
Analyses of MADS-box Genes Suggest HvMADS56 to Regulate Lateral Spikelet Development in Barley

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Status and Scanning Electron Microscopy (SEM) of the Tested Barley Lines
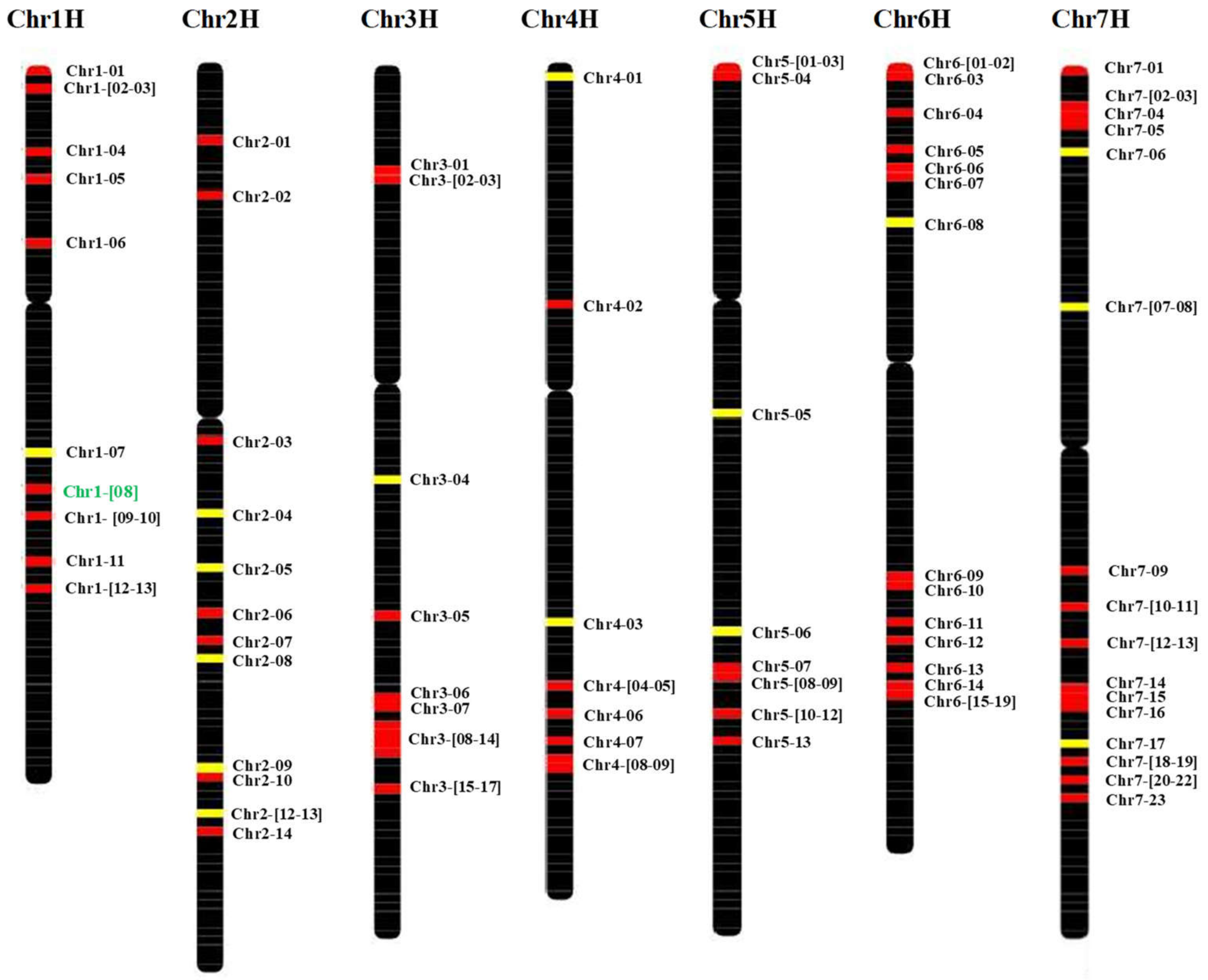
2.2. HvMADS-box Genes Map
2.3. Phylogenetic Analysis of HvMADS-box Genes
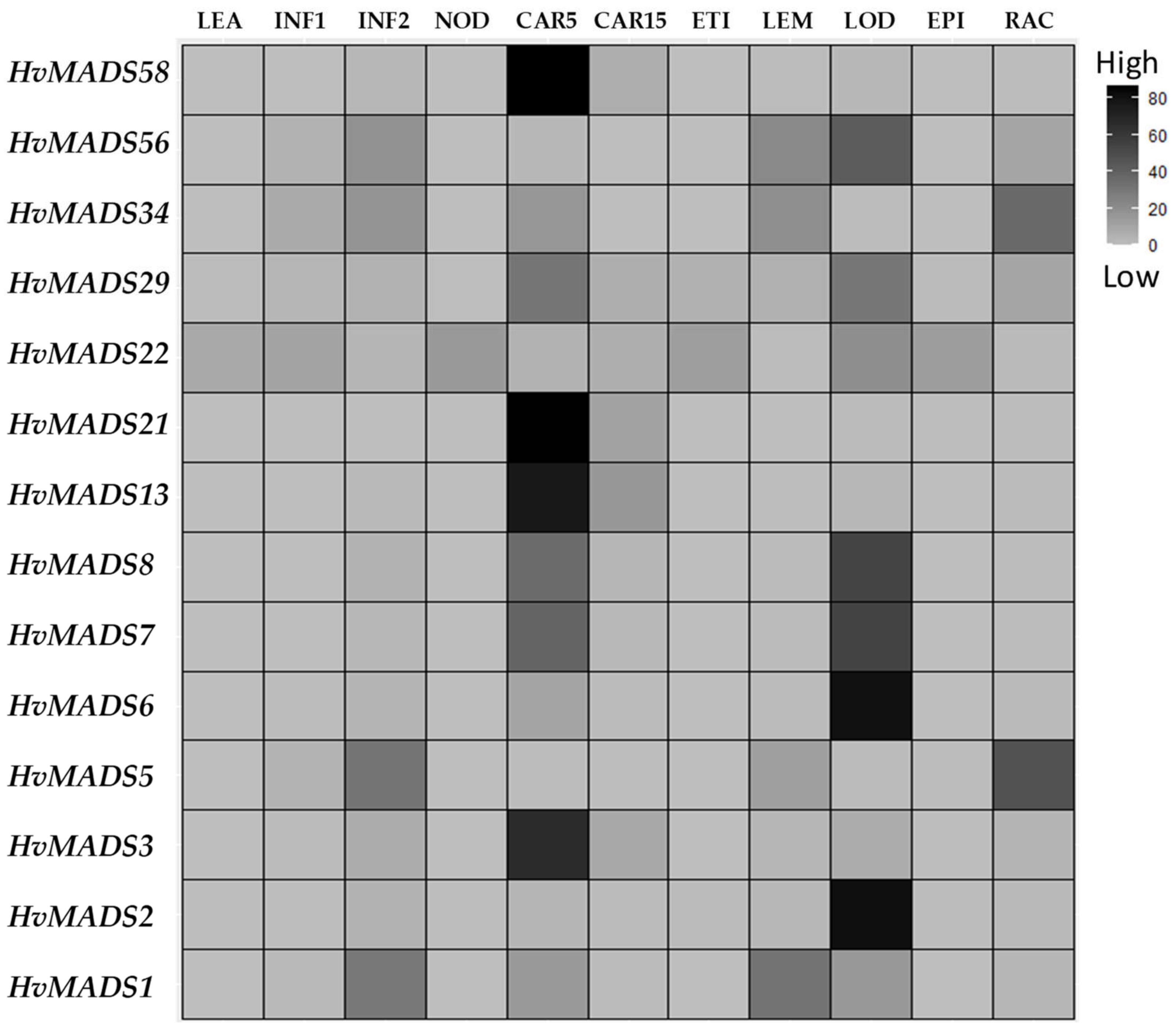
2.4. In Silico Analysis of Barley HvMADS-box Genes Expression
2.5. MADS-box Genes Polymorphism and Lateral Spikelet Development
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials
5.2. Growth Conditions and Spike Phenotyping
5.3. Scanning Electron Microscopy
5.4. Genomic DNA Isolation, Amplification and Sequencing
5.5. Sequence Analysis and Sequence Homology Searches
5.6. Phylogenetic Analysis
5.7. Expression Data Analysis
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donald, C.M. The breeding of crop ideotypes. Euphytica 1968, 17, 385–403. [Google Scholar] [CrossRef]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Plant science: Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Bommert, P.; Nagasawa, N.S.; Jackson, D. Quantitative variation in maize kernel row number is controlled by the FASCIATED EAR2 locus. Nat. Genet. 2013, 45, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaya, O.; Pont, C.; Sibout, R.; Martinek, P.; Badaeva, E.; Murat, F.; Chosson, A.; Watanabe, N.; Prat, E.; Gautier, N.; et al. Frizzy panicle drives supernumerary spikelets in bread wheat. Plant Physiol. 2015, 167, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Poursarebani, N.; Seidensticker, T.; Koppolu, R.; Trautewig, C.; Gawroński, P.; Bini, F.; Govind, G.; Rutten, T.; Sakuma, S.; Tagiri, A.; et al. The genetic basis of composite spike form in barley and ‘miracle-wheat’. Genetics 2015, 201, 155–165. [Google Scholar] [CrossRef]
- Youssef, H.M.; Mascher, M.; Ayoub, M.A.; Stein, N.; Kilian, B.; Schnurbusch, T. Natural diversity of inflorescence architecture traces cryptic domestication genes in barley (Hordeum vulgare L.). Genet. Resour. Crop Evol. 2017, 64, 843–853. [Google Scholar] [CrossRef]
- Forster, B.P.; Franckowiak, J.D.; Lundqvist, U.; Lyon, J.; Pitkethly, I.; Thomas, W.T.B. The barley phytomer. Ann. Bot. 2007, 100, 725–733. [Google Scholar] [CrossRef] [Green Version]
