
Oxidative Stress and Antioxidative Activity in Leaves and Roots of Carrot Plants Induced by Candidatus Phytoplasma Solani


 , ,
, ,
Abstract
1. Introduction
2. Results
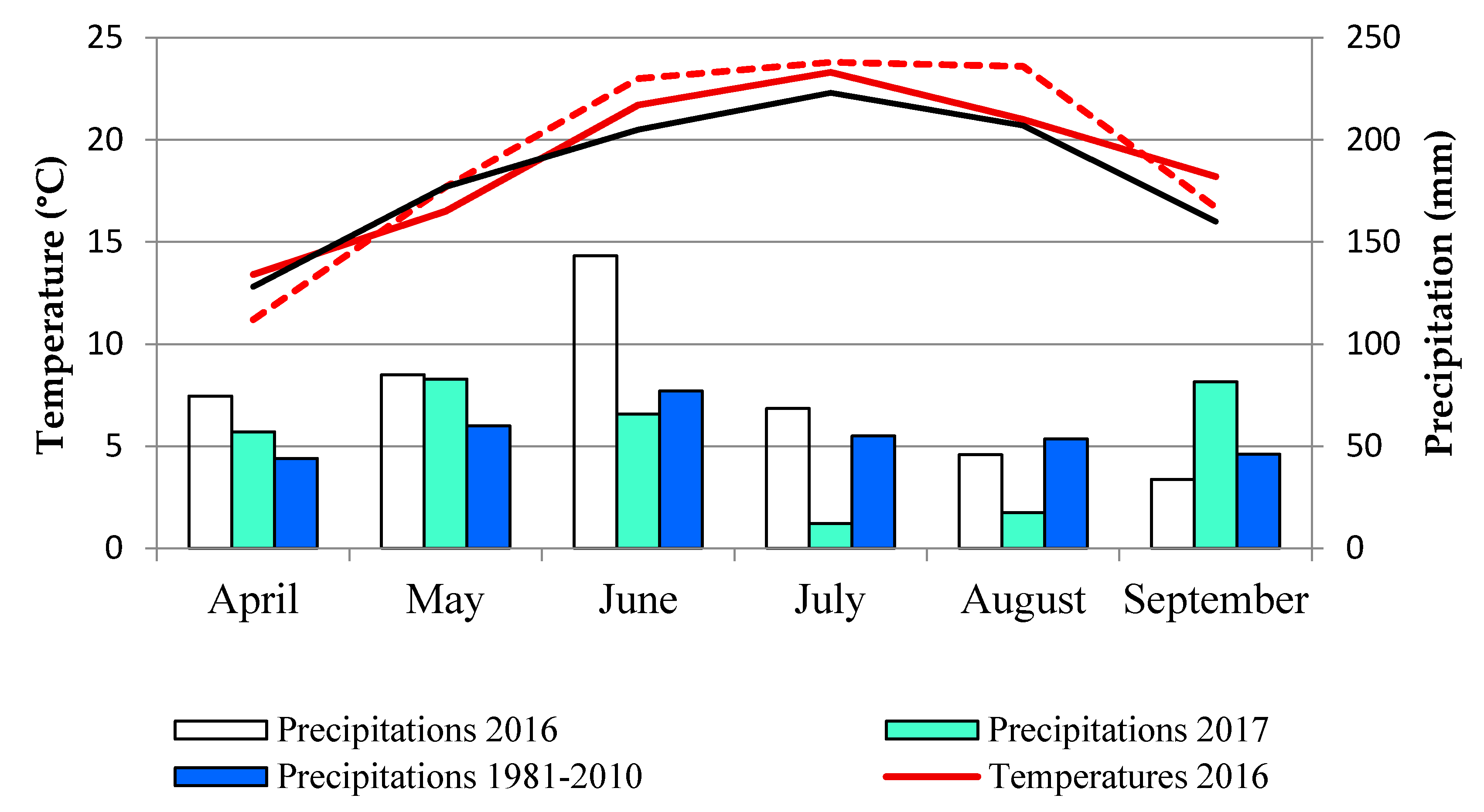
2.1. Symptoms Observation
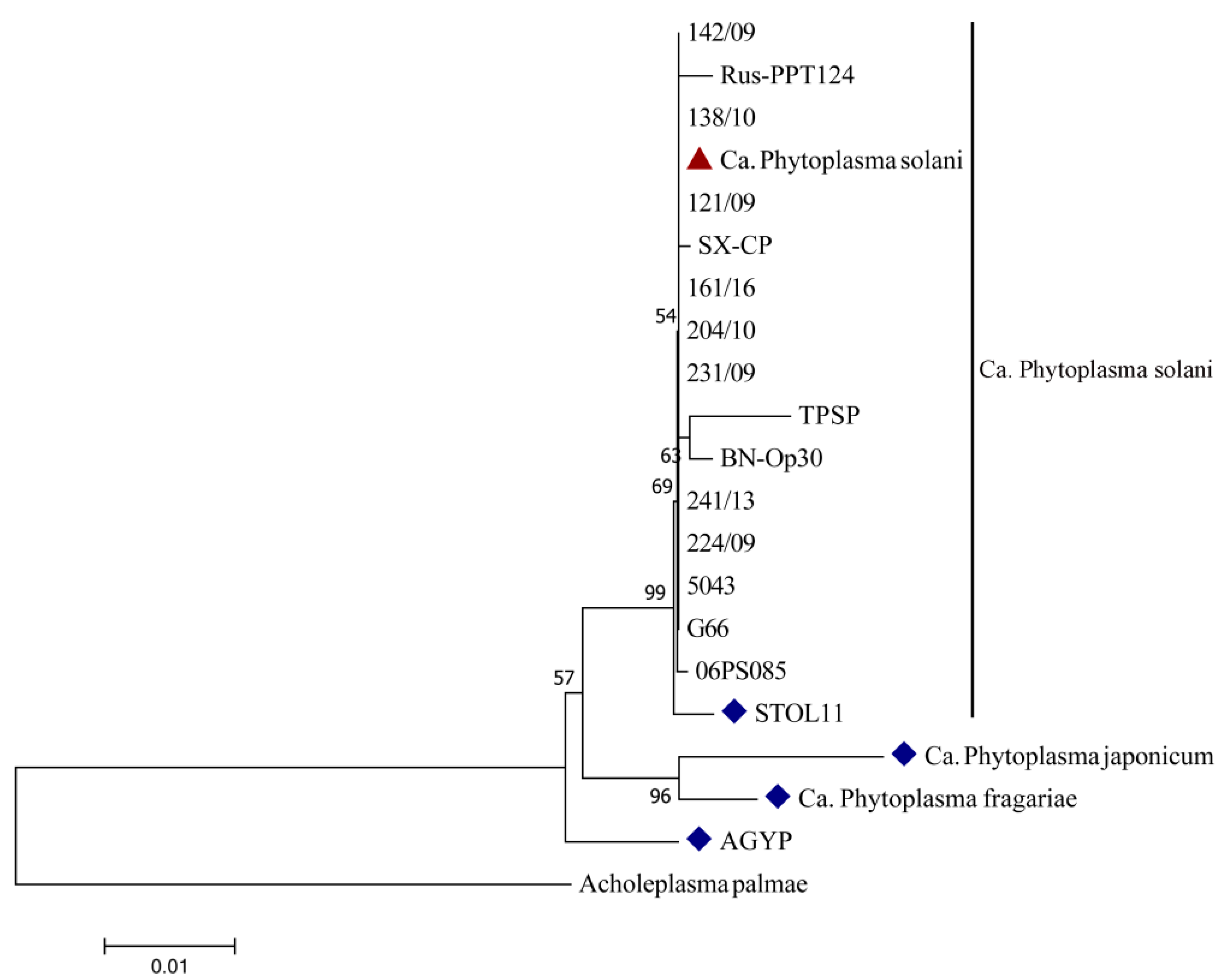
2.2. Molecular Identification of Candidatus Phytoplasma Solani
2.3. Oxidative Stress and Antioxidative Activity
3. Discussion
4. Materials and Methods
4.1. Symptoms Observation and Sampling of Plant Material
4.2. Molecular Identification of Candidatus Phytoplasma Solani
4.3. Determination of Lipid Peroxidation Intensity (LP) and Total Carbohydrate Content (TCC)
4.4. Determination of PAL Activity, Total Polyphenols, Flavonoids, and Anthocyanins
4.5. Determination of Chlorophylls and Carotenoids Contents
4.6. Determination of Antioxidant Capacity
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lee, I.M.; Davis, R.E.; Gundersen-Rindal, D.E. Phytoplasma: Phytopathogenic mollicutes. Annu. Rev. Microbiol. 2000, 54, 221–255. [Google Scholar] [CrossRef] [PubMed]
- Ermacora, P.; Osler, R. Symptoms of Phytoplasma Diseases. In Phytoplasmas. Methods in Molecular Biology; Musetti, R., Pagliari, L., Eds.; Humana Press: New York, NY, USA, 2019; Volume 1875, pp. 53–67. [Google Scholar] [CrossRef]
- Kumari, S.; Krishnan, N.; Rai, A.B.; Singh, B.; Rao, G.P.; Bertaccini, A. Global status of phytoplasma diseases in vegetable crops. Front. Microbiol. 2019, 10, 1349. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox Signaling in Plants. Antioxid. Redox Signal 2013, 18, 2087–2090. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. Intervention of phytohormone pathways by pathogen effectors. Plant Cell 2014, 26, 2285–2309. [Google Scholar] [CrossRef] [PubMed]
- Tomkins, M.; Kliot, A.; Marée, A.F.; Hogenhout, S.A. A multi-layered mechanistic modelling approach to understand how effector genes extend beyond phytoplasma to modulate plant hosts, insect vectors and the environment. Curr. Opin. Plant Biol. 2018, 44, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Musetti, R. Biochemical changes in plants infected by phytoplasmas. In Phytoplasmas: Genomes, Plant Hosts and Vectors; Weintraub, P.G., Jones, P., Eds.; CAB International: Wallingford, UK, 2010; pp. 132–146. [Google Scholar] [CrossRef]
