
Influence of Culture Conditions on In Vitro Asymbiotic Germination of Anacamptis longicornu and Ophrys panormitana (Orchidaceae)

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
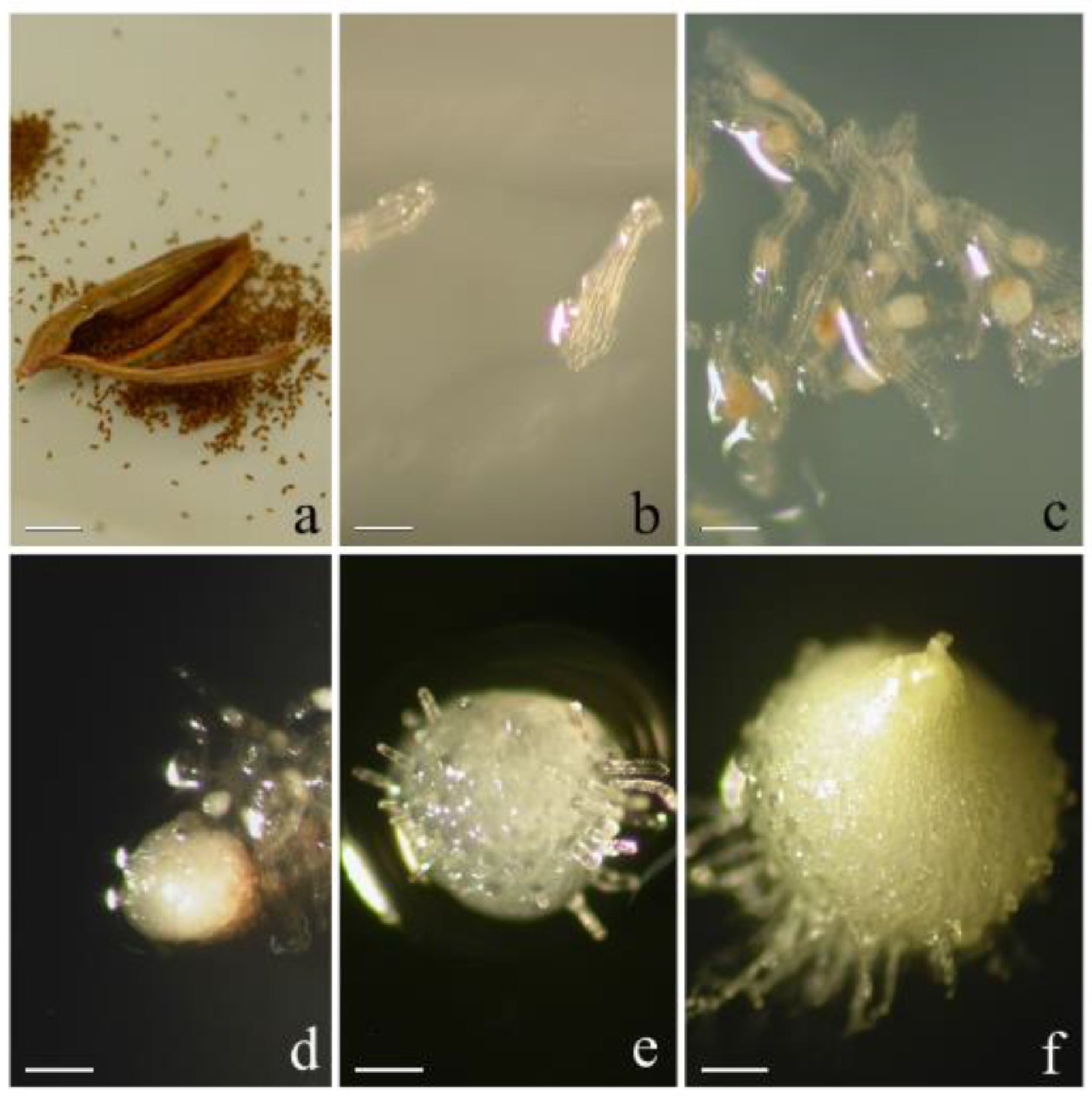
2.1. Plant Material, Seed Collection, and Axenic Culture Establishment
2.2. Culture Conditions
2.3. Germination and Protocorm Formation
2.4. Plant Acclimatization
3. Results
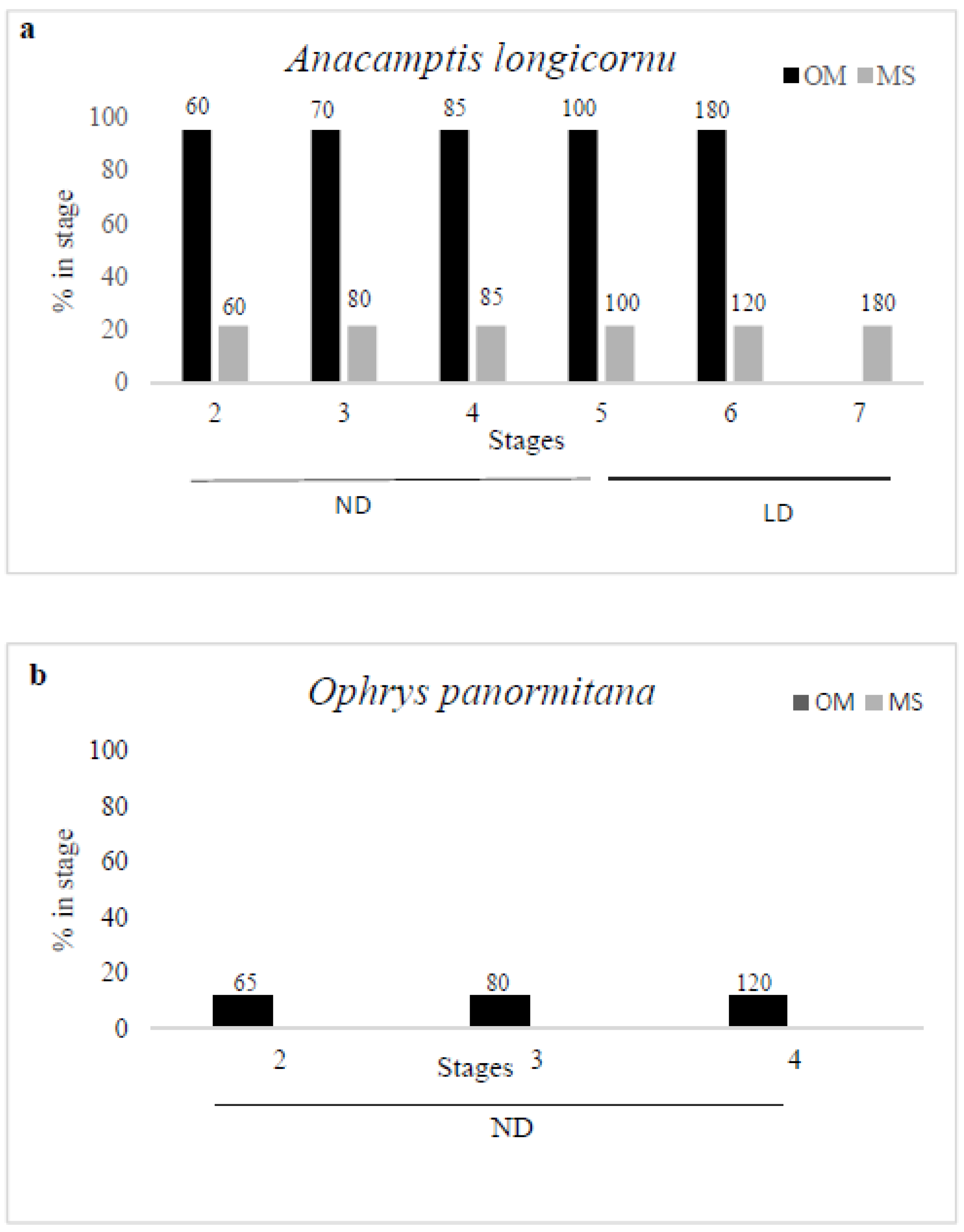
3.1. Anacamptis Longicornu
3.2. Ophrys Panormitana
3.3. Plant Acclimatization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Willis, K.J. State of the World’s Plants 2017 Report; Royal Botanic Gardens, Kew: London, UK, 2017. [Google Scholar]
- Hagsater, E.; Dumont, V.; Pridgeon, A.M. Orchids—Status Survey and Conservation Action Plan; IUCN/SSC Orchid Specialist Group; IUCN: Gland, Switzerland; Cambridge, UK, 1996; Volume 28. [Google Scholar]
- Diantina, S.; Kartikaningrum, S.; McCormick, A.C.; Millner, J.; McGill, C.; Pritchard, H.W.; Nadarajan, J. Comparative In Vitro seed germination and seedling development in tropical and temperate epiphytic and temperate terrestrial orchids. Plant Cell Tissue Organ Cult. 2020, 143, 619–633. [Google Scholar] [CrossRef]
- Kasparek, M.; Grimm, U. European trade in Turkish salep with special reference to Germany. Econ. Bot. 1999, 53, 396–406. [Google Scholar] [CrossRef]
- Lawler, L. Ethnobotany of the Orchidaceae. In Orchid Biology, Reviews & Perspectives III; Arditti, J., Ed.; Cornell University: Ithaca, NY, USA, 1984; pp. 27–149. [Google Scholar]
- Hinsley, A.; De Boer, H.J.; Fay, M.F.; Gale, S.W.; Gardiner, L.M.; Gunasekara, R.S.; Veldman, S. A review of the trade in orchids and its implications for conservation. Bot. J. Linn. Soc. 2018, 186, 435–455. [Google Scholar] [CrossRef]
- Janakiram, T.; Baskaran, V. Commercialisation and conservation aspects of orchids. J. Orchid. Soc. India 2018, 32, 55–61. [Google Scholar]
- Seaton, P.T.; Hu, H.; Perner, H.; Pritchard, H.W. Ex situ conservation of orchids in a warming world. Bot. Rev. 2010, 76, 193–203. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef]
- Volis, S.; Blecher, M. Quasi in situ: A bridge between ex situ and in situ conservation of plants. Biodivers. Conserv. 2010, 19, 2441–2454. [Google Scholar] [CrossRef]
