
Transformation of European Ash (Fraxinus excelsior L.) Callus as a Starting Point for Understanding the Molecular Basis of Ash Dieback

 , , , and
, , , and 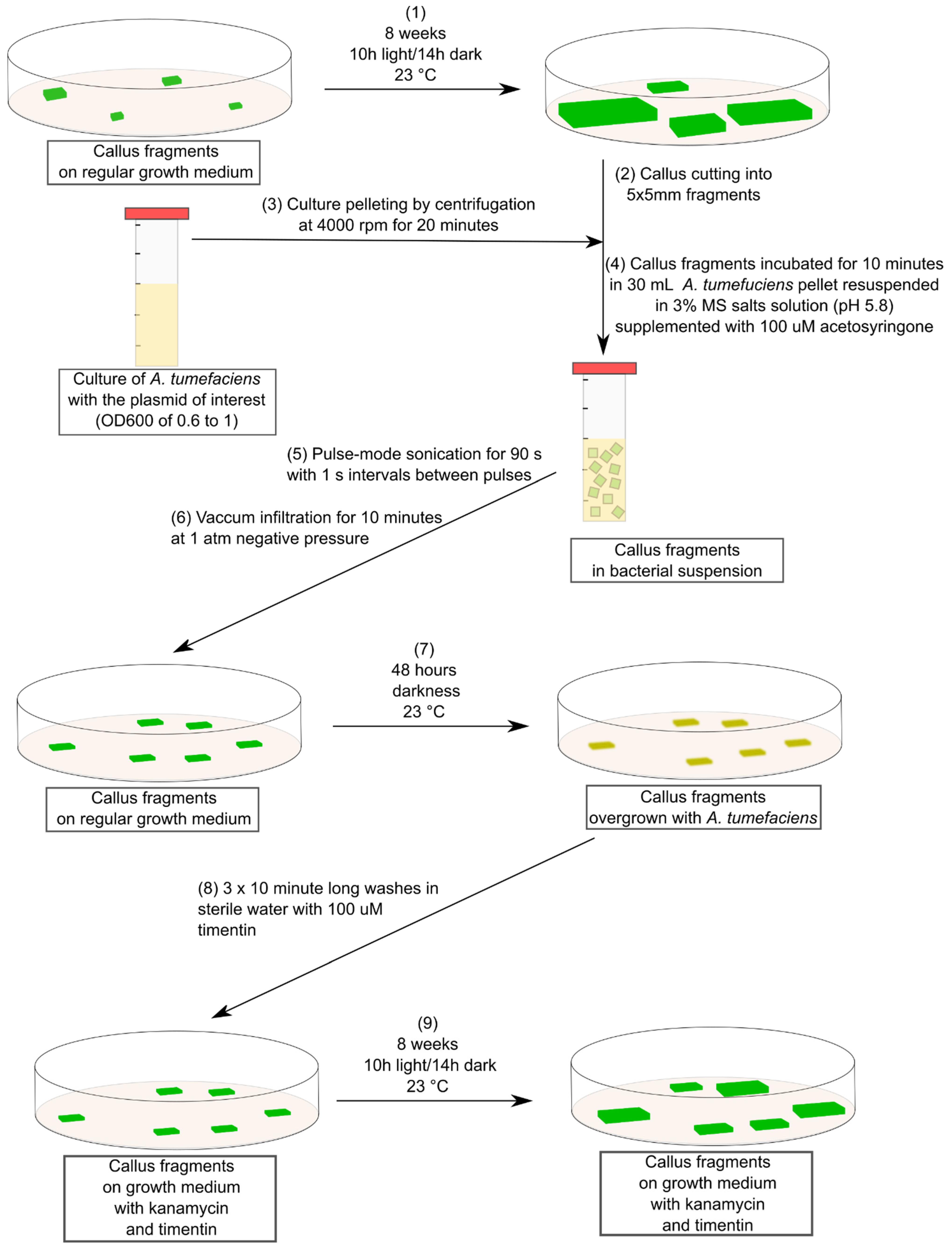{kind=link}
{kind=link}
Abstract
:1. Introduction
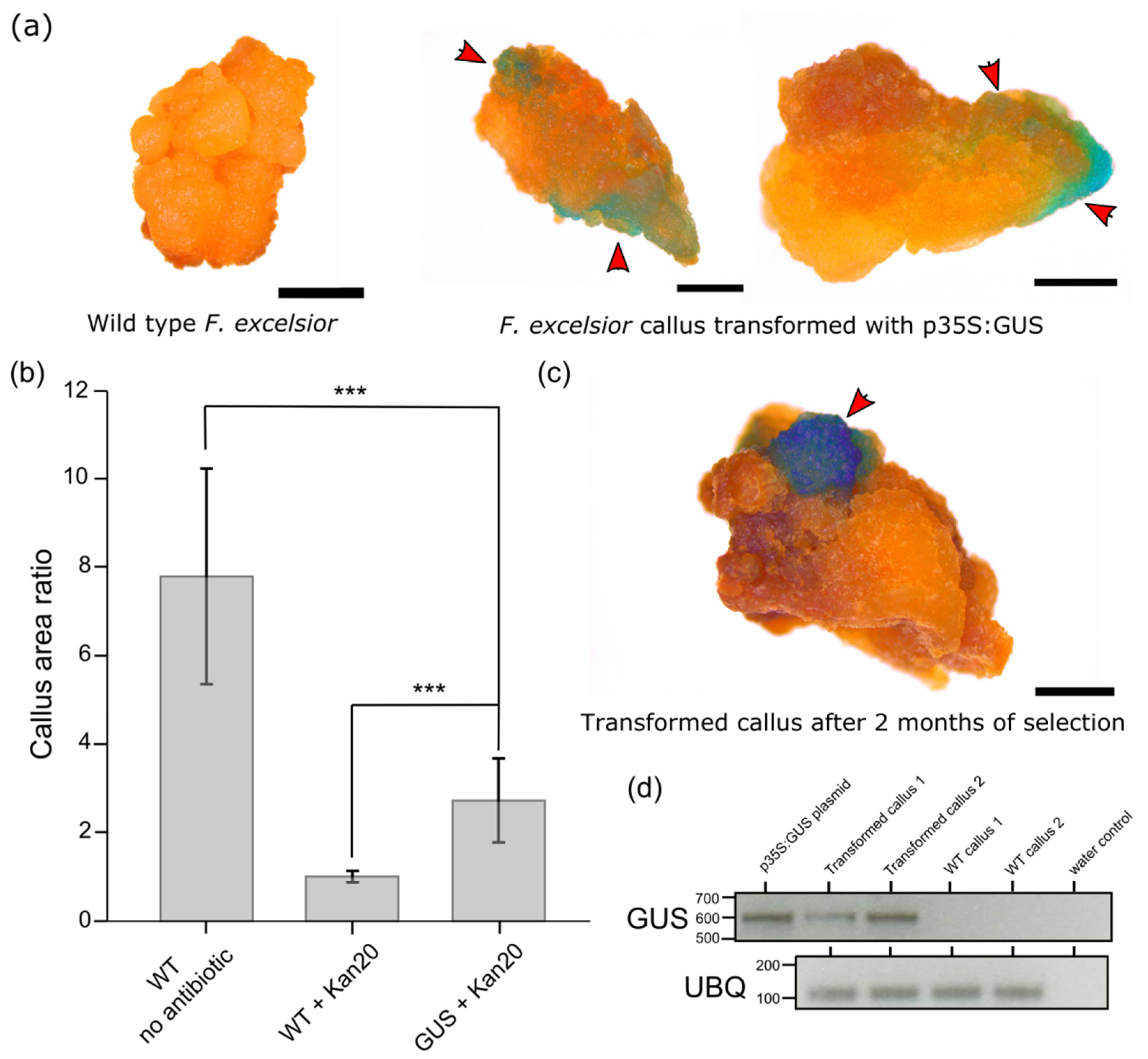
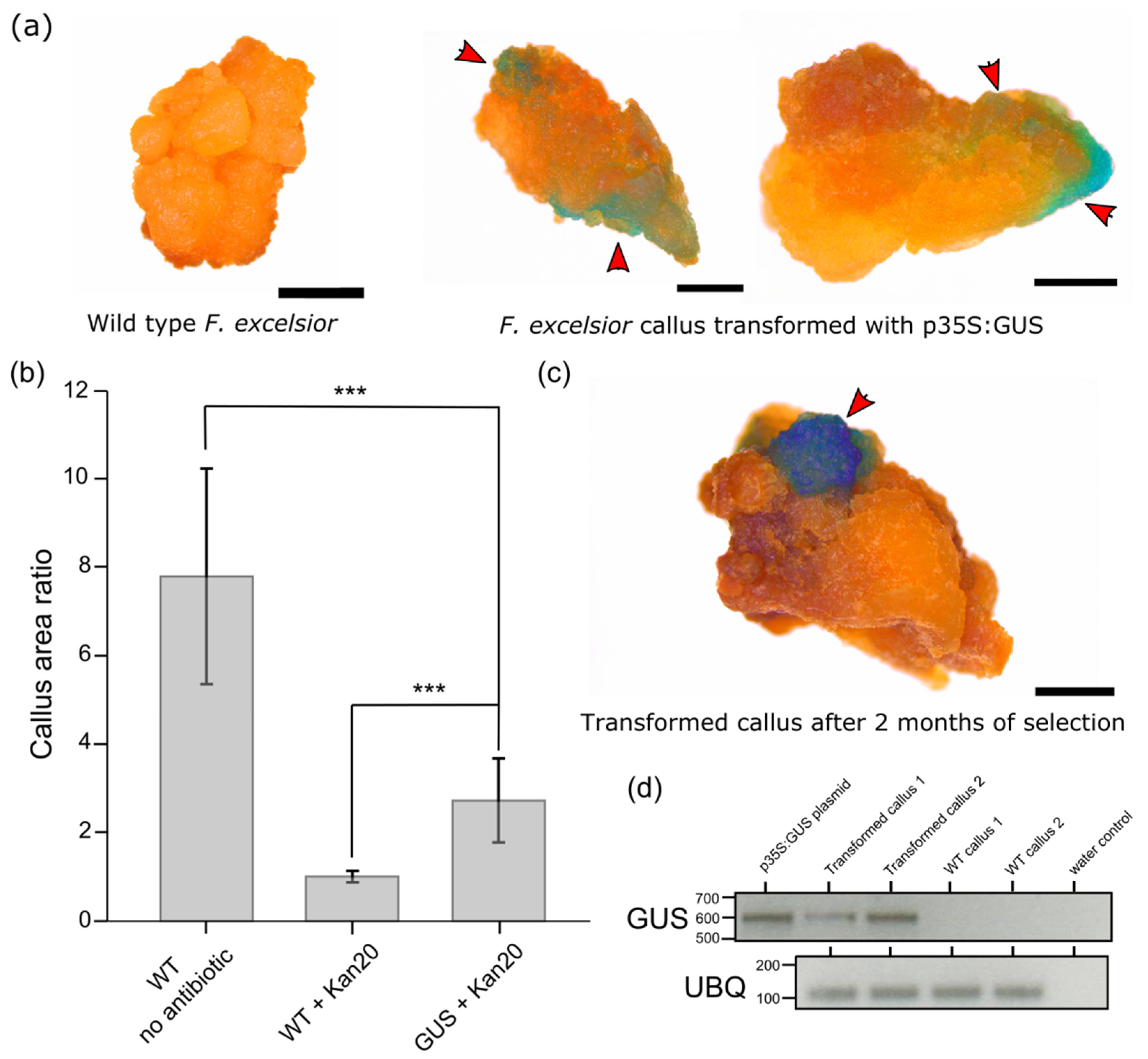
2. Results
3. Discussion
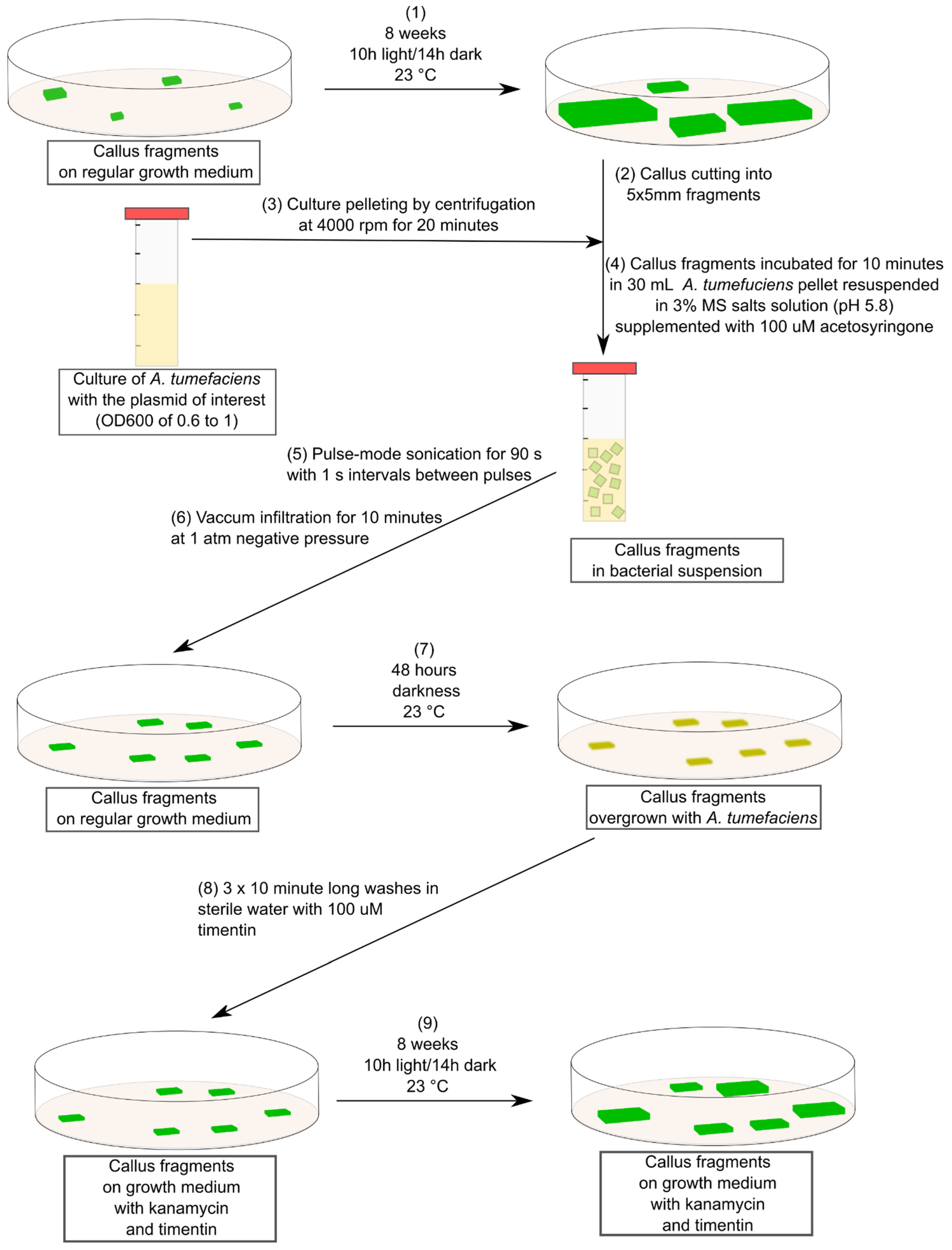
4. Materials and Methods
4.1. Plant Material Used and Culturing Conditions
4.2. Agrobacterium tumefaciens Culturing and Transformation
4.3. Callus Transformation and Selection
4.4. GUS Staining and Imaging
4.5. RT-PCR Analysis of Gene Expression
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobrowolska, D.; Hein, S.; Oosterbaan, A.; Wagner, S.; Clark, J.; Skovsgaard, J.P. A review of European ash (Fraxinus excelsior L.): Implications for silviculture. Forestry 2011, 84, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, T. Chalara fraxinea sp nov associated with dieback of ash (Fraxinus excelsior) in Poland. For. Pathol. 2006, 36, 264–270. [Google Scholar] [CrossRef]