- Komatsuda, T.; Pourkheirandish, M.; He, C.; Azhaguvel, P.; Kanamori, K.; Perovic, D.; Stein, N.; Graner, A.; Wicker, T.; Tagiri, A.; et al. Six-rowed barley originated from a mutation in a homeodomain-leucine zipper I-class homeobox gene. Proc. Natl. Acad. Sci. USA 2007, 104, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Mansfeld, R. Das morphologische System der Saatgerste, Hordeum vulgare L. sl. Züchter 1950, 64, 843–853. [Google Scholar] [CrossRef]
- Youssef, H.M.; Eggert, K.; Koppolu, R.; Alqudah, A.M.; Poursarebani, N.; Fazeli, A.; Sakuma, S.; Tagiri, A.; Rutten, T.; Govind, G.; et al. VRS2 regulates hormone-mediated inflorescence patterning in barley. Nat. Genet. 2017, 49, 157–161. [Google Scholar] [CrossRef]
- Bull, H.; Casao, M.C.; Zwirek, M.; Flavell, A.J.; Thomas, W.T.B.; Guo, W.; Zhang, R.; Rapazote-Flores, P.; Kyriakidis, S.; Russell, J.; et al. Barley SIX-ROWED SPIKE3 encodes a putative Jumonji C-type H3K9me2/me3 demethylase that represses lateral spikelet fertility. Nat. Commun. 2017, 8, 936. [Google Scholar] [CrossRef] [Green Version]
- Van Esse, G.W.; Walla, A.; Finke, A.; Koornneef, M.; Pecinka, A.; von Korff, M. Six-rowed spike3 (VRS3) is a histone demethylase that controls lateral spikelet development in Barley. Plant Physiol. 2017, 174, 2397–2408. [Google Scholar] [CrossRef] [Green Version]
- Koppolu, R.; Anwar, N.; Sakuma, S.; Tagiri, A.; Lundqvist, U.; Pourkheirandish, M.; Rutten, T.; Seiler, C.; Himmelbach, A.; Ariyadasa, R.; et al. Six-rowed spike4 (Vrs4) controls spikelet determinacy and row-type in barley. Proc. Natl. Acad. Sci. USA 2013, 110, 13198–13203. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, L.; Comadran, J.; Druka, A.; Marshall, D.F.; Thomas, W.T.B.; MacAulay, M.; MacKenzie, K.; Simpson, C.; Fuller, J.; Bonar, N.; et al. INTERMEDIUM-C, a modifier of lateral spikelet fertility in barley, is an ortholog of the maize domestication gene TEOSINTE BRANCHED 1. Nat. Genet. 2011, 43, 169–172. [Google Scholar] [CrossRef]
- Boden, S.A. How hormones regulate floral architecture in barley. Nat. Genet. 2017, 49, 8–9. [Google Scholar] [CrossRef]
- Youssef, H.M.; Hansson, M. Crosstalk among hormones in barley spike contributes to the yield. Plant Cell Rep. 2019, 38, 1013–1016. [Google Scholar] [CrossRef] [Green Version]
- Shore, P.; Sharrocks, A.D. The MADS-Box Family of Transcription Factors. Eur. J. Biochem. 1995, 229, 1–13. [Google Scholar] [CrossRef]
- Lawton-Rauh, A.L.; Alvarez-Buylla, E.R.; Purugganan, M.D. Molecular evolution of flower development. Trends Ecol. Evol. 2000, 15, 144–149. [Google Scholar] [CrossRef]