- Kreziou, A.; de Boer, H.; Gravendeel, B. Harvesting of salep orchids in north-western Greece continues to threaten natural populations. Oryx 2016, 50, 393–396. [Google Scholar] [CrossRef]
- Ghorbani, A.; Gravendeel, B.; Zarre, S.; de Boer, H.J. Illegal wild collection and international trade of CITES-listed terrestrial orchid tubers in Iran. TRAFFIC Bull. 2014, 26, 52–58. [Google Scholar]
- Rasmussen, H.; Andersen, T.F.; Johansen, B. Temperature sensitivity of In Vitro germination and seedling development of Dactylorhiza majalis (Orchidaceae) with and without a mycorrhizal fungus. Plant Cell Environ. 1990, 13, 171–177. [Google Scholar] [CrossRef]
- Scrugli, A. Simbiosi Micorrizica. In AA.VV Orchidee d’Italia. Guida Alle Orchidee Spontanee; GIROS, Gruppo Italiano per la Ricerca delle Orchidee Spontanee; Il Castello: Cornaredo, Italy, 2009; pp. 27–29. [Google Scholar]
- Hadley, G.; Pegg, G.F. Host-fungus relationships in orchid mycorrhizal systems. In Modern Methods in Orchid Conservation: The Role of Physiology, Ecology and Management; Pritchard, H.W., Ed.; Cambridge University Press: Cambridge, UK, 1989; pp. 57–74. [Google Scholar]
- Dutra, D.; Johnson, T.R.; Kauth, P.J.; Stewart, S.L.; Kane, M.E.; Richardson, L. Asymbiotic seed germination, In Vitro seedling development, and greenhouse acclimatization of the threatened terrestrial orchid Bletia purpurea. Plant Cell Tissue Organ Cult. 2008, 94, 11–21. [Google Scholar] [CrossRef]
- Bartolo, G.; Pulvirenti, S. A check-list of Sicilian orchids. Bocconea 1997, 5, 797–824. [Google Scholar]
- Gulli, V.; Tosi, G. Anacamptis longicornu. In (Italian Group for Research on Spontaneous Orchids) Orchidee d’Italia, 2nd ed.; Guida alle Orchidee Spontanee; Il Castello: Cornaredo, Italy, 2016; p. 152. [Google Scholar]
- Biagioli, M. Ophrys sphegodes subsp. panormitana. In Orchidee d’Italia. Guida Alle Orchidee Spontanee, Cornaredo, 2nd ed.; Il Castello: Cornaredo, Italy, 2016; p. 229. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Stewart, S.L.; Zettler, L.W. Symbiotic germination of three semi-aquatic rein orchids (Habenaria repens, H. quinquiseta, H. macroceratitis) from Florida. Aquat. Bot. 2002, 72, 25–35. [Google Scholar] [CrossRef]