- McMullan, M.; Rafiqi, M.; Kaithakottil, G.; Clavijo, B.J.; Bilham, L.; Orton, E.; Percival-Alwyn, L.; Ward, B.; Edwards, A.; Saunders, D.G.O.; et al. The ash dieback invasion of Europe was founded by two genetically divergent individuals. Nat. Ecol. Evol. 2018, 2, 1000–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sollars, E.S.A.; Harper, A.L.; Kelly, L.J.; Sambles, C.M.; Ramirez-Gonzalez, R.H.; Swarbreck, D.; Kaithakottil, G.; Cooper, E.D.; Uauy, C.; Havlickova, L.; et al. Genome sequence and genetic diversity of European ash trees. Nature 2017, 541, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Stocks, J.J.; Metheringham, C.L.; Plumb, W.J.; Lee, S.J.; Kelly, L.J.; Nichols, R.A.; Buggs, R.J.A. Genomic basis of European ash tree resistance to ash dieback fungus. Nat. Ecol. Evol. 2019, 3, 1686–1696. [Google Scholar] [CrossRef]
- Coker, T.L.R.; Rozsypálek, J.; Edwards, A.; Harwood, T.P.; Butfoy, L.; Buggs, R.J.A. Estimating mortality rates of European ash (Fraxinus excelsior) under the ash dieback (Hymenoscyphus fraxineus) epidemic. Plants People Planet 2019, 1, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Enderle, R.; Stenlid, J.; Vasaitis, R. An overview of ash (Fraxinus spp.) and the ash dieback disease in Europe. CAB Rev. 2019, 14, 25. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.R. Will natural resistance result in populations of ash trees remaining in British woodlands after a century of ash dieback disease? R. Soc. Open Sci. 2019, 6, 190908. [Google Scholar] [CrossRef] [Green Version]