- Kiprovski, B.; Đalović, I.; Adamović, D.; Mitrović, P.; Marjanović-Jeromela, A.; Malenčić, Đ.; Popović, T. Biochemical changes in Oenothera biennis plants infected by ‘Candidatus Phytoplasma solani’. J. Plant Pathol. 2018, 100, 209–214. [Google Scholar] [CrossRef]
- Malamy, J.; Sanchez-Casas, P.; Hennig, J.; Guo, A.; Klessig, D.F. Dissection of the salicylic acid signaling pathway in tobacco. Mol. Plant Microbe Interact. MPMI 1996, 9, 474–482. [Google Scholar] [CrossRef]
- Mur, L.A.J.; Bi, Y.M.; Darby, R.M.; Firek, S.; Draper, J. Compromising early salicylic acid accumulation delays the hypersensitive response and increases viral dispersal during lesion establishment in TMV-infected tobacco. Plant J. 1997, 12, 1113–1126. [Google Scholar] [CrossRef] [PubMed]
- Shirasu, K.; Nakajima, H.; Rajasekhar, V.K.; Dixon, R.A.; Lamb, C. Salicylic acid potentiates an agonist-dependent gain control that amplifies pathogen signals in the activation of defense mechanisms. Plant Cell 1997, 9, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, D.M.A.; Shah, J.; Klessig, D.F. Salicylic acid and disease resistance in plants. Crit. Rev. Plant Sci. 1999, 18, 547–575. [Google Scholar] [CrossRef]
- Felton, G.W.; Korth, K.L.; Bi, J.L.; Wesley, S.V.; Huhman, D.V.; Mathews, M.C.; Murphy, J.B.; Lamb, C.; Dixon, R.A. Inverse relationship between systemic resistance of plants to microorganisms and to insect herbivory. Curr. Biol. 1999, 9, 317–320. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Bolwell, P.G.; Bramley, P.M.; Pridham, J.B. The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 1995, 22, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef]
- Jović, J.; Cvrković, T.; Mitrović, M.; Krnjajić, S.; Petrović, A.; Redinbaugh, G.M.; Pratt, C.R.; Hogenhout, A.S.; Toševski, I. Stolbur Phytoplasma transmission to maize by Reptalus panzeri and the disease cycle of maize redness in Serbia. Phytopathology 2009, 99, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Adamović, D.; Đalović, I.; Mitrović, P.; Kojić, S.; Starović, M.; Purar, B.; Jošić, D. First report of 16SrXII-A subgroup Phytoplasma (Stolbur) associated with reddening of Oenothera biennis in Serbia. Plant Dis. 2014, 98, 841. [Google Scholar] [CrossRef] [PubMed]