- Arditti, J. Factors affecting the germination of orchid seeds. Bot. Rev. 1967, 33, 1–97. [Google Scholar] [CrossRef]
- Lee, Y.I.; Lee, N.; Yeung, E.C.; Chung, M.C. Embryo development of Cypripedium formosanum in relation to seed germination In Vitro. J. Am. Soc. Hortic. Sci. 2005, 130, 747–753. [Google Scholar] [CrossRef]
- Lo, S.F.; Nalawade, S.M.; Kuo, C.L.; Chen, C.L.; Tsay, H.S. Asymbiotic germination of immature seeds, plantlet development and ex vitro establishment of plants of Dendrobium tosaense makino—A medicinally important orchid. In Vitro Cell. Dev. Biol.-Plant 2004, 40, 528–535. [Google Scholar] [CrossRef]
- Gayatri, M.C.; Kavyashree, R. Influence of carbon sources on In Vitro seed germination, protocorm and shoot formation in Vanilla planifolia. J. Curr. Sci. 2005, 7, 43–48. [Google Scholar]
- Suzuki, R.M.; Moreira, V.C.; Pescador, R.; de Melo Ferreira, W. Asymbiotic seed germination and In Vitro seedling development of the threatened orchid Hoffmannseggella cinnabarina. In Vitro Cell. Dev. Biol.-Plant 2012, 48, 500–511. [Google Scholar] [CrossRef]
- Lee, Y.I. In Vitro culture and germination of terrestrial Asian orchid seeds. In Plant Embryo Culture; Methods and Proto-Cols; Thorpe, T.A., Yeung, E.C., Eds.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2011; pp. 53–62. [Google Scholar]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Těšitelová, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Kauth, P.J.; Dutra, D.; Johnson, T.R.; Stewart, S.L.; Kane, M.E.; Vendrame, W. Techniques and applications of In Vitro orchid seed germination. In Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues; da Silva, T., Ed.; Global Science Books 5; Global Science Books, Ltd.: Isleworth, UK, 2008; pp. 375–391. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Thompson, D.I.; Edwards, T.J.; van Staden, J. Evaluating asymbiotic seed culture methods and establishing Disa (Orchidaceae) germinability In Vitro: Relationships, requirements and first-time reports. Plant Growth Regul. 2006, 49, 269–284. [Google Scholar] [CrossRef]
- Baskin, C.C.; Thompson, K.; Baskin, J.M. Mistakes in germination ecology and how to avoid them. Seed Sci. Res. 2006, 16, 165–168. [Google Scholar] [CrossRef]