- Theißen, G.; Saedler, H. Floral quartets. Nature 2001, 409, 469–471. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Hirano, H.Y. Function and diversification of MADS-box genes in rice. Sci. World J. 2006, 6, 1923–1932. [Google Scholar] [CrossRef]
- Callens, C.; Tucker, M.R.; Zhang, D.; Wilson, Z.A. Dissecting the role of MADS-box genes in monocot floral development and diversity. J. Exp. Bot. 2018, 69, 2435–2459. [Google Scholar] [CrossRef]
- Yoshida, H.; Nagato, Y. Flower development in rice. J. Exp. Bot. 2011, 62, 4719–4730. [Google Scholar] [CrossRef] [Green Version]
- Kuijer, H.N.J.; Shirley, N.J.; Khor, S.F.; Shi, J.; Schwerdt, J.; Zhang, D.; Li, G.; Burton, R.A. Transcript Profiling of MIKCc MADS-Box Genes Reveals Conserved and Novel Roles in Barley Inflorescence Development. Front. Plant Sci. 2021, 12, 1834. [Google Scholar] [CrossRef]
- Kirby, E.J.M.; Appleyard, M. Cereal Development Guide, 2nd ed.; NAC Cereal Unit: Stoneleigh, UK, 1987; 85p. [Google Scholar]
- Colmsee, C.; Beier, S.; Himmelbach, A.; Schmutzer, T.; Stein, N.; Scholz, U.; Mascher, M. BARLEX—The barley draft genome explorer. Mol. Plant 2015, 8, 964–966. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, S.; Lundqvist, U.; Kakei, Y.; Thirulogachandar, V.; Suzuki, T.; Hori, K.; Wu, J.; Tagiri, A.; Rutten, T.; Koppolu, R.; et al. Extreme suppression of lateral floret development by a single amino acid change in the VRS1 transcription factor. Plant Physiol. 2017, 175, 1720–1731. [Google Scholar] [CrossRef] [Green Version]
- Schwarz-Sommer, Z.; Huijser, P.; Nacken, W.; Saedler, H.; Sommer, H. Genetic control of flower development by homeotic genes in Antirrhinum majus. Science 1990, 250, 931–936. [Google Scholar] [CrossRef] [Green Version]
- Pelucchi, N.; Fornara, F.; Favalli, C.; Masiero, S.; Lago, C.; Pè, E.M.; Colombo, L.; Kater, M.M. Comparative analysis of rice MADS-box genes expressed during flower development. Sex. Plant Reprod. 2002, 15, 113–122. [Google Scholar] [CrossRef]
- Becker, A.; Theißen, G. The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phylogenet. Evol. 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, G.; Ren, X.; Du, B.; Cheng, Y.; Wang, Y.; Li, C.; Sun, D. Dissecting the genetic basis of grain size and weight in barley (Hordeum vulgare L.) by QTL and comparative genetic analyses. Front. Plant Sci. 2019, 10, 469. [Google Scholar] [CrossRef]
- Youssef, H.M.; Allam, M.; Boussora, F.; Himmelbach, A.; Milner, S.G.; Mascher, M.; Schnurbusch, T. Dissecting the genetic basis of lateral and central spikelet development and grain traits in intermedium-spike barley (Hordeum vulgare convar. intermedium). Plants 2020, 9, 1655. [Google Scholar] [CrossRef]