- Starović, M.; Pavlović, S.; Stojanović, S.; Jošić, D. Medicinal plants phytoplasma. Plant Prot. 2015, 66, 7–31. (In Serbian) [Google Scholar] [CrossRef]
- Mitrović, P.; Trkulja, V.; Adamović, D.; Đalović, I.; Milovac, Ž.; Kovačić–Jošić, D.; Mihić–Salapura, J. First report of Stolbur Phytoplasma on Mentha x piperita in Serbia. Plant Dis. 2016, 100, 853. [Google Scholar] [CrossRef]
- Duduk, B.; Bulajić, A.; Duduk, N.; Calari, A.; Paltrinieri, S.; Krstić, B.; Bertaccini, A. Identification of phytoplasmas belonging to aster yellows ribosomal group in vegetables in Serbia. Bull. Insectology 2007, 60, 341. [Google Scholar]
- Duduk, B.; Perić, P.; Marčić, D.; Drobnjaković, T.; Picciau, L.; Alma, A.; Bertaccini, A. Phytoplasmas in carrots disease and potential vectors in Serbia. Bull. Insectol. 2008, 61, 327–331. [Google Scholar]
- Martinović, M.; Bjegović, P. O nekim bolestima i štetočinama utvrđenim u NR Srbiji u 1949 godini. Plant Protect. 1950, 2, 59–68. [Google Scholar]
- Duduk, B.; Bertaccini, A. Corn with symptoms of reddening: New host of stolbur phytoplasma. Plant Dis. 2006, 90, 1313–1319. [Google Scholar] [CrossRef]
- Mitrović, J.; Duduk, B. Occurrence of a new stolbur strain in tobacco in Serbia. Bull. Insectol. 2011, 64, S107–S108. [Google Scholar]
- Medić Pap, S.; Gvozdanović Varga, J.; Červenski, J.; Stepanović, J.; Rekanović, E.; Stepanović, M.; Duduk, B. First Report of ‘Candidatus Phytoplasma solani’Infecting Parsnip in Serbia. Plant Dis. 2018, 102, 1026. [Google Scholar] [CrossRef]
- Duduk, B.; Botti, S.; Ivanović, M.; Krstić, B.; Dukić, N.; Bertaccini, A. Identification of phytoplasmas associated with grapevine yellows in Serbia. J. Phytopathol. 2004, 152, 575–579. [Google Scholar] [CrossRef]
- Starović, M.; Kojić, S.; Kuzmanović, S.T.; Stojanović, S.D.; Pavlović, S.; Jošić, D. First Report of Blueberry Reddening Disease in Serbia Associated with 16SrXIIA (Stolbur) Phytoplasma. Plant Dis. 2013, 97, 1653. [Google Scholar] [CrossRef]
- Mitrović, J.; Pavlović, S.; Duduk, B. Survey and multigene characterization of stolbur phytoplasmas on various plant species in Serbia. Phytopathol. Mediterr. 2013, 434–441. [Google Scholar] [CrossRef]