- McKinney, L.V.; Nielsen, L.R.; Hansen, J.K.; Kjaer, E.D. Presence of natural genetic resistance in Fraxinus excelsior (Oleraceae) to Chalara fraxinea (Ascomycota): An emerging infectious disease. Heredity 2011, 106, 788–797. [Google Scholar] [CrossRef] [Green Version]
- McKinney, L.V.; Nielsen, L.R.; Collinge, D.B.; Thomsen, I.M.; Hansen, J.K.; Kjaer, E.D. The ash dieback crisis: Genetic variation in resistance can prove a long-term solution. Plant Pathol. 2014, 63, 485–499. [Google Scholar] [CrossRef]
- Wohlmuth, A.; Essl, F.; Heinze, B. Genetic analysis of inherited reduced susceptibility of Fraxinus excelsior L. seedlings in Austria to ash dieback. Forestry 2018, 91, 514–525. [Google Scholar] [CrossRef]
- Villari, C.; Dowkiw, A.; Enderle, R.; Ghasemkhani, M.; Kirisits, T.; Kjaer, E.D.; Marciulyniene, D.; McKinney, L.V.; Metzler, B.; Munoz, F.; et al. Advanced spectroscopy-based phenotyping offers a potential solution to the ash dieback epidemic. Sci. Rep. 2018, 8, 17448. [Google Scholar] [CrossRef] [Green Version]
