
Floral Nectar Chemistry in Orchids: A Short Review and Meta-Analysis


Abstract
1. Introduction
2. Results
2.1. Description of the Database
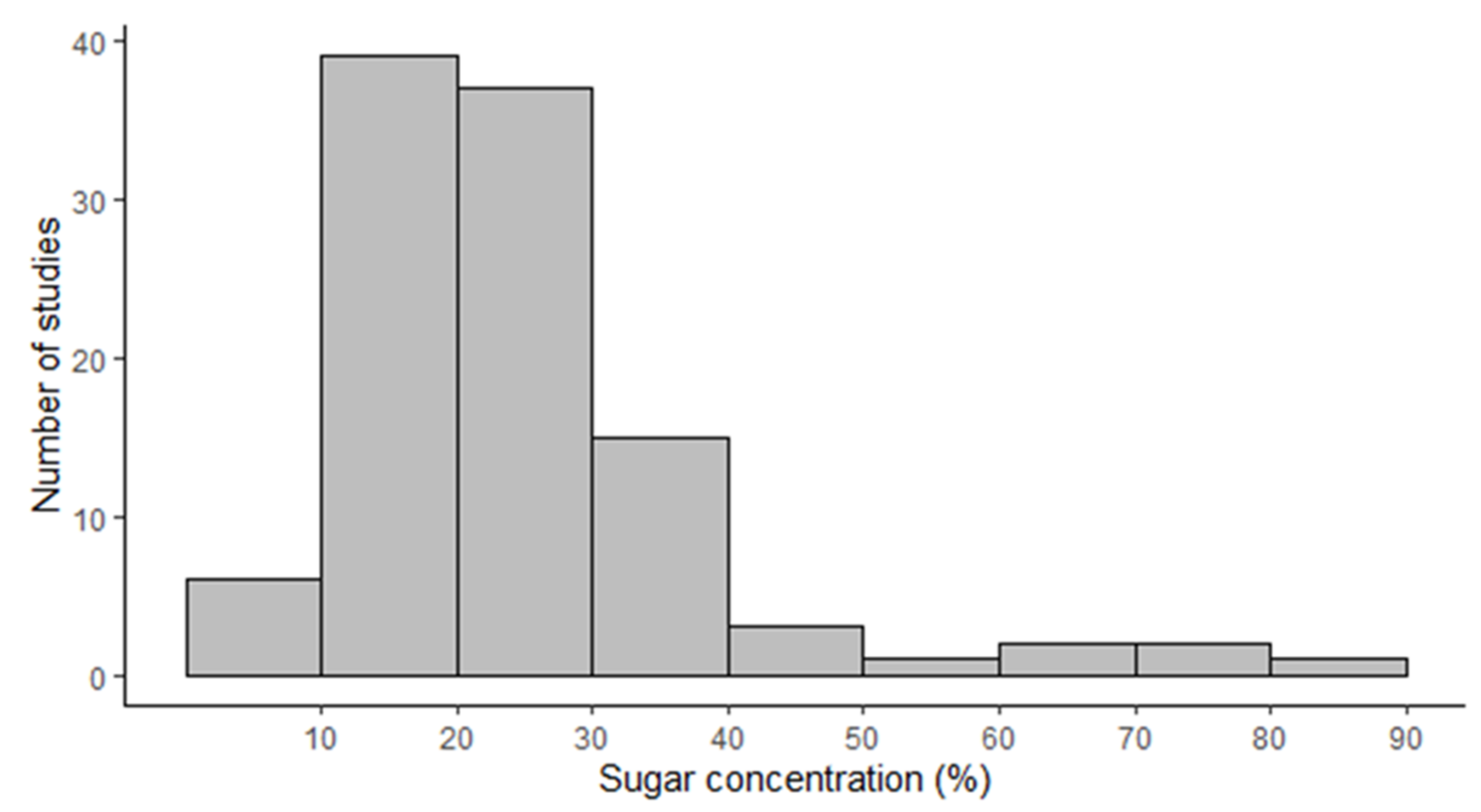
2.2. Overall Characteristics of Nectar in Orchids
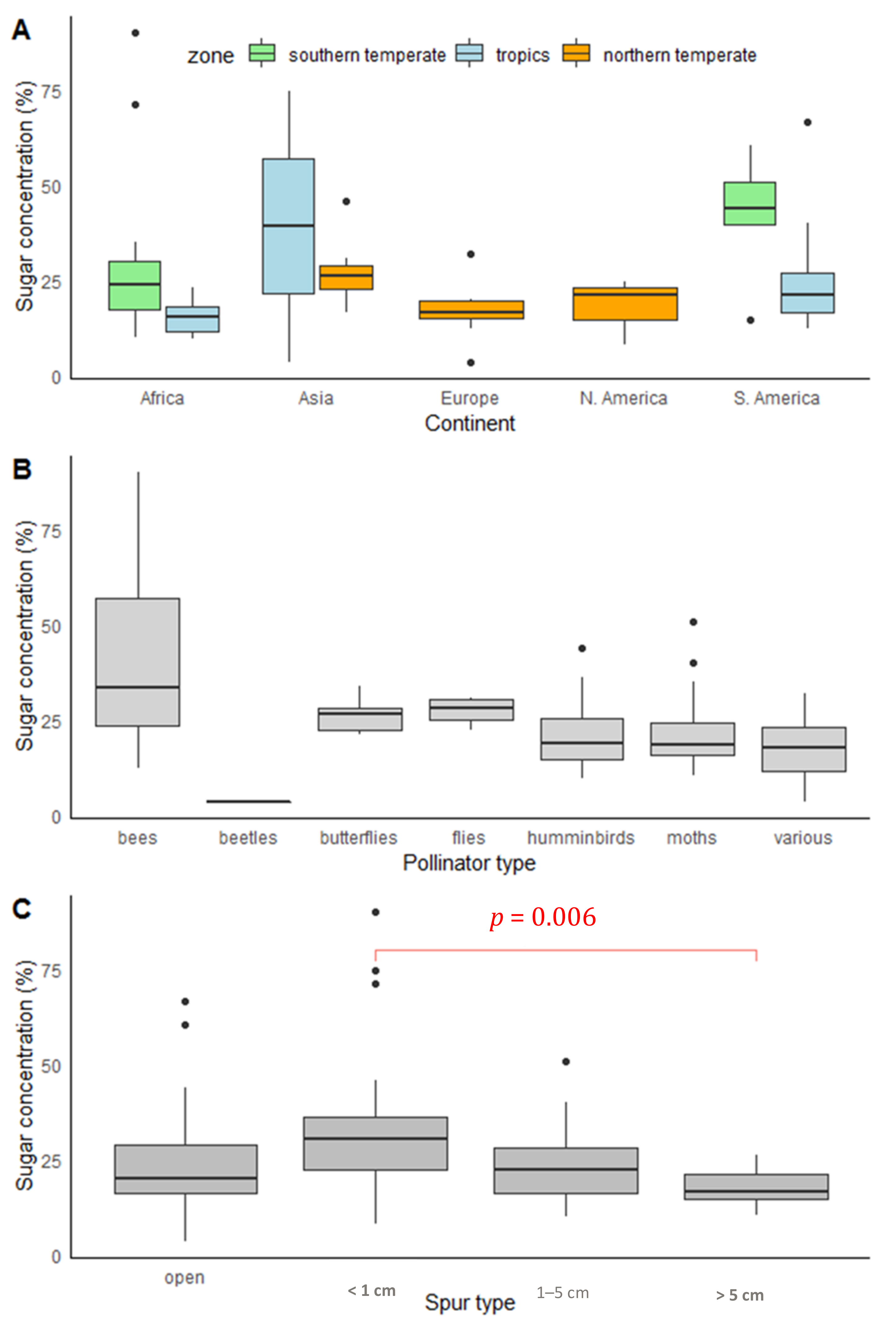
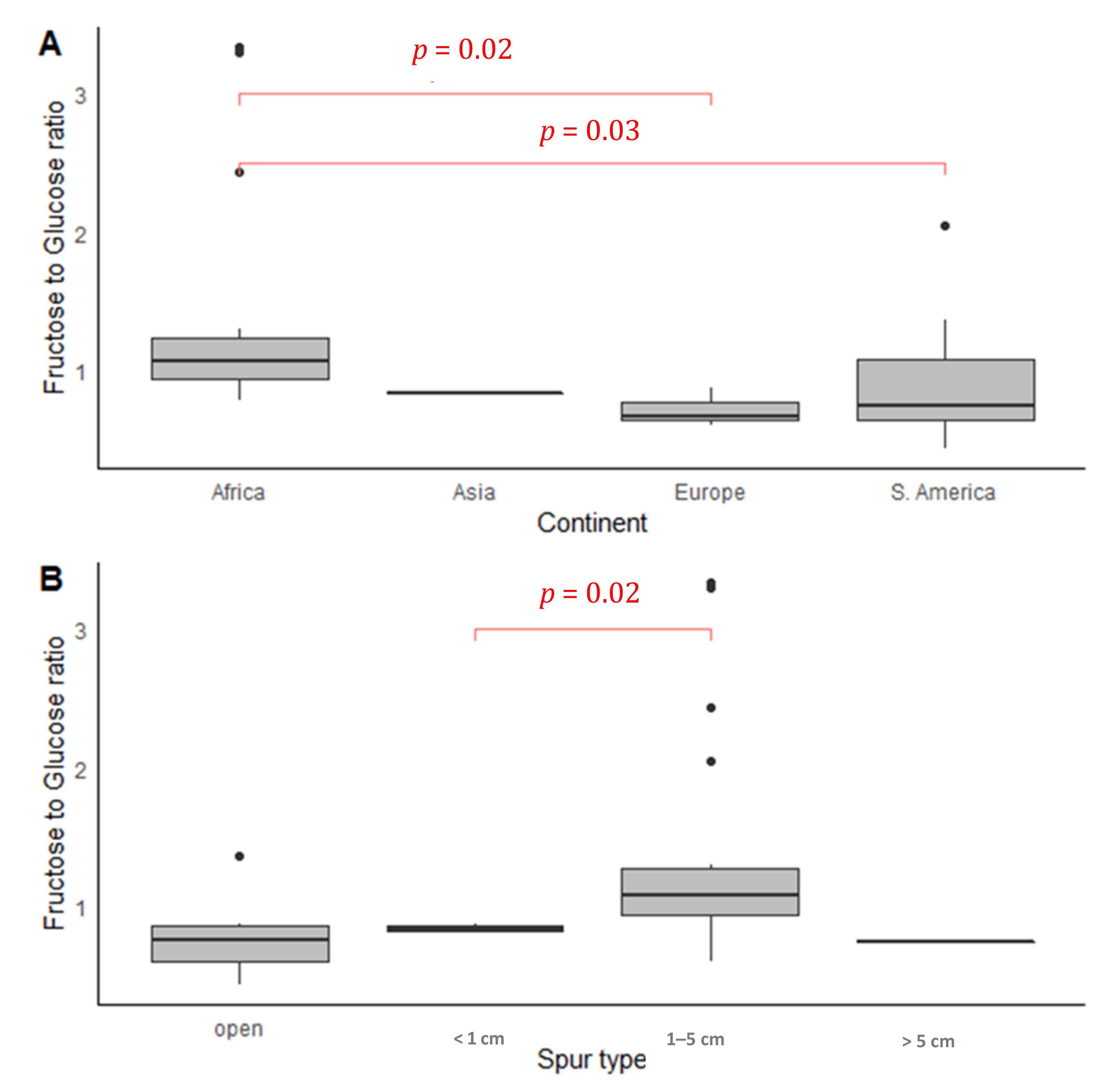
2.3. Geographical Patterns
2.4. Pollinators
2.5. Sugar Concentration According to the Life Form of the Orchids
2.6. Nectar Presentation
2.7. Nectar as Taxonomic Trait
3. Discussion
3.1. Spatial Variation in Nectar Traits
3.2. What Kinds of Nectar Do Orchid Pollinators Prefer?
3.3. Nectar Presentation
3.4. Taxonomy of Nectar
4. Materials and Methods
4.1. Data Collection and Selection
4.2. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ackerman, J.D. Mechanisms and evolution of food-deceptive pollination systems in orchids. Lindleyana 1986, 1, 108–113. [Google Scholar]
- Cozzolino, S.; Widmer, A. Orchid diversity: An evolutionary consequence of deception? Trends Ecol. Evol. 2005, 20, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, R.L.; Ackerman, J.D.; Zimmerman, J.K.; Calvo, R.N. Variation in sexual reproduction in orchids and its evolutionary consequences: A spasmodic journey to diversification. Biol. J. Lin. Soc. 2005, 84, 1–54. [Google Scholar] [CrossRef]
- Renner, S.S. Rewardless flowers in the angiosperms and the role of insect cognition in their evolution. In Plant-Pollinator Interactions; Waser, N.M., Ollerton, J., Eds.; University of Chicago Press: Chicago, IL, USA, 2006; pp. 123–144. [Google Scholar]
- Shrestha, M.; Dyer, A.G.; Dorin, A.; Ren, Z.-X.; Burd, M. Rewardlessness in orchids: How frequent and how rewardless? Plant Biol. J. 2020, 22, 555–561. [Google Scholar] [CrossRef]
- Dressler, R.L. The Orchids: Natural History and Classification; Harvard University Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Ackerman, J.D.; Rodriguez-Robles, J.A.; Melendez, E.J. A meager nectar offering by an epiphytic orchid is better than nothing. Biotropica 1994, 26, 44–49. [Google Scholar] [CrossRef]
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671. [Google Scholar] [CrossRef]
