
Assessing the Weed-Suppressing Potential of Cotton Chromosome Substitution Lines Using the Stair-Step Assay

 ,
,
Abstract
1. Introduction
2. Results
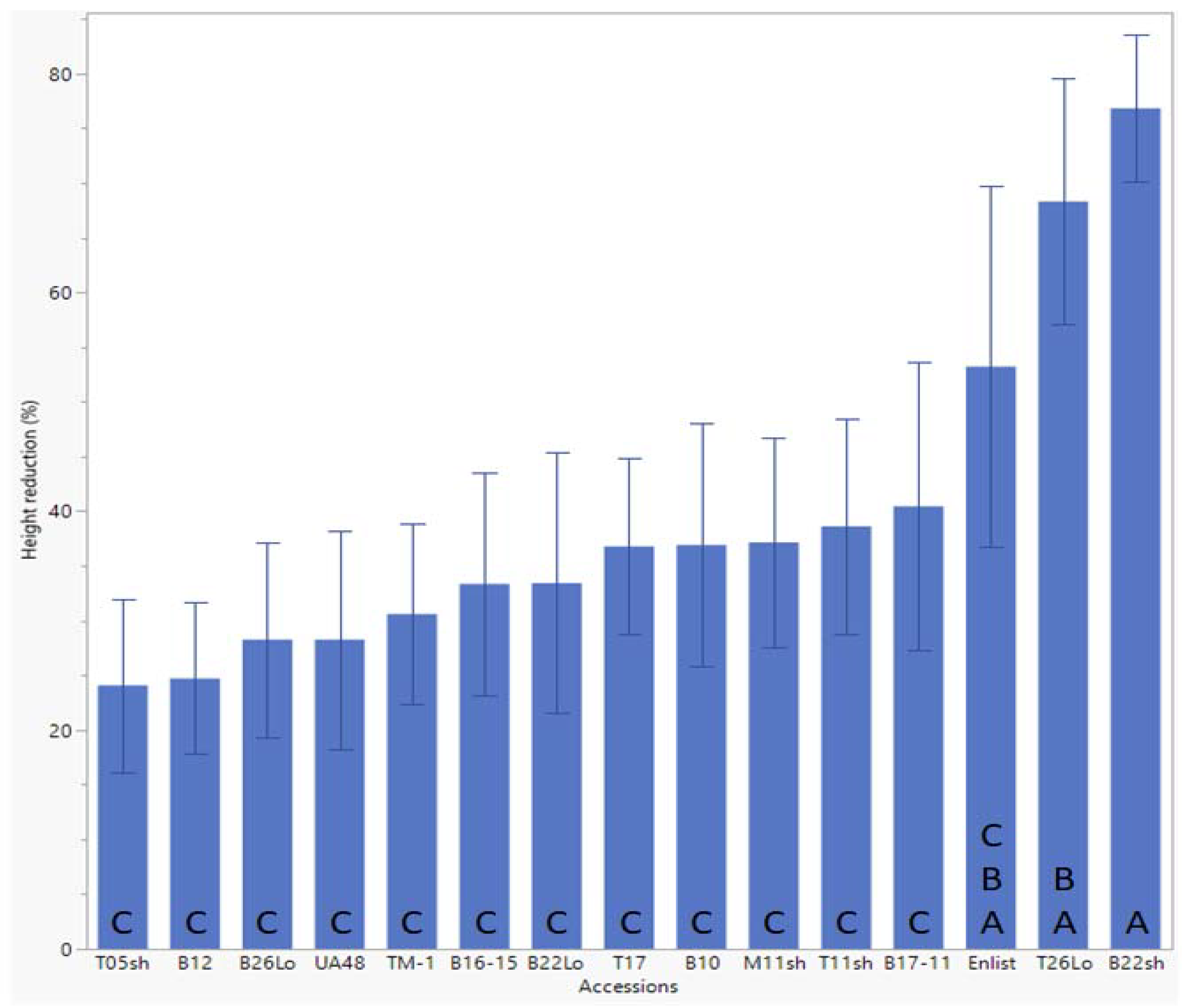
2.1. Height Reduction
2.2. Chlorophyll Reduction
2.3. Cotton Reduction
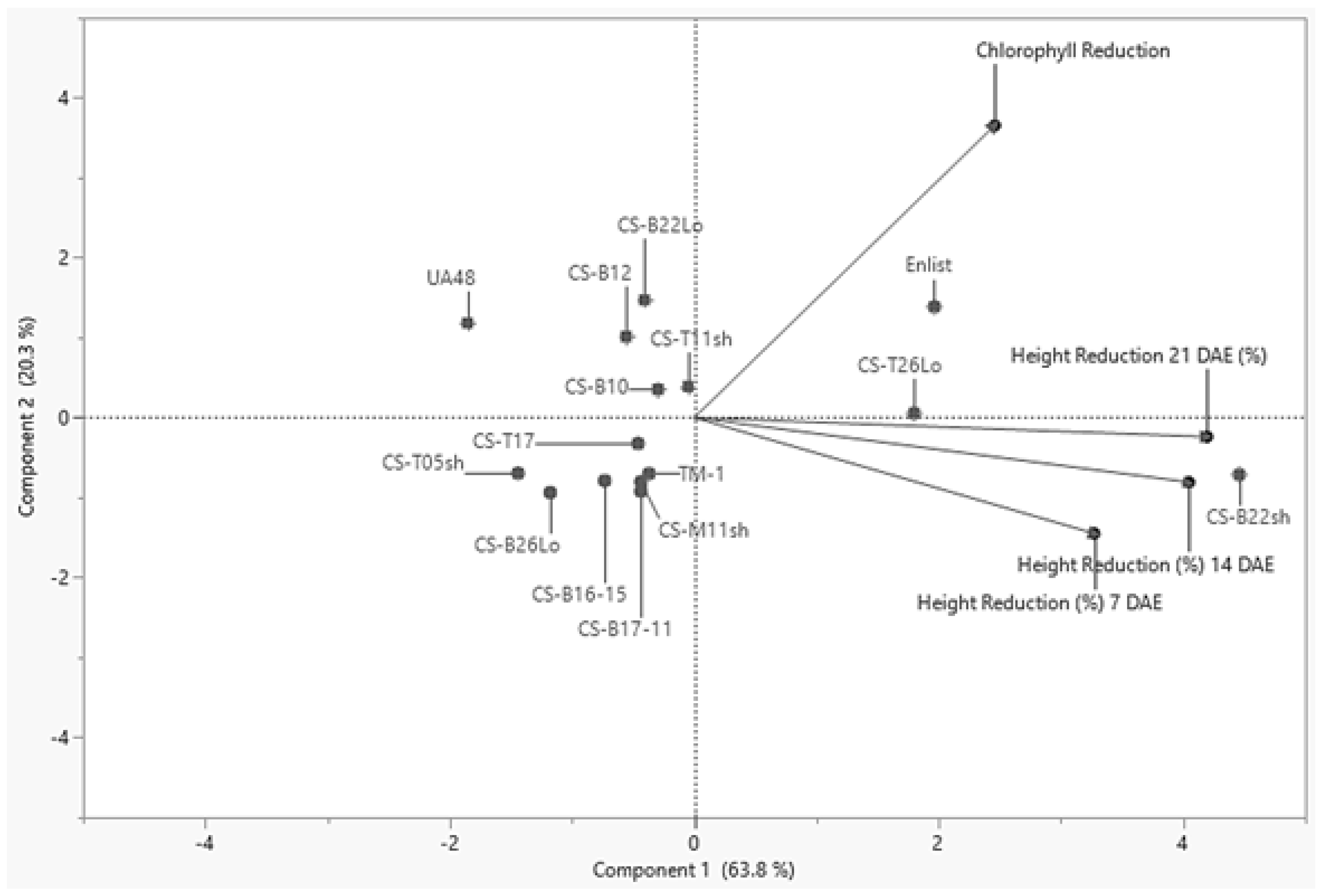
2.4. Hierarchical Clustering and Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
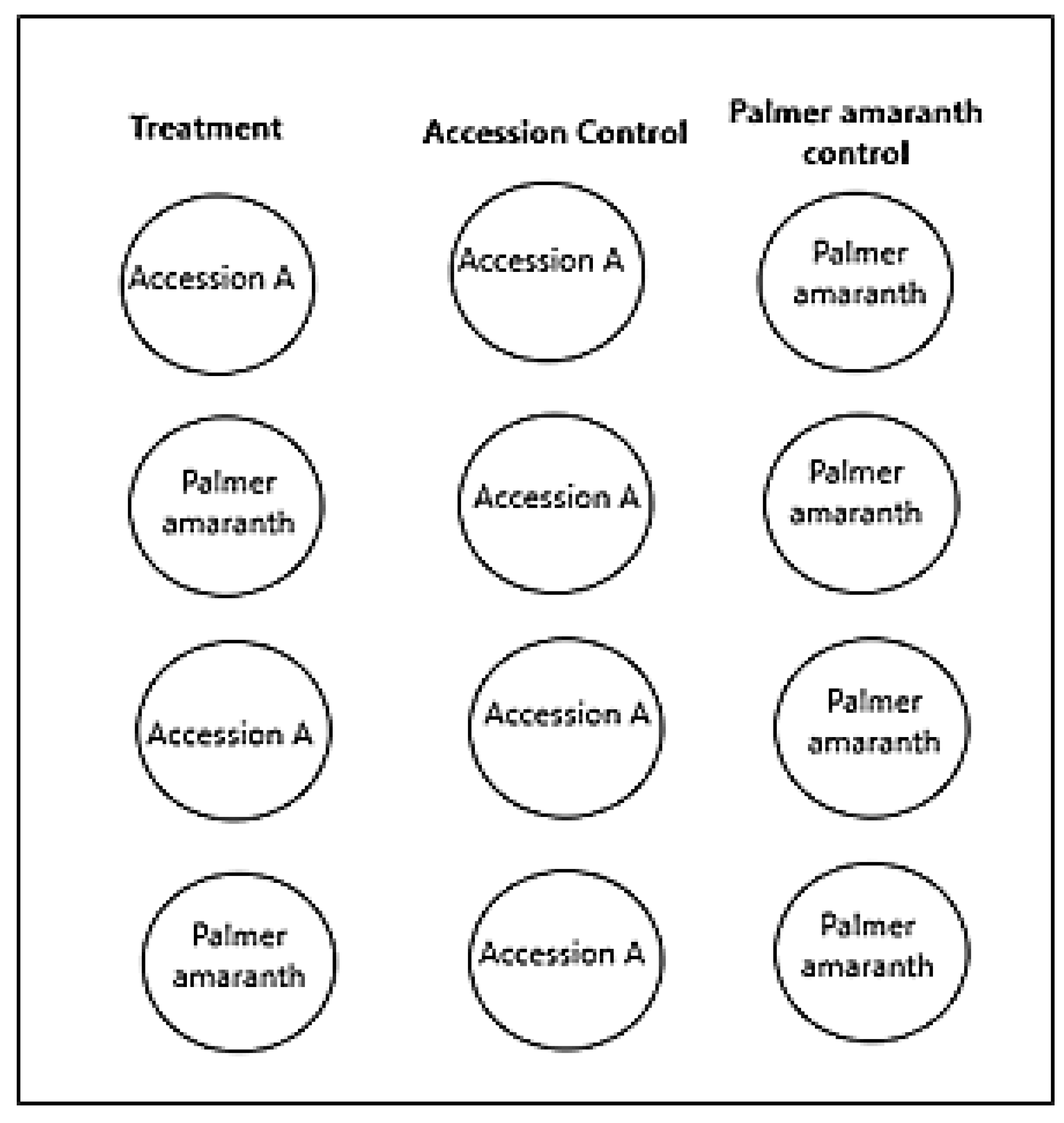
4.2. Germination and Preparation of Plants Used in the Stairstep Structure
4.3. Design of Stairstep Structure
4.4. Methods
4.5. Calculations
PA (cm)] × 100
/cc of control PA (unit)] × 100
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zimdahl, R.L. Fundamentals of Weed Science; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Buchholtz, K.P. Report of the terminology committee of the Weed Science Society of America. Weeds 1967, 15, 388–389. [Google Scholar]