- Maixner, M. Recent advances in Bois noir research. Petria 2011, 21, 85–190. [Google Scholar]
- Mehle, N.; Mermal, S.; Vidmar, S.; Marn, M.V.; Dreo, T.; Dermastia, M. First Report of Carrot Infection with Phytoplasmas in Slovenia. In V: Bois Noir 5th Workshop; City Hotel: Ljubljana, Slovenia, 2018; Volume 18, pp. 1–4. [Google Scholar]
- Langer, M.; Maixner, M. Molecular characterisation of grapevine yellows associated phytoplasmas of the stolbur-group based on RFLP–analysis of non-ribosomal DNA. Vitis 2004, 43, 191–199. [Google Scholar] [CrossRef]
- Cimerman, A.; Pacifico, D.; Salar, P.; Marzachì, C.; Foissac, X. Striking diversity of vmp1, a variable gene encoding a putative membrane protein of the stolbur phytoplasma. Appl. Environ. Microbiol. 2009, 75, 2951–2957. [Google Scholar] [CrossRef] [PubMed]
- Quaglino, F.; Zhao, Y.; Casati, P.; Bulgari, D.; Bianco, P.A.; Wei, W.; Davis, R.E. ‘Candidatus Phytoplasma solani’, a novel taxon associated with stolbur-and bois noir-related diseases of plants. Int. J. Syst. Evol. Microbiol. 2013, 63, 2879–2894. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Oshima, K.; Ammar, E.D.; Kakizawa, S.; Kingdom, H.N.; Namba, S. Phytoplasmas: Bacteria that manipulate plants and insects. Mol. Plant Pathol. 2008, 9, 403–423. [Google Scholar] [CrossRef]
- Himeno, M.; Kitazawa, Y.; Yoshida, T.; Maejima, K.; Yamaji, Y.; Oshima, K.; Namba, S. Purple top symptoms are associated with reduction of leaf cell death in phytoplasma-infected plants. Sci. Rep. 2014, 4, 4111. [Google Scholar] [CrossRef] [PubMed]
- Matros, A.; Peshev, D.; Peukert, M.; Mock, H.P.; van den Ende, W. Sugars as hydroxyl radical scavengers: Proof-of-concept by studying the fate of sucralose in Arabidopsis. Plant J. 2015, 82, 822–839. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Mock, R.; Huang, Q.; Abad, J.; Hartung, J.; Kinard, G. A reliable and inexpensive method of nucleic acid extraction for the PCR-based detection of diverse plant pathogens. J. Virol. Methods 2008, 154, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Hiruki, C. Amplification of 16S rRNA genes from culturable and nonculturable mollicutes. J. Microbiol. Methods. 1991, 14, 53–61. [Google Scholar] [CrossRef]