- Gijbels, P.; Van den Ende, W.; Honnay, O. Landscape scale variation in nectar amino acid and sugar composition in a Lepidoptera pollinated orchid species and its relation with fruit set. J. Ecol. 2014, 102, 136–144. [Google Scholar] [CrossRef]
- Gijbels, P.; Ceulemans, T.; Van den Ende, W.; Honnay, O. Experimental fertilization increases amino acid content in floral nectar, fruit set and degree of selfing in the orchid Gymnadenia conopsea. Oecologia 2015, 179, 785–795. [Google Scholar] [CrossRef]
- Gijbels, P.; Van den Ende, W.; Honnay, O. Phenotypic selection on nectar amino acid composition in the Lepidoptera pollinated orchid species Gymnadenia conopsea. Oikos 2015, 124, 421–427. [Google Scholar] [CrossRef]
- Brzosko, E.; Bajguz, A. Nectar composition in moth-pollinated Platanthera bifolia and P. chlorantha and its importance for reproductive success. Planta 2019, 250, 263–279. [Google Scholar] [CrossRef]
- Parachnowitsch, A.L.; Manson, J.S.; Sletvold, N. Evolutionary ecology of nectar. Ann. Bot. 2019, 123, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.D.; Brown, M. Transfer of pollinaria on bird’s feet: A new pollination system in orchids. Plant Syst. Evol. 2004, 244, 181–188. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D. Lack of floral nectar reduces self-pollination in a fly-pollinated orchid. Oecologia 2006, 147, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P.; Pupin, A.C. Effect of nectar supplementation on male and female components of pollination success in the deceptive orchid Dactylorhiza sambucina. Acta Oecol. 2008, 33, 300–306. [Google Scholar] [CrossRef]
- Tong, Z.Y.; Wang, X.P.; Wu, L.Y.; Huang, S.Q. Nectar supplementation changes pollinator behaviour and pollination mode in Pedicularis dichotoma: Implications for evolutionary transitions. Ann. Bot. 2019, 123, 373–380. [Google Scholar] [CrossRef]
- Wasserthal, L.T. The Pollinators of the Malagasy Star Orchids Andraecum sesquipedale, A. sororium and A. compactum and the Evolution of Extremely Long Spurs by Pollinator Shift. Bot. Acta. 1997, 110, 343–430. [Google Scholar] [CrossRef]