- Mahendran, G.; Muniappan, V.; Ashwini, M.; Muthukumar, T.; Narmatha Bai, V. Asymbiotic seed germination of Cymbidium bicolor Lindl. (Orchidaceae) and the influence of mycorrhizal fungus on seedling development. Acta Physiol. Plant 2013, 35, 829–840. [Google Scholar] [CrossRef]
- Vogel, I.N.; Macedo, A.F. Influence of IAA, TDZ, and light quality on asymbiotic germination, protocorm formation, and plantlet development of Cyrtopodium glutiniferum Raddi., a medicinal orchid. Plant Cell Tissue Organ Cult. 2011, 104, 147–155. [Google Scholar] [CrossRef]
- Arditti, J.; Michaud, J.D.; Oliva, A.P. Practical germination of North American and related orchids. I. Epipactis atrorubens, Epipactis gigantea and Epipactis helleborine. Am. Orchid. Soc. Bull. 1982, 51, 162–171. [Google Scholar]
- Arditti, J.; Ernst, R. Physiology of germinating orchid seeds. Orchid. Biol. Rev. Perspect. 1984, 3, 177–222. [Google Scholar]
- Zeigler, E.; Grivet, C.; Assmann, S.M.; Deitzer, G.F.; Hannegan, M.W. Stomatal limitation of carbon gain in Paphiopedilum sp. (Orchidaceae) and its reversal by blue light. Plant Physiol. 1985, 77, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Van Waes, J.M.; Debergh, P.C. In vitro germination of some Western European orchids. Physiol. Plant 1986, 67, 253–261. [Google Scholar] [CrossRef]
- Zeng, S.; Wu, K.; da Silva, J.A.T.; Zhang, J.; Chen, Z.; Xia, N.; Duan, J. Asymbiotic seed germination, seedling development and reintroduction of Paphiopedilum wardii Sumerh., an endangered terrestrial orchid. Sci. Hortic. 2012, 138, 198–209. [Google Scholar] [CrossRef]
- Kunakhonnuruk, B.; Inthima, P.; Kongbangkerd, A. In vitro propagation of Epipactis flava Seidenf., an endangered rheophytic orchid: A first study on factors affecting asymbiotic seed germination, seedling development and greenhouse acclimatization. Plant Cell Tissue Organ Cult. 2018, 135, 419–432. [Google Scholar] [CrossRef]
- Parthibhan, S.; Benjamin, J.H.F.; Muthukumar, M.; Ahamed Sherif, N.; Senthil Kumar, T.; Rao, M.V. Influence of nutritional media and photoperiods on In Vitro asymbiotic seed germination and seedling development of Dendrobium aqueum Lindley. Afr. J. Plant Sci. 2012, 6, 383–393. [Google Scholar] [CrossRef]
- Anderson, A.B. The reintroduction of Platanthera ciliaris in Canada. In Proceedings of the North American Native Terrestrial Orchids: Propagation and Production. North American Native Terrestrial Orchid Conference, Washington, DC, USA, 16–17 March 1996; pp. 73–76. [Google Scholar]
