |
| 200 | 2.0 ± 0.8 *** | 11.3 ± 2.1 *** | 24.4 ± 12.1 *** | 5.0 ± 5.5 *** | |
| 300 | – | – | – | – | |
| Chronic irradiation | 0 | 3.3 ± 0.8 | 17.2 ± 1.0 | 58.2 ± 18.9 | 86.6 ± 8.6 |
| 100 | 2.1 ± 0.7 *** | 14.9 ± 2.8 ** | 44.7 ± 20.6 | 0 | |
| 200 | – | – | – | – | |
| 300 | – | – | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.-I.; Han, S.M.; Jo, Y.D.; Hong, M.J.; Kim, S.H.; Kim, J.-B. Effects of Acute and Chronic Gamma Irradiation on the Cell Biology and Physiology of Rice Plants. Plants 2021, 10, 439. https://doi.org/10.3390/plants10030439
Choi H-I, Han SM, Jo YD, Hong MJ, Kim SH, Kim J-B. Effects of Acute and Chronic Gamma Irradiation on the Cell Biology and Physiology of Rice Plants. Plants. 2021; 10(3):439. https://doi.org/10.3390/plants10030439
Chicago/Turabian StyleChoi, Hong-Il, Sung Min Han, Yeong Deuk Jo, Min Jeong Hong, Sang Hoon Kim, and Jin-Baek Kim. 2021. "Effects of Acute and Chronic Gamma Irradiation on the Cell Biology and Physiology of Rice Plants" Plants 10, no. 3: 439. https://doi.org/10.3390/plants10030439
APA StyleChoi, H.-I., Han, S. M., Jo, Y. D., Hong, M. J., Kim, S. H., & Kim, J.-B. (2021). Effects of Acute and Chronic Gamma Irradiation on the Cell Biology and Physiology of Rice Plants. Plants, 10(3), 439. https://doi.org/10.3390/plants10030439