- Stpiczyńska, M. Nectar production characteristics and nectar chemistry of Platanthera chlorantha (Custer) Rchb. (Orchidaceae). Acta Hortic. 2001, 561, 137–140. [Google Scholar] [CrossRef]
- Petanidou, T. Sugars in Mediterranean floral nectars: An ecological and evolutionary approach. J. Chem. Ecol. 2005, 31, 1065–1088. [Google Scholar] [CrossRef]
- Nicolson, S.W.; Thornburg, R.W. Nectar chemistry. In Nectaries and Nectar; Springer: Dordrecht, The Netherlands, 2007; pp. 215–264. [Google Scholar]
- Nocentini, D.; Pacini, E.; Guarnieri, M.; Martelli, D.; Nepi, M. Intrapopulation heterogeneity in floral nectar attributes and foraging insects of an ecotonal Mediterranean species. Plant Ecol. 2013, 214, 799–809. [Google Scholar] [CrossRef]
- Roy, R.; Schmitt, A.J.; Thomas, J.B.; Carter, C.J. Review: Nectar biology: From molecules to ecosystems. Plant Sci. 2017, 262, 148–164. [Google Scholar] [CrossRef] [PubMed]
- Zych, M.; Junker, R.R.; Nepi, M.; Stpiczyńska, M.; Stolarska, B.; Roguz, K. Spatiotemporal variation in the pollination systems of a supergeneralist plant: Is Angelica sylvestris (Apiaceae) locally adapted to its most effective pollinators? Ann. Bot. 2018, 123, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Göttlinger, T.; Lohaus, G. Influence of light, dark, temperature and drought on metabolite and ion composition in nectar and nectaries of an epiphytic bromeliad species (Aechmea fasciata). Plant Biol. 2020, 22, 781–793. [Google Scholar] [CrossRef]
- Pacini, E.; Nepi, M.; Vesprini, J.L. Nectar biodiversity: A short review. Plant Syst. Evol. 2003, 238, 7–21. [Google Scholar] [CrossRef]
- Galetto, L.; Bernardello, G.; Rivera, G.L. Nectar, nectaries, flower visitors, and breeding system in five terrestrial Orchidaceae from central Argentina. J. Plant Res. 1997, 110, 393–403. [Google Scholar] [CrossRef]
- Petanidou, T.; Van Laere, A.; Ellis, W.N.; Smets, E. What shapes amino acid and sugar composition in Mediterranean floral nectars? Oikos 2006, 115, 155–169. [Google Scholar] [CrossRef]