- Schneider, B.; Seemuller, E.; Smart, C.D.; Kirkpatrick, B.C. Phylogenetic classification of plant pathogenetic mycoplasmalike organisms or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmology; Razin, S., Tully, J.G., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 369–380. [Google Scholar]
- Lee, M.; Martini, M.; Marcone, C.; Zhu, S.F. Classification of phytoplasma strains in the elm yellows group (16SrV) and proposal of ‘Candidatus Phytoplasma ulmi’ for the phytoplasma associated with elm yellows. Int. J. Syst. Evol. Microbiol. 2004, 54, 337–347. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Albalasmeh, A.A.; Berhe, A.A.; Ghezzehei, T.A. A new method for rapid determination of carbohydrate and total carbon concentrations using UV spectrophotometry. Carbohydr. Polym. 2013, 97, 253–261. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159. [Google Scholar] [CrossRef]
- Gerasimova, N.G.; Pridvorova, S.M.; Ozeretskovskaya, O.L. Role of L–phenylalanine ammonia Lyase in the induced resistance and susceptibility of sotato plants. Appl. Biochem. Microbiol. 2005, 41, 103–105. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Quantification of Tannins in Tree and Shrub Foliage: A Laboratory Manual; Springer Science & Business Media: Berlin, Germany, 2003; p. 102. [Google Scholar] [CrossRef]
- Pękal, A.; Pyrzynska, K. Evaluation of aluminium complexation reaction for flavonoid content assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef]
- Lee, J.; Durst, R.W.; Wrolstad, R.E. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef]
- Von Wettstein, D. Chlorophyll-letale und der submikroskopische Formwechsel der plastiden. Exp. Cell Res. 1957, 12, 427–506. [Google Scholar] [CrossRef]
- De Carvalho, L.M.J.; Gomes, P.B.; de Oliveira Godoy, R.L.; Pacheco, S.; do Monte, P.H.F.; de Carvalho, J.L.V.; Nutti, M.R.; Neves, A.C.L.; Vieira, A.; Ramos, S.R.R. Total carotenoid content, α-carotene and β-carotene, of landrace pumpkins (Cucurbita moschata Duch): A preliminary study. Food Res. Int. 2012, 47, 337–340. [Google Scholar] [CrossRef]