- Schilling, S.; Kennedy, A.; Pan, S.; Jermiin, L.S.; Melzer, R. Genome-wide analysis of MIKC-type MADS-box genes in wheat: Pervasive duplications, functional conservation and putative neofunctionalization. New Phytol. 2020, 225, 511–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciaffi, M.; Paolacci, A.R.; Tanzarella, O.A.; Porceddu, E. Molecular aspects of flower development in grasses. Sex. Plant Reprod. 2011, 24, 247–282. [Google Scholar] [CrossRef] [PubMed]
- Bolot, S.; Abrouk, M.; Masood-Quraishi, U.; Stein, N.; Messing, J.; Feuillet, C.; Salse, J. The ‘inner circle’ of the cereal genomes. Curr. Opin. Plant Biol. 2009, 12, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Mayer, K.F.X.; Martis, M.; Hedley, P.E.; Šimková, H.; Liu, H.; Morris, J.A.; Steuernagel, B.; Taudien, S.; Roessner, S.; Gundlach, H.; et al. Unlocking the barley genome by chromosomal and comparative genomics. Plant Cell 2011, 23, 1249–1263. [Google Scholar] [CrossRef] [Green Version]
- Wicker, T.; Mayer, K.F.X.; Gundlach, H.; Martis, M.; Steuernagel, B.; Scholz, U.; Šimková, H.; Kubaláková, M.; Choulet, F.; Taudien, S.; et al. Frequent gene movement and pseudogene evolution is common to the large and complex genomes of wheat, barley, and their relatives. Plant Cell 2011, 23, 1706–1718. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, K.; Melzer, R.; Theißen, G. MIKC-type MADS-domain proteins: Structural modularity, protein interactions and network evolution in land plants. Gene 2005, 347, 183–198. [Google Scholar] [CrossRef]
- Samach, A.; Onouchi, H.; Gold, S.E.; Ditta, G.S.; Schwarz-Sommer, Z.; Yanofsky, M.F.; Coupland, G. Distinct roles of constans target genes in reproductive development of Arabidopsis. Science 2000, 288, 1613–1616. [Google Scholar] [CrossRef] [Green Version]
- Papaefthimiou, D.; Kapazoglou, A.; Tsaftaris, A.S. Cloning and characterization of SOC1 homologs in barley (Hordeum vulgare) and their expression during seed development and in response to vernalization. Physiol. Plant. 2012, 146, 71–85. [Google Scholar] [CrossRef]
- Lee, H.; Suh, S.S.; Park, E.; Cho, E.; Ahn, J.H.; Kim, S.G.; Lee, J.S.; Kwon, Y.M.; Lee, I. The AGAMOUS-lIKE 20 MADS domain protein integrates floral inductive pathways in Arabidopsis. Genes Dev. 2000, 14, 2366–2376. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Kim, J.; Han, J.J.; Han, M.J.; An, G. Functional analyses of the flowering time gene OsMADS50, the putative Suppressor of Overexpression of CO1/Agamous-Like 20 (SOC1/AGL20) ortholog in rice. Plant J. 2004, 38, 754–764. [Google Scholar] [CrossRef]