- Brzosko, E.; Bajguz, A.; Chmur, M.; Burzyńska, J.; Jermakowicz, E.; Mirski, P.; Zieliński, P. How Are the Flower Structure and Nectar Composition of the Generalistic Orchid Neottia ovata Adapted to a Wide Range of Pollinators? Int. J. Mol. Sci. 2021, 22, 2214. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.G.; Baker, I. Floral nectar sugar constituents in relation to pollinator type. In Handbook of Experimental Pollination Biology; Jones, C.E., Little, R.J., Eds.; Van Nostrand Reinhold Company Inc.: New York, NY, USA, 1983; pp. 131–141. [Google Scholar]
- Willmer, P. Pollination by butterflies and moths. In Pollination and Floral Ecology; Willmer, P., Ed.; Princeton University Press: Princeton, NJ, USA, 2011; pp. 322–336. [Google Scholar] [CrossRef]
- Baker, H.G.; Baker, I. The predictive value of nectar chemistry to the recognition of pollinator types. Isr. J. Plant Sci. 1990, 39, 157–166. [Google Scholar] [CrossRef]
- Pyke, G.H. Plant-pollinator co-evolution: It’s time to reconnect with Optimal Foraging Theory and Evolutionarily Stable Strategies. Persp. Plant Ecol. Evol. Syst. 2016, 19, 70–76. [Google Scholar] [CrossRef]
- Abrahamczyk, S.; Kessler, M.; Hanley, D.; Karger, D.N.; Müller, M.P.J.; Knauer, A.C.; Keller, F.; Schwerdtfeger, M.; Humphreys, A.M. Pollinator adaptation and the evolution of floral nectar sugar composition. J. Evol. Biol. 2017, 30, 112–127. [Google Scholar] [CrossRef]
- Heil, M. Nectar: Generation, regulation and ecological functions. Trends Plant Sci. 2011, 16, 191–200. [Google Scholar] [CrossRef]
- Gardener, M.C.; Gillman, M.P. The taste of nectar–a neglected area of pollination ecology. Oikos 2002, 98, 552–557. [Google Scholar] [CrossRef]
- Zhang, W.L.; Gao, J.Y. Multiple factors contribute to reproductive isolation between two co-existing Habenaria species (Orchidaceae). PLoS ONE 2017, 12, e0188594. [Google Scholar] [CrossRef] [PubMed]
- Pais, M.S.; Figueiredo, M.C.S. Floral nectaries from Limodorum abortivum (L.) Sw and Epipactis atropurpurea Rafin (Orchidaceae: Ultrastructural changes in plastids during the secretory processes. Apidologie 1994, 25, 615–626. [Google Scholar] [CrossRef]