- Panda, S.K. Assay guided comparison for enzymatic andnonenzymatic antioxidant activities with special reference to medicinal plants. In Antioxidant Enzyme, 3rd ed.; El-Missiry, M.A., Ed.; IntechOpen: Rijeka, Croatia, 2012; Volume 14, pp. 382–400. [Google Scholar] [CrossRef]
- Ahmed, D.; Saman, Z.; Hira, B. In vitro analysis of antioxidant activities of Oxalis corniculata Linn. fractions in various solvents. Afr. J. Tradit. Complement Altern. Med. 2013, 10, 158–165. [Google Scholar] [CrossRef][Green Version]
- Sánchez-Moreno, C. Methods used to evaluate the free radical scavenging activity in foods and biological systems. Int. J. Food Sci. Technol. 2002, 8, 121–137. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Host | Country | Acc. Number |
|---|---|---|---|
| Ca. Phytoplasma solani | |||
| 121/09 | Corn | Serbia | JQ730750 |
| 142/09 | Tobacco | Serbia | JQ730739 |
| 224/09 | Valerian | Serbia | JQ730742 |
| 231/09 | Parsley | Serbia | JQ730741 |
| 138/10 | Grapevine | Serbia | JQ730746 |
| 204/10 | Periwinkle | Serbia | JQ730744 |
| 161/16 | Parsnip | Serbia | KY579338 |
| STOL11 (STOL) * | Periwinkle | France | AF248959 |
| 241/13 | Corn | Bulgaria | KF907506 |
| 5043 | Tomato | Greece | JX311953 |
| G66 | Pea | Poland | JN887313 |
| SX-CP | Red sage | China | KT844645 |
| 06PS085 | Grapevine | Canada | EU086529 |
| Rus-PPT124 a | Potato | Russia | EU344890 |
| TPSP b | Potato | Turkey | HM485579 |
| BN-Op30 c | Grapevine | Italy | EU836652 |
| Ca. Phytoplasma australiense | |||
| AGYP * | Grapevine | Australia | L76865 |
| Ca. Phytoplasma japonicum * | Hydrangea | Japan | AB010425 |
| Ca. Phytoplasma fragariae * | Strawberry | Lithuania | DQ086423 |
| Acholeplasma palmae | L33734 |
| 2016 | 2017 | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatments | Mean | St. Error | A/S | Mean | St. Error | A/S | 2016/2017 | |
| Total sugars | AL | 5.79 | 0.01 | * | 6.99 | 0.52 | * | * |
| SL | 5.62 | 0.00 | 16.61 | 0.59 | * | |||
| AR | 4.10 | 0.01 | * | 12.94 | 1.48 | * | * | |
| SR | 5.15 | 0.01 | 16.03 | 0.75 | * | |||
| Reduced glutathione | AL | 2.75 | 0.01 | * | 6.94 | 0.05 | * | * |
| SL | 5.49 | 0.01 | 17.88 | 0.11 | * | |||
| AR | 3.10 | 0.01 | * | 2.56 | 0.05 | * | * | |
| SR | 3.29 | 0.01 | 8.92 | 0.19 | * | |||