- Lee, J.; Oh, M.; Park, H.; Lee, I. SOC1 translocated to the nucleus by interaction with AGL24 directly regulates LEAFY. Plant J. 2008, 55, 832–843. [Google Scholar] [CrossRef]
- Cseke, L.J.; Zheng, J.; Podila, G.K. Characterization of PTM5 in aspen trees: A MADS-box gene expressed during woody vascular development. Gene 2003, 318, 55–67. [Google Scholar] [CrossRef]
- Ferrario, S.; Busscher, J.; Franken, J.; Gerats, T.; Vandenbussche, M.; Angenent, G.C.; Immink, R.G.H. Ectopic expression of the petunia MADS box gene UNSHAVEN accelerates flowering and confers leaf-like characteristics to floral organs in a dominant-negative manner. Plant Cell 2004, 16, 1490–1505. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Song, I.J.; Fukuda, T.; Yokoyama, J.; Maki, M.; Ochiai, T.; Kameya, T.; Kanno, A. Characterization of TrcMADS1 gene of Trillium camtschatcense (Trilliaceae) reveals functional evolution of the SOC1/TM3-like gene family. J. Plant Res. 2005, 118, 229–234. [Google Scholar] [CrossRef]
- Liu, C.; Zhou, J.; Bracha-Drori, K.; Yalovsky, S.; Ito, T.; Yu, H. Specification of Arabidopsis floral meristem identity by repression of flowering time genes. Development 2007, 134, 1901–1910. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Xi, W.; Shen, L.; Tan, C.; Yu, H. Regulation of Floral Patterning by Flowering Time Genes. Dev. Cell 2009, 16, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Melzer, S.; Lens, F.; Gennen, J.; Vanneste, S.; Rohde, A.; Beeckman, T. Flowering-time genes modulate meristem determinacy and growth form in Arabidopsis thaliana. Nat. Genet. 2008, 40, 1489–1492. [Google Scholar] [CrossRef] [Green Version]
- Castelán-Muñoz, N.; Herrera, J.; Cajero-Sánchez, W.; Arrizubieta, M.; Trejo, C.; García-Ponce, B.; Sánchez, M.; Álvarez-Buylla, E.R.; Garay-Arroyo, A. MADS-Box Genes Are Key Components of Genetic Regulatory Networks Involved in Abiotic Stress and Plastic Developmental Responses in Plants. Front. Plant Sci. 2019, 10, 853. [Google Scholar] [CrossRef] [Green Version]
- Teo, Z.W.N.; Zhou, W.; Shen, L. Dissecting the Function of MADS-Box Transcription Factors in Orchid Reproductive Development. Front. Plant Sci. 2019, 10, 1474. [Google Scholar] [CrossRef]