- Humburg, N.E.; Colby, S.R. Herbicide Handbook of the Weed Science Society of America, 6th ed.; Weed Science Society of America: Chicago, IL, USA, 1989. [Google Scholar]
- Culpepper, A.S.; Webster, T.M.; Sosnoskie, L.M.; York, A.C.; Nandula, V.K. Glyphosate-resistant Palmer amaranth in the United States. In Glyphosate Resistance in Crops and Weeds: History, Development, and Management; John Wiley & Sons: Hoboken, NJ, USA, 2010; pp. 195–212. [Google Scholar]
- Morgan, G.D.; Baumann, P.A.; Chandler, J.M. Competitive Impact of Palmer Amaranth (Amaranthus palmeri) on Cotton (Gossypium hirsutum) Development and Yield. Weed Technol. 2001, 15, 408–412. [Google Scholar] [CrossRef]
- Webster, T.M.; Nichols, R.L. Changes in the prevalence of weed species in the major agronomic crops of the Southern United States: 1994/1995 to 2008/2009. Weed Sci. 2012, 60, 145–157. [Google Scholar] [CrossRef]
- Braxton, L.B.; Richburg, J.S.; York, A.C.; Culpepper, A.S.; Haygood, R.A.; Lovelace, M.L.; Walton, L.C. Resistance of Enlist™(AAD-12) Cotton to Glufosinate. Weed Technol. 2017, 31, 380–386. [Google Scholar] [CrossRef]
- Smith, D.T.; Baker, R.V.; Stelle, G.L. Palmer Amaranth (Amaranthus palmeri) Impacts o Yield, Harvesting, and Ginning in Dryland Cotton (Gossypium hirsutum). Weed Technol. 2000, 14, 122–126. [Google Scholar] [CrossRef]
- Appleby, A.P. A history of weed control in the United States and Canada—A sequel. Weed Sci. 2005, 53, 762–768. [Google Scholar] [CrossRef]
- Webster, T.M.; Sosnoskie, L.M. Loss of glyphosate efficacy: A changing weed spectrum in Georgia cotton. Weed Sci. 2010, 58, 73–79. [Google Scholar] [CrossRef]
- Srinivasan, P.R.; Shigeura, H.D.T.; Sprecher, M.; Sprinson, D.B.; Davis, B.D. The biosynthesis of shikimic acid from D-glucose. J. Biol. Chem. 1956, 220, 477–497. [Google Scholar] [CrossRef]
- Xu, J.; Smith, S.; Smith, G.; Wang, W.; Li, Y. Glyphosate contamination in grains and foods: An overview. Food Control. 2019, 106, 106710. [Google Scholar] [CrossRef]
- Székács, A.; Darvas, B. Re-registration challenges of glyphosate in the European Union. Front. Environ. Sci. 2018, 6, 78. [Google Scholar] [CrossRef]
- Mesnage, R.; Bernay, B.; Séralini, G.E. Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology 2013, 313, 122–128. [Google Scholar] [CrossRef] [PubMed]
- VanGessel, M.J. Glyphosate-resistant horseweed from Delaware. Weed Sci. 2001, 49, 703–705. [Google Scholar] [CrossRef]
- Comont, D.; Hicks, H.; Crook, L.; Hull, R.; Cocciantelli, E.; Hadfield, J.; Neve, P. Evolutionary epidemiology predicts the emergence of glyphosate resistance in a major agricultural weed. New Phytol. 2019, 223, 1584–1594. [Google Scholar] [CrossRef] [PubMed]
- Tranel, P.J.; Wright, T.R.; Heap, I.M. The International Survey of Herbicide Resistant Weeds. 2019. Available online: www.weedscience.org/Summary/MOA.aspx?MOAID=3 (accessed on 7 November 2021).