| Lipid peroxidation intensity | AL | 871.57 | 0.26 | * | 70.00 | 3.70 | * | * |
| SL | 1360.13 | 0.47 | 198.71 | 6.63 | * | |||
| AR | 1064.43 | 3.80 | * | 108.84 | 6.17 | * | * | |
| SR | 1264.00 | 3.06 | 188.32 | 9.03 | * | |||
| 2016 | 2017 | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Mean | St. Error | A/S | Mean | St. Error | A/S | 2016/2017 | |
| Phenylalanine ammonia-lyase activity | AL | 258.82 | 0.29 | nd | 281.23 | 3.33 | * | nd |
| SL | 336.71 | 11.00 | 518.32 | 12.42 | * | |||
| AR | 70.94 | 0.03 | nd | 80.22 | 2.12 | nd | nd | |
| SR | 84.85 | 0.11 | 100.45 | 4.70 | nd | |||
| Total polyphenols | AL | 4.03 | 0.02 | * | 5.57 | 0.03 | nd | nd |
| SL | 6.88 | 0.02 | 5.47 | 0.01 | nd | |||
| AR | 1.42 | 0.01 | * | 1.13 | 0.01 | nd | nd | |
| SR | 1.92 | 0.01 | 1.34 | 0.00 | nd | |||
| Total tannins | AL | 3.33 | 0.04 | * | 2.32 | 0.28 | nd | nd |
| SL | 3.94 | 0.01 | 2.70 | 0.09 | nd | |||
| AR | 1.18 | 0.03 | * | 0.87 | 0.12 | nd | nd | |
| SR | 0.76 | 0.02 | 1.12 | 0.10 | nd | |||
| Total flavonoids | AL | 0.043 | 0.002 | * | 0.325 | 0.066 | nd | * |
| SL | 0.100 | 0.006 | 0.235 | 0.078 | * | |||
| AR | 0.000 | 0.000 | nd | 0.033 | 0.038 | nd | * | |
| SR | 0.000 | 0.000 | 0.025 | 0.075 | * | |||
| Total proanthocyanidins | AL | 0.40 | 0.00 | * | 0.003 | 0.00 | * | * |
| SL | 0.15 | 0.00 | 0.001 | 0.00 | * | |||
| AR | 6.02 | 0.00 | * | 0.001 | 0.00 | * | * | |
| SR | 4.87 | 0.01 | 0.002 | 0.00 | * | |||
| Total anthocyanins | AL | 0.003 | 0.00 | * | 0.088 | 0.00 | * | * |
| SL | 0.798 | 0.00 | 0.025 | 0.00 | * | |||
| AR | 0.00 | 0.00 | * | 0.00 | 0.00 | nd | nd | |
| SR | 0.00 | 0.00 | 0.00 | 0.00 | nd | |||
| 2016 | 2017 | |||||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Mean | St. Error | A/S | Mean | St. Error | A/S | 2016/2017 | |
| Total chlorophyll a | AL | 0.78 | 0.01 | * | 0.25 | 0.08 | * | * |
| SL | 0.24 | 0.01 | 0.10 | 0.02 | * | |||
| AR | 0.13 | 0.01 | nd | 0.00 | 0.00 | nd | nd | |
| SR | 0.15 | 0.01 | 0.00 | 0.00 | nd | |||
| Total chlorophyll b | AL | 0.18 | 0.00 | * | 0.15 | 0.09 | * | nd |
| SL | 0.15 | 0.00 | 0.06 | 0.04 | * | |||
| AR | 0.25 | 0.00 | nd | 0.00 | 0.00 | nd | * | |
| SR | 0.28 | 0.01 | 0.00 | 0.00 | * | |||