- Lolas, I.B.; Himanen, K.; Grønlund, J.T.; Lynggaard, C.; Houben, A.; Melzer, M.; Lijsebettens, M.V.; Grasser, K.D. The transcript elongation factor FACT affects Arabidopsis vegetative and reproductive development and genetically interacts with HUB1/2. Plant J. 2010, 61, 686–697. [Google Scholar] [CrossRef]
- Matyszczak, I.; Tominska, M.; Zakhrabekova, S.; Dockter, C.; Hansson, M. Analysis of early-flowering genes at barley chromosome 2H expands the repertoire of mutant alleles at the Mat-c locus. Plant Cell Rep. 2020, 39, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Mascher, M.; Richmond, T.A.; Gerhardt, D.J.; Himmelbach, A.; Clissold, L.; Sampath, D.; Ayling, S.; Steuernagel, B.; Pfeifer, M.; D’Ascenzo, M.; et al. Barley whole exome capture: A tool for genomic research in the genus Hordeum and beyond. Plant J. 2013, 76, 494–505. [Google Scholar] [CrossRef] [Green Version]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Anand, L.; Rodriguez Lopez, C.M. ChromoMap: An R package for Interactive Visualization and Annotation of Chromosomes. bioRxiv 2019, 605600. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.S. Using DECIPHER v2.0 to analyze big biological sequence data in R. R J. 2016, 8, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- SAS Institute Inc. The SAS System for Windows, Release 9.0; SAS Institute: Cary, NC, USA, 2002. [Google Scholar]
- Padi, F.K. Genotype × environment interaction and yield stability in a cowpea-based cropping system. Euphytica 2007, 158, 11–25. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Squares | |||||||
|---|---|---|---|---|---|---|---|
| LSW (g) | LSWi (mm) | LSL (mm) | LSA (mm2) | TSW (g) | |||
| Means of Tested Lines | |||||||
| Two-rowed | Bonus | 0.0004 | 1.37 | 9.22 | 7.20 | 0.37 | |
| Bowman | 0.0005 | 1.69 | 8.49 | 6.61 | 0.54 | ||
| Foma | 0.0005 | 1.62 | 9.37 | 7.63 | 0.48 | ||
| Kristina | 0.0002 | 0.98 | 6.87 | 3.79 | 0.19 | ||
| Average | 0.0004 | 1.42 | 8.49 | 6.31 | 0.40 | ||
| Deficiens | def.2 | 0.0001 | 0.54 | 3.46 | 0.97 | 0.07 | |
| def.4 | 0.0001 | 0.64 | 5.12 | 1.52 | 0.09 | ||
| def.5 | 0.0001 | 0.88 | 6.46 | 2.49 | 0.14 | ||
| def.6 | 0.0002 | 1.09 | 5.12 | 1.75 | 0.17 | ||
| def.8 | 0.0002 | 1.00 | 6.4 | 2.80 | 0.16 | ||
| Average | 0.0001 | 0.83 | 5.31 | 1.91 | 0.13 | ||
| Semi-deficiens | semi-def.1 | 0.0001 | 0.89 | 6.75 | 3.20 | 0.15 | |
| semi-def.4 | 0.0002 | 0.97 | 5.86 | 3.00 | 0.17 | ||
| semi-def.5 | 0.0001 | 0.87 | 6.58 | 2.56 | 0.15 | ||
| semi-def.7 | 0.0002 | 0.91 | 7.38 | 3.17 | 0.17 | ||
| semi-def.8 | 0.0002 | 0.92 | 6.79 | 3.09 | 0.18 | ||