- Norsworthy, J.K.; Griffith, G.M.; Scott, R.C.; Smith, K.L.; Oliver, L.R. Confirmation and control of glyphosate-resistant Palmer amaranth (Amaranthus palmeri) in Arkansas. Weed Technol. 2008, 22, 108–113. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Ward, S.M.; Shaw, D.R.; Llewellyn, R.S.; Nichols, R.L.; Webster, T.M.; Barrett, M. Reducing the risks of herbicide resistance: Best management practices and recommendations. Weed Sci. 2012, 60, 31–62. [Google Scholar] [CrossRef]
- Charles, G.; Taylor, I.; Roberts, G. The impact of the cotton farming system on weed succession: Implications for herbicide resistance and adoption of an integrated weed management approach. In Proceedings of the 14th Australian Weeds Conference, Wagga Wagga, NSW, Australia, 6–9 September 2004; Sindel, B.M., Johnson, S.B., Eds.; 2004; pp. 410–413. [Google Scholar]
- Kong, C.H.; Chen, X.H.; Hu, F.; Zhang, S.Z. Breeding of commercially acceptable allelopathic rice cultivars in China. Pest. Manag. Sci. 2011, 67, 1100–1106. [Google Scholar] [CrossRef]
- Sun, B.; Kong, C.H.; Wang, P.; Qu, R. Response and relation of allantoin production in different rice cultivars to competing barnyardgrass. Plant. Ecol. 2012, 213, 1917–1926. [Google Scholar] [CrossRef]
- Rizvi, S.J. Allelopathy: Basic and Applied Aspects; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Hoffman, M.L.; Weston, L.A.; Snyder, J.C.; Regnier, E.E. Separating the effects of sorghum (Sorghum bicolor) and rye (Secale cereale) root and shoot residues on weed development. Weed Sci. 1996, 44, 402–407. [Google Scholar] [CrossRef]
- Gui-Ying, J.; Jian-Guo, L.; Yan-Bin, L. Allelochemicals from cotton (Gossypium hirsutum) rhizosphere soil: Inhibitory effects on cotton seedlings. Allelopath. J. 2015, 35, 153–162. [Google Scholar]
- Hall, A.B.; Blum, U.; Fites, R.C. Stress modification of allelopathy of Helianthus annuus L. debris on seed germination. Am. J. Bot. 1982, 69, 776–783. [Google Scholar] [CrossRef]
- Saha, S.; Jenkins, J.N.; Wu, J.; McCarty, J.C.; Gutiérrez, O.A.; Percy, R.G.; Stelly, D.M. Effects of chromosome-specific introgression in upland cotton on fiber and agronomic traits. Genetics 2006, 172, 1927–1938. [Google Scholar] [CrossRef][Green Version]
- Chen, Z.J.; Scheffler, B.E.; Dennis, E.; Triplett, B.A.; Zhang, T.; Guo, W.; Paterson, A.H. Toward sequencing cotton (Gossypium) genomes. Plant. Physiol. 2007, 145, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Wendel, J.F.; Brubaker, C.L.; Percival, A.E. Genetic diversity in Gossypium hirsutum and the origin of upland cotton. Am. J. Bot. 1992, 79, 1291–1310. [Google Scholar] [CrossRef]
- Saha, S.; Raska, D.A.; Stelly, D.M. Upland Cotton (Gossypium hirsutum L.) x Hawaiian Cotton (G. tomentosum Nutt. Ex. Seem.) F1 hybrid hypoaneuploid chromosome substitution series. J. Cotton Sci. 2006, 10, 146–154. [Google Scholar]