- Stewart, S.L.; Kane, M.E. Asymbiotic seed germination and In Vitro seedling development of Habenaria macroceratitis (Orchidaceae), a rare Florida terrestrial orchid. Plant Cell Tissue Organ Cult. 2006, 86, 147–158. [Google Scholar] [CrossRef]
- Magrini, S.; De Vitis, M.; Torelli, D.; Santi, L.; Zucconi, L. Seed banking of terrestrial orchids: Evaluation of seed quality in Anacamptis following 4-year dry storage. Plant Biol. 2019, 21, 544–550. [Google Scholar] [CrossRef]
- Dulić, J.; Ljubojević, M.; Prlainović, I.; Barać, G.; Narandžić, T.; Ognjanov, V. Germination and Protocorm Formation of Ophrys sphegodes Mill. In Vitro Protocol for a Rare Orchid Species. Contemp. Agric. 2018, 67, 196–201. [Google Scholar] [CrossRef][Green Version]
- Dulić, J.; Ljubojević, M.; Ognjanov, V.; Barać, G.; Dulić, T. In vitro germination and seedling development of two European orchid species, Himantoglossum jankae Somlyay, Kreutz & Óvári and Spiranthes spiralis (L.) Chevall. In Vitro Cell. Dev. Biol.-Plant 2019, 55, 380–391. [Google Scholar]
- Ponert, J.; Figura, T.; Vosolsobě, S.; Lipavská, H.; Vohník, M.; Jersáková, J. Asymbiotic germination of mature seeds and protocorm development of Pseudorchis albida (Orchidaceae) are inhibited by nitrates even at extremely low concentrations. Botany 2013, 91, 662–670. [Google Scholar] [CrossRef]
- Sgarbi, E.; Grimaudo, M.; Del Prete, C. In vitro asymbiotic germination and seedling development of Limodorum abortivum (Orchidaceae). Plant Biosyst. 2009, 143, 114–119. [Google Scholar] [CrossRef]
- Raghavan, V.; Torrey, J.G. Inorganic nitrogen nutrition of the seedlings of the orchid, Cattleya. Am. J. Bot. 1964, 51, 264–274. [Google Scholar] [CrossRef]
- Zhang, Y.; Lee, Y.I.; Deng, L.; Zhao, S. Asymbiotic germination of immature seeds and the seedling development of Cypripedium macranthos Sw., an endangered lady’s slipper orchid. Sci. Hortic. 2013, 164, 130–136. [Google Scholar] [CrossRef]
- Rasmussen, H.N. Terrestrial Orchids: From Seed to Mycotrophic Plant; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Calevo, J.; Monroy, F.; Peccenini, S.; Cornara, L.; Giovannini, A. First time In Vitro asymbiotic germination of Orchis patens Desf.: A preliminary study on an endangered Mediterranean orchid. GIROS Orchid. Spontanee D’europa 2017, 60, 94–104. [Google Scholar]