| Average | 0.0002 | 0.91 | 6.67 | 3.00 | 0.16 | ||
| Intermedium mutants | int-a.1 | 0.0014 | 1.81 | 11.22 | 10.99 | 1.39 | |
| int-c.5 | 0.0029 | 2.28 | 10.18 | 13.99 | 2.9 | ||
| int-e.20 | 0.0007 | 1.44 | 9.67 | 8.34 | 0.72 | ||
| int-e.4 | 0.0026 | 2.15 | 10.41 | 12.47 | 2.59 | ||
| int-f.19 | 0.0009 | 1.45 | 12.58 | 9.28 | 0.94 | ||
| int-h.43 | 0.0003 | 1.33 | 6.94 | 4.76 | 0.32 | ||
| int-i.39 | 0.0004 | 1.13 | 8.51 | 5.63 | 0.41 | ||
| int-l.81 | 0.002 | 2.23 | 10.71 | 12.81 | 2.02 | ||
| int-m.85 | 0.0007 | 1.81 | 10.18 | 8.42 | 0.7 | ||
| Average | 0.00132 | 1.74 | 10.04 | 9.63 | 1.33 | ||
| Intermedium barley | HOR1555 | 0.0416 | 4.12 | 9.77 | 26.84 | 41.56 | |
| HOR6211 | 0.0008 | 1.52 | 9.15 | 8.01 | 0.8 | ||
| HOR7191 | 0.0477 | 4.23 | 10.11 | 27.93 | 47.74 | ||
| HOR10166 | 0.0026 | 1.73 | 12.04 | 13.83 | 2.57 | ||
| HOR6407 | 0.0007 | 1.19 | 10.13 | 6.79 | 0.75 | ||
| HOR7041 | 0.0227 | 3.43 | 14.55 | 32.13 | 22.71 | ||
| Average | 0.0194 | 2.70 | 10.96 | 19.26 | 19.36 | ||
| Hexastichon mutant | |||||||
| hex-v.3 | 0.0224 | 3.15 | 11.22 | 19.04 | 22.37 | ||
| LSD | 4.2E-06 | 0.0841 | 0.6501 | 0.0324 | 0.0421 | ||
| Mean | 0.0051 g | 1.67 mm | 8.78 mm | 9.16 mm2 | 5.13 g | ||
| Minimum | 0.0001 g | 0.54 mm | 3.46 mm | 0.97 mm2 | 0.07 g | ||
| Maximum | 0.0477 g | 4.23 mm | 14.55 mm | 32.13 mm2 | 47.74 g | ||
| HvMADS3 | HvMADS6 | HvMADS8 | HvMADS34 | HvMADS21 | HvMADS56 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G/A (Glu/Glu) nt 84 | G/C (Ser/Thr) nt 104 | Del nt (627-647) | Ins in promotor | C/T (Asn/Asn) nt 285 | G/A promotor | A/C (Arg/Arg) nt 9 | G/C (Ala/Ala) nt 123 | A/G nt 411 (Gly/Gly) | G/T nt 573 (Gly/Gly) | GGC Ins nt 702-704 | C Ins at nt 702 | T/A (Leu/Gln) in nt 83 | A/C (Leu/Leu) in nt 105 | G del at nt 321 | 67 bp del | |
| Bowman | G | G | C | G | A | G | A | G | T | C | ||||||
| Bonus | G | G | 4x GGC | C | G | A | G | A | G | T | C | |||||
| Foma | G | G | C | G | A | G | A | G | C | T | C | |||||
| Kristina | G | G | C | G | A | C | G | G | T | C | - | |||||
| def.2 | G | G | C | G | A | G | A | G | T | C | ||||||
| def.4 | A | G | C | G | A | C | G | G | T | A | ||||||
| def.5 | G | G | C | G | A | C | A | G | T | A | - | |||||
| def.6 | G | G | C | G | A | C | A | G | T | A | - | |||||
| def.8 | G | G | C | G | A | G | A | G | T | C | ||||||
| semi-def.1 | G | G | C | G | A | G | A | G | T | C | -67- | |||||
| semi-def.4 | G | G | C | G | A | G | A | G | A | C | ||||||