| Total carotenoids | AL | 0.36 | 0.01 | * | 0.009 | 0.02 | * | * |
| SL | 0.17 | 0.00 | 0.002 | 0.02 | * | |||
| AR | 1.94 | 0.00 | nd | 0.200 | 0.02 | nd | nd | |
| SR | 1.59 | 0.00 | 0.170 | 0.05 | * | |||
| 2016 | 2017 | |||||||
|---|---|---|---|---|---|---|---|---|
| Antioxidant test | Treatment | Mean | St. Error | A/S | Mean | St. Error | A/S | 2016/2017 |
| NBT-test | AL | 20.33 | 0.28 | nd | 55.89 | 0.10 | nd | * |
| SL | 19.80 | 0.06 | 54.93 | 0.02 | * | |||
| AR | 82.57 | 0.23 | * | 67.94 | 0.02 | nd | * | |
| SR | 99.23 | 0.09 | 80.95 | 0.00 | nd | |||
| •OH-test | AL | 10.81 | 0.69 | * | 69.68 | 0.06 | nd | * |
| SL | 20.32 | 1.93 | 55.98 | 0.01 | * | |||
| AR | 2.42 | 0.04 | * | 11.54 | 0.01 | nd | * | |
| AL | 39.97 | 4.16 | 13.75 | 0.00 | * | |||
| DPPH-test | SL | 19.95 | 0.02 | * | 55.99 | 0.15 | nd | * |
| AR | 32.58 | 0.22 | 48.09 | 1.17 | * | |||
| SR | 10.90 | 0.01 | * | 19.52 | 1.00 | nd | * | |
| SR | 21.25 | 0.01 | 18.49 | 1.47 | nd | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitrovic, P.; Djalovic, I.; Kiprovski, B.; Veljović Jovanović, S.; Trkulja, V.; Jelušić, A.; Popović, T. Oxidative Stress and Antioxidative Activity in Leaves and Roots of Carrot Plants Induced by Candidatus Phytoplasma Solani. Plants 2021, 10, 337. https://doi.org/10.3390/plants10020337
Mitrovic P, Djalovic I, Kiprovski B, Veljović Jovanović S, Trkulja V, Jelušić A, Popović T. Oxidative Stress and Antioxidative Activity in Leaves and Roots of Carrot Plants Induced by Candidatus Phytoplasma Solani. Plants. 2021; 10(2):337. https://doi.org/10.3390/plants10020337
Chicago/Turabian StyleMitrovic, Petar, Ivica Djalovic, Biljana Kiprovski, Sonja Veljović Jovanović, Vojislav Trkulja, Aleksandra Jelušić, and Tatjana Popović. 2021. "Oxidative Stress and Antioxidative Activity in Leaves and Roots of Carrot Plants Induced by Candidatus Phytoplasma Solani" Plants 10, no. 2: 337. https://doi.org/10.3390/plants10020337
APA StyleMitrovic, P., Djalovic, I., Kiprovski, B., Veljović Jovanović, S., Trkulja, V., Jelušić, A., & Popović, T. (2021). Oxidative Stress and Antioxidative Activity in Leaves and Roots of Carrot Plants Induced by Candidatus Phytoplasma Solani. Plants, 10(2), 337. https://doi.org/10.3390/plants10020337