- Magrini, S.; De Carli, A.; Onofri, S.; Scoppola, A. A comparative study of the seed germination capabilities of Anacamptis palustris (Orchidaceae), a threatened terrestrial orchid, and other more common Anacamptis species, by asymbiotic culture In Vitro. Eur. J. Environ. Sci. 2011, 1. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Conditions | Time from Sowing | ||||
|---|---|---|---|---|---|
| 0 Days | 45 Days | 120 Days | 6 Months | 10 Months | |
| Medium | MS | MS | MS | MS | MS |
| OM | OM | OM | OM | MS | |
| Photoperiod | LD | ND | LD | LD | LD |
| Temperature | 25 ± 1 °C | 18 ± 1 °C | 25 ± 1 °C | 25 ± 1 °C | 25 ± 1 °C |
| Stage | Description |
|---|---|
| 0 | No germination |
| 1 | Pre-germination |
| 2 | Germination |
| 3 | Protocorms |
| 4 | Appearance of protomeristem |
| 5 | Emergence of first leaf |
| 6 | Elongation of first leaf |
| 7 | Emergence of second leaf |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcidiacono, M.; Catalano, C.; Motisi, A.; Sajeva, M.; Carimi, F.; Carra, A. Influence of Culture Conditions on In Vitro Asymbiotic Germination of Anacamptis longicornu and Ophrys panormitana (Orchidaceae). Plants 2021, 10, 2543. https://doi.org/10.3390/plants10112543
Arcidiacono M, Catalano C, Motisi A, Sajeva M, Carimi F, Carra A. Influence of Culture Conditions on In Vitro Asymbiotic Germination of Anacamptis longicornu and Ophrys panormitana (Orchidaceae). Plants. 2021; 10(11):2543. https://doi.org/10.3390/plants10112543
Chicago/Turabian StyleArcidiacono, Myriam, Caterina Catalano, Antonio Motisi, Maurizio Sajeva, Francesco Carimi, and Angela Carra. 2021. "Influence of Culture Conditions on In Vitro Asymbiotic Germination of Anacamptis longicornu and Ophrys panormitana (Orchidaceae)" Plants 10, no. 11: 2543. https://doi.org/10.3390/plants10112543
APA StyleArcidiacono, M., Catalano, C., Motisi, A., Sajeva, M., Carimi, F., & Carra, A. (2021). Influence of Culture Conditions on In Vitro Asymbiotic Germination of Anacamptis longicornu and Ophrys panormitana (Orchidaceae). Plants, 10(11), 2543. https://doi.org/10.3390/plants10112543