- Vandelook, F.; Janssens, S.B.; Gijbels, P.; Fischer, E.; Van den Ende, W.; Honnay, O.; Abrahamczyk, S. Nectar traits differ between pollination syndromes in Balsaminaceae. Ann. Bot. 2019, 124, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Percival, M.S. Types of nectar in angiosperms. New Phytol. 1961, 60, 235–281. [Google Scholar] [CrossRef]
- Gottsberger, G.; Schrauwen, J.; Linskens, H.F. Amino Acids and Sugars in nectar, and Their Putative Evolutionary Significance. Plant Syst. Evol. 1984, 145, 55–77. [Google Scholar] [CrossRef]
- Pais, M.S.S.; Neves, H.C.D.; Vasconcelos, A.M.P. Amino acid and sugar content of the nectar exudate from Limodorum abortivum (Orchidaceae). Comparison with Epipactis atropurpurea nectar composition. Apidologie 1986, 17, 125–136. [Google Scholar] [CrossRef]
- Jeffrey, D.C.; Arditti, J.; Koopowitz, H. Sugar content in floral and extrafloral exudates of orchids: Pollination, myrmecology and chemotaxonomy implication. New Phytol. 1970, 69, 187–195. [Google Scholar] [CrossRef]
- Bernardello, G.; Galetto, L.; Forcone, A. Floral nectar chemical composition of some species from Patagonia. II. Biochem. Syst. Ecol. 1999, 27, 779–790. [Google Scholar] [CrossRef]
- Krömer, T.; Kessler, M.; Lohaus, G.; Schmidt-Lebuhn, A.N. Nectar sugar composition and concentration in relation to pollination syndromes in Bromeliaceae. Plant Biol. 2008, 10, 502–511. [Google Scholar] [CrossRef]
- Witt, T.; Jürgens, A.; Gottsberg, G. Nectar sugar composition of European Caryophylloideae (Caryophyllaceae) in relation to flower length, pollination biology and phylogeny. J. Evol. Biol. 2013, 26, 2244–2259. [Google Scholar] [CrossRef]
- Wolff, D. Nectar Sugar Composition and Volumes of 47 Species of Gentianales from a Southern Ecuadorian Montane Forest. Ann. Bot. 2006, 97, 767–777. [Google Scholar] [CrossRef]
- Claessens, J.; Bacallado, J.J.; Bogarin, D.; Dedroog, L.; Heijungs, R.; Langelaan, R.; van Nieukerken, E.J.; van den Berg, K.; Gravendeel, B. Pollination of Habenaria tridactylites on the Canary Islands. Nord. J. Bot. 2019, e02401. [Google Scholar] [CrossRef]
- Chalcoff, V.R.; Gleiser, G.; Ezcurra, C.; Aizen, M.A. Pollinator type and secondarily climate are related to nectar sugar composition across the angiosperms. Evol. Ecol. 2017, 31, 585–602. [Google Scholar] [CrossRef]