- Saha, S.; Stelly, D.M.; Raska, D.A.; Wu, J.; Jenkins, J.N.; McCarty, J.C.; Campbell, B.T. Chromosome substitution lines: Concept, development and utilization in the genetic improvement of Upland cotton. Plant. Breed. InTech Slavka Krautzeka 2011, 83, 107–128. [Google Scholar]
- Wu, Z.; Soliman, K.M.; Bolton, J.J.; Saha, S.; Jenkins, J.N. Identification of differentially expressed genes associated with cotton fiber development in a chromosomal substitution line (CS-CS-B22sh). Funct. Integr. Genom. 2008, 8, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Brubaker, C.L.; Bourland, F.M.; Wendel, J.F. The origin and domestication of cotton. In Cotton: Origin, History, Technology, and Production; John Wiley & Sons: New York, NY, USA, 1999; pp. 3–31. [Google Scholar]
- Awasthi, A.; Reddy, K.R.; Saha, S.; Jenkins, J.N.; Stelly, D.M. Morph-physiological responses of cotton interspecific chromosome substitution lines to low temperature and drought stresses. Euphytica 2018, 214, 1–19. [Google Scholar] [CrossRef]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology. Evol. Syst. 2004, 6, 207–215. [Google Scholar]
- Kruse, M.; Strandberg, M.; Strandberg, B. Ecological Effects of Allelopathic Plants-A Review; NERI Technical Report; National Environmental Research Institute: Silkeborg, Denmark, 2000; p. 315. [Google Scholar]
- Schumaker, B.C.; Stallworth, S.; de Castro, E.; Fuller, M.G.; Shrestha, S.; Tseng, T.M. Repeatable Stair-Step Assay to Access the Allelopathic Potential of Weedy Rice (Oryza sativa ssp.). JoVE (J. Vis. Exp.) 2016, 55, e60764. [Google Scholar] [CrossRef]
- Camberato, J.J. Cation exchange capacity-everything you want to know and much more. Magnesium 2007, 2, 240. [Google Scholar]
- Bednarz, C.W.; Oosterhuis, D.M.; Evans, R.D. Leaf photosynthesis and carbon isotope discrimination of cotton in response to potassium deficiency. Environ. Exp. Bot. 1998, 39, 131–139. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T. Allelopathy in wheat (Triticum aestivum). Ann. Appl. Biol. 2001, 139, 1–9. [Google Scholar] [CrossRef]
- Putnam, A.R.; DeFrank, J.; Barnes, J.P. Exploitation of allelopathy for weed control in annual and perennial cropping systems. J. Chem. Ecol. 1983, 9, 1001–1010. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cotton Accession | PA Mean Height Reduction 7 DAE (%) | PA Mean Height Reduction 14 DAE (%) | PA Mean Height Reduction 21 DAE (%) | PA Mean Chlorophyll Reduction (%) | Mean Susceptibility (%) |
|---|---|---|---|---|---|
| CS-T05sh | 22.94 | 32.92 | 24.06 c | 9.78 | 22.43 |
| CS-B12 | 12.17 | 41.50 | 24.70 c | 19.66 | 24.51 |
| CS-B26Lo | 24.65 | 35.27 | 28.24 c | 8.98 | 24.29 |