- Aoun, M.; Rioux, D.; Simard, M.; Bernier, L. Fungal Colonization and Host Defense Reactions in Ulmus americana Callus Cultures Inoculated with Ophiostoma Novo-Ulmi. Phytopathol. 2009, 99, 642–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrot-Chorabik, K.; Grad, B.; Kowalski, T. Interactions between callus cultures of Pinus sylvestris and pine fungi with different trophic properties. For. Pathol. 2016, 46, 179–186. [Google Scholar] [CrossRef]
- Efferth, T. Biotechnology Applications of Plant Callus Cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Du, N.X.; Pijut, P.M. Agrobacterium-mediated transformation of Fraxinus pennsylvanica hypocotyls and plant regeneration. Plant Cell Rep. 2009, 28, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Palla, K.J.; Pijut, P.M. Agrobacterium-mediated genetic transformation of Fraxinus americana hypocotyls. Plant Cell Tissue Organ Cult. 2015, 120, 631–641. [Google Scholar] [CrossRef]
- Stevens, M.E.; Pijut, P.M. Agrobacterium-mediated genetic transformation and plant regeneration of the hardwood tree species Fraxinus profunda. Plant Cell Rep. 2014, 33, 861–870. [Google Scholar] [CrossRef]
- Bellincampi, D.; Cervone, F.; Lionetti, V. Plant cell wall dynamics and wall-related susceptibility in plant-pathogen interactions. Front. Plant Sci. 2014, 5, 228. [Google Scholar] [CrossRef] [Green Version]
- Cleary, M.R.; Daniel, G.; Stenlid, J. Light and scanning electron microscopy studies of the early infection stages of Hymenoscyphus pseudoalbidus on Fraxinus excelsior. Plant Pathol. 2013, 62, 1294–1301. [Google Scholar] [CrossRef]
- Bacete, L.; Melida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Latowski, D.; Nawrot-Chorabik, K. The Method of Obtaining Saplings of the Common Ash (Fraxinus excelsior L.) and the Media Suitable for Use in This Method. European Patent WO2021187995A2, 23 September 2021. Available online: https://worldwide.espacenet.com/patent/search/family/075954228/publication/WO2021187995A2?q=PCT%2FPL2021%2F050017 (accessed on 3 November 2021).
- Song, G.Q.; Prieto, H.; Orbovic, V. Agrobacterium-Mediated Transformation of Tree Fruit Crops: Methods, Progress, and Challenges. Front. Plant Sci. 2019, 10, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutu, C.; Brandle, J.; Brown, D.; Brown, K.; Miki, B.; Simmonds, J.; Hegedus, D.D. pORE: A modular binary vector series suited for both monocot and dicot plant transformation. Transgenic Res. 2007, 16, 771–781. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hebda, A.; Liszka, A.; Zgłobicki, P.; Nawrot-Chorabik, K.; Lyczakowski, J.J. Transformation of European Ash (Fraxinus excelsior L.) Callus as a Starting Point for Understanding the Molecular Basis of Ash Dieback. Plants 2021, 10, 2524. https://doi.org/10.3390/plants10112524
Hebda A, Liszka A, Zgłobicki P, Nawrot-Chorabik K, Lyczakowski JJ. Transformation of European Ash (Fraxinus excelsior L.) Callus as a Starting Point for Understanding the Molecular Basis of Ash Dieback. Plants. 2021; 10(11):2524. https://doi.org/10.3390/plants10112524
Chicago/Turabian StyleHebda, Anna, Aleksandra Liszka, Piotr Zgłobicki, Katarzyna Nawrot-Chorabik, and Jan J. Lyczakowski. 2021. "Transformation of European Ash (Fraxinus excelsior L.) Callus as a Starting Point for Understanding the Molecular Basis of Ash Dieback" Plants 10, no. 11: 2524. https://doi.org/10.3390/plants10112524
APA StyleHebda, A., Liszka, A., Zgłobicki, P., Nawrot-Chorabik, K., & Lyczakowski, J. J. (2021). Transformation of European Ash (Fraxinus excelsior L.) Callus as a Starting Point for Understanding the Molecular Basis of Ash Dieback. Plants, 10(11), 2524. https://doi.org/10.3390/plants10112524