- Galetto, L.; Bernardello, G. Nectar sugar composition in angiosperms from Chaco and Patagonia (Argentina): An animal visitor’s matter? Plant Syst. Evol. 2003, 238, 69–86. [Google Scholar] [CrossRef]
- Chalcoff, V.R.; Aizen, M.A.; Galetto, L. Nectar Concentration and Composition of 26 Species from the Temperate Forest of South America. Ann. Bot. 2006, 97, 413–421. [Google Scholar] [CrossRef]
- Nilsson, L.A.; Rabakonandrianina, E. Hawk-moth scale analysis and pollination specialization in the epilithic Malagasy endemic Aerangis ellisii (Reichenb. fiI.) Schltr. (Orchidaceae). Bot. J. Linn. Soc. 1988, 97, 49–61. [Google Scholar] [CrossRef]
- Rodríguez-Robles, J.A.; Meléndez, E.J.; Ackerman, J.D. Effects of Display Size, Flowering Phenology, and Nectar Availability on Effective Visitation Frequency in Comparettia falcata (Orchidaceae). Amer. J. Bot. 1992, 79, 1009–1017. [Google Scholar] [CrossRef]
- Neubig, K.M.; Carlsward, B.S.; Whitten, W.M.; Williams, N.H. Nectary structure and nectar in Sobralia and Elleanthus (Sobralieae: Orchidaceae). Lankesteriana 2015, 15, 113–127. [Google Scholar] [CrossRef]
- Pansarin, E.R.; Ferreira, A.W.C. Butterfly pollination in Pteroglossa (Orchidaceae, Orchidoideae): A comparative study on the reproductive biology of two species of a Neotropical genus of Spiranthinae. J. Plant Res. 2015, 128, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Pamminger, T.; Becker, R.; Himmelreich, S.; Schneider, C.W.; Bergtold, M. The nectar report: Quantitative review of nectar sugar concentrations offered by bee visited flowers in agricultural and non-agricultural landscapes. PeerJ 2019, 7, e6329. [Google Scholar] [CrossRef]
- Pyke, G.H.; Waser, N.M. The production of dilute nectars by hummingbird and honeyeater flowers. Biotropica 1981, 13, 260–270. [Google Scholar] [CrossRef]
- Perret, M.; Chautems, A.; Spichiger, R.; Peixoto, M.; Savolainen, V. Nectar Sugar Composition in Relation to Pollination Syndromes in Sinningieae (Gesneriaceae). Ann. Bot. 2001, 87, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Gilet, T.; Bush, J.W. Optimal concentrations in nectar feeding. Proc. Natl. Acad. Sci. USA 2011, 108, 16618–16621. [Google Scholar] [CrossRef]