| UA48 | 10.68 | 27.71 | 28.24 c | 16.50 | 20.78 |
| TM-1 | 21.66 | 44.96 | 30.58 c | 11.85 | 27.26 |
| CS-B16-15 | 20.82 | 46.23 | 33.34 c | 9.81 | 27.55 |
| CS-B22Lo | 20.13 | 41.72 | 33.42 c | 21.18 | 29.11 |
| CS-T17 | 30.23 | 39.70 | 36.77 c | 12.85 | 29.89 |
| CS-B10 | 29.65 | 38.11 | 36.92 c | 16.49 | 30.30 |
| CS-M11sh | 30.10 | 44.42 | 37.15 c | 10.46 | 30.53 |
| CS-T11sh | 23.17 | 42.17 | 38.60 c | 17.00 | 30.24 |
| CS-B17-11 | 27.73 | 35.71 | 40.42 bc | 10.17 | 28.51 |
| Enlist® | 19.18 | 44.60 | 53.20 abc | 26.49 | 35.87 |
| CS-T26Lo | 21.93 | 71.52 | 68.32 ab | 16.42 | 44.55 |
| CS-B22sh | 55.23 | 88.11 | 76.82 a | 20.07 | 60.06 |
| Cotton Accession | Cotton Height Reduction 7 DAE (%) | Cotton Height Reduction 14 DAE (%) | Cotton Height Reduction 21 DAE (%) | Cotton Chlorophyll Reduction (%) | Mean Susceptibility (%) |
|---|---|---|---|---|---|
| CS-B22sh | 4.99 | 14.41 abc | 2.56 c | 0.47 | 5.61 |
| CS-B16-15 | 18.19 | 6.54 c | 9.28 bcd | 3.13 | 9.29 |
| CS-T11sh | 10.97 | 5.43 c | 3.58 cd | 2.87 | 5.71 |
| CS-T26Lo | 10.50 | 26.93 ab | 9.13 bcd | 15.02 | 15.40 |
| CS-B17-11 | 26.29 | 3.18 c | 2.53 d | 3.50 | 8.88 |
| CS-M11sh | 16.91 | 10.16 c | 6.59 bcd | 5.92 | 9.90 |
| CS-B10 | 21.57 | 14.10 bc | 6.55 bcd | 2.86 | 11.27 |
| UA48 | 5.16 | 12.25 bc | 7.65 bcd | 0.41 | 6.37 |
| TM-1 | 3.25 | 12.25 bc | 3.41 cd | 4.93 | 5.96 |
| Enlist® | 16.67 | 4.66 c | 21.61 ab | 5.27 | 12.05 |
| CS-T17 | 12.61 | 4.09 c | 3.94 cd | 4.76 | 6.35 |
| CS-T05sh | 4.65 | 5.84 c | 5.49 cd | 3.33 | 4.83 |
| CS-B26Lo | 11.70 | 9.79 c | 16.51 abc | 2.40 | 10.10 |
| CS-B22Lo | 16.40 | 31.96 a | 31.33 a | 4.07 | 20.94 |
| CS-B12 | 16.50 | 5.58 c | 9.08 bcd | 4.88 | 9.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuller, M.G.; Saha, S.; Stelly, D.M.; Jenkins, J.N.; Tseng, T.M. Assessing the Weed-Suppressing Potential of Cotton Chromosome Substitution Lines Using the Stair-Step Assay. Plants 2021, 10, 2450. https://doi.org/10.3390/plants10112450
Fuller MG, Saha S, Stelly DM, Jenkins JN, Tseng TM. Assessing the Weed-Suppressing Potential of Cotton Chromosome Substitution Lines Using the Stair-Step Assay. Plants. 2021; 10(11):2450. https://doi.org/10.3390/plants10112450
Chicago/Turabian StyleFuller, Mary Gracen, Sukumar Saha, David M. Stelly, Johnie N. Jenkins, and Te Ming Tseng. 2021. "Assessing the Weed-Suppressing Potential of Cotton Chromosome Substitution Lines Using the Stair-Step Assay" Plants 10, no. 11: 2450. https://doi.org/10.3390/plants10112450
APA StyleFuller, M. G., Saha, S., Stelly, D. M., Jenkins, J. N., & Tseng, T. M. (2021). Assessing the Weed-Suppressing Potential of Cotton Chromosome Substitution Lines Using the Stair-Step Assay. Plants, 10(11), 2450. https://doi.org/10.3390/plants10112450