| semi-def.5 | G | G | C | G | A | G | G | G | C | A | C | |||||
| semi-def.7 | G | G | C | G | A | G | A | G | C | A | C | |||||
| semi-def.8 | G | G | C | G | A | C | A | G | T | A | - | |||||
| int-a.1 | G | G | C | G | A | G | A | G | T | C | ||||||
| int-c.5 | G | G | C | G | A | G | A | G | C | T | C | |||||
| int-e.4 | G | G | C | G | A | G | A | G | T | C | ||||||
| Int-e.20 | G | G | C | G | A | G | A | G | C | T | C | |||||
| Int-f.19 | G | G | C | G | A | G | A | G | C | T | C | |||||
| int-h.43 | G | G | C | G | A | C | G | G | T | A | - | |||||
| int-i.39 | G | G | C | G | A | G | G | G | T | A | - | |||||
| int-l.81 | G | G | C | G | A | C | A | G | T | C | ||||||
| int-m.85 | G | G | C | G | A | G | A | G | T | C | ||||||
| hex-v.3 | G | G | C | G | A | G | A | G | C | T | C | |||||
| HOR10166 | G | G | -21- | C | G | A | G | G | G | T | A | - | ||||
| HOR1555 | G | C | C | A | C | G | A | T | GGC | T | A | |||||
| HOR6211 | G | G | -18- | C | G | A | G | A | G | T | A | - | ||||
| HOR6407 | G | G | -18- | C | G | A | G | A | G | T | A | - | ||||
| HOR7191 | G | G | C | A | C | G | A | T | GGC | T | A | |||||
| HOR7041 | G | C | T | G | A | G | A | T | GGC | T | A | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, M.A.; Allam, M.; Heck, Q.K.; Urbanavičiūtė, I.; Rutten, T.; Stuart, D.; Zakhrabekova, S.; Börner, A.; Pillen, K.; Hansson, M.; et al. Analyses of MADS-box Genes Suggest HvMADS56 to Regulate Lateral Spikelet Development in Barley. Plants 2021, 10, 2825. https://doi.org/10.3390/plants10122825
Sayed MA, Allam M, Heck QK, Urbanavičiūtė I, Rutten T, Stuart D, Zakhrabekova S, Börner A, Pillen K, Hansson M, et al. Analyses of MADS-box Genes Suggest HvMADS56 to Regulate Lateral Spikelet Development in Barley. Plants. 2021; 10(12):2825. https://doi.org/10.3390/plants10122825
Chicago/Turabian StyleSayed, Mohammed A., Mohamed Allam, Quinn Kalby Heck, Ieva Urbanavičiūtė, Twan Rutten, David Stuart, Shakhira Zakhrabekova, Andreas Börner, Klaus Pillen, Mats Hansson, and et al. 2021. "Analyses of MADS-box Genes Suggest HvMADS56 to Regulate Lateral Spikelet Development in Barley" Plants 10, no. 12: 2825. https://doi.org/10.3390/plants10122825
APA StyleSayed, M. A., Allam, M., Heck, Q. K., Urbanavičiūtė, I., Rutten, T., Stuart, D., Zakhrabekova, S., Börner, A., Pillen, K., Hansson, M., & Youssef, H. M. (2021). Analyses of MADS-box Genes Suggest HvMADS56 to Regulate Lateral Spikelet Development in Barley. Plants, 10(12), 2825. https://doi.org/10.3390/plants10122825