- Tiedge, K.; Lohaus, G. Nectar sugars and amino acids in day and night-flowering Nicotiana species are more strongly shaped by pollinators’ preferences than organic acids and inorganic ions. PLoS ONE 2017, 12, e0176865. [Google Scholar] [CrossRef]
- Josens, R.B.; Farina, W.M. Nectar feeding by the hovering hawk moth Macroglossum stellatarum: Intake rate as a function of viscosity and concentration of sucrose solutions. J. Comp. Physiol. A 2001, 187, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Heyneman, A.J. Optimal sugar concentrations of floral nectars–Dependence on sugar intake efficiency and foraging costs. Oecologia 1983, 60, 198–213. [Google Scholar] [CrossRef]
- Nicolson, S.W. Amino acid concentrations in the nectars of southern African bird-pollinated flowers, especially Aloe and Erythrina. J. Chem. Ecol. 2007, 33, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, S.W.; Fleming, P.A. Nectar as food for birds: The physiological consequences of drinking dilute sugar solutions. Plant Syst. Evol. 2003, 238, 139–153. [Google Scholar] [CrossRef]
- Peter, C.I.; Johnson, S.D. Reproductive biology of Acrolophia cochlearis (Orchidaceae): Estimating rates of cross-pollination in epidendroid orchids. Ann. Bot. 2009, 104, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.D.; Nicolson, S.W. Evolutionary associations between nectar properties and specificity in bird pollination systems. Biol. Lett. 2008, 4, 49–52. [Google Scholar] [CrossRef]
- Nilsson, L.A. Deep flowers for long tongues: Reply from L.A. Nilsson. Trends Ecol. Evol. 1998, 13, 509. [Google Scholar] [CrossRef]
- Armbruster, W.S. Plant-pollinator interactions from flower to landscape. The specialization continuum in pollination systems: Diversity of concepts and implications for ecology, evolution and conservation. Funct. Ecol. 2017, 31, 88–100. [Google Scholar] [CrossRef]
- Nepi, M. Beyond nectar sweetness: The hidden ecological role of non-protein amino acids in nectar. J. Ecol. 2014, 102, 108–115. [Google Scholar] [CrossRef]
- Reiter, N.; Bohman, B.; Freestone, M.; Brown, G.R.; Phillips, R.D. Pollination by nectar foraging thynnine wasps in the endangered Caladenia arenaria and Caladenia concolor (Orchidaceae). Austr. J. Bot. 2019, 67, 490–500. [Google Scholar] [CrossRef]
- Phillips, R.D.; Bohman, B.; Brown, G.R.; Tomlinson, S.; Peakall, R. A specialised pollination system using nectar-seeking thynnine wasps in Caladenia nobilis (Orchidaceae). Plant Biol. 2020, 22, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.E.P.; Castro, M.D.; Galetto, L.; Sazima, M. Anatomy of the floral nectary of ornithophilous Elleanthus brasiliensis (Orchidaceae: Sobralieae). Bot. J. Linn. Soc. 2013, 171, 764–772. [Google Scholar] [CrossRef]
- Jürgens, A.; Bosch, S.R.; Webber, A.C.; Witt, T.; Frame, D.; Gottsberger, G. Pollination biology of Eulophia alta (Orchidaceae) in Amazonia: Effects of pollinator composition on reproductive success in different populations. Ann. Bot. 2009, 104, 897–912. [Google Scholar] [CrossRef]
- Davies, K.L.; Stpiczyńska, M.; Gregg, A. Nectar-secreting Floral Stomata in Maxillaria anceps Ames & C. Schweinf. (Orchidaceae). Ann. Bot. 2005, 96, 217–227. [Google Scholar] [CrossRef]
- Hodges, S.A. The influence of nectar production on hawkmoth behavior, self pollination, and seed production in Mirabilis multiflora (Nyctaginaceae). Am. J. Bot. 1995, 82, 197–204. [Google Scholar] [CrossRef]
- Maad, J.; Nilsson, L.A. On the mechanism of floral shifts in speciation: Gained pollination efficiency from tongue- to eye-attachment of pollinia in Platanthera (Orchidaceae). Biol. J. Linn. Soc. 2004, 83, 481–495. [Google Scholar] [CrossRef]
- Boberg, E.; Ägren, J. Despite their apparent integration, spur length but not perianth size affects reproductive success in the moth-pollinated orchid Platanthera bifolia. Funct Ecol. 2009, 23, 1022–1028. [Google Scholar] [CrossRef]
- Boberg, E.; Alexandersson, R.; Jonsson, M.; Maad, J.; Ägren, J.; Nilsson, L.A. Pollinator shifts and the evolution of spur length in the moth-pollinated orchid Platanthera bifolia. Ann. Bot. 2014, 113, 267–275. [Google Scholar] [CrossRef]
- Roberts, D.L. Observations on the effect of nectar-robbery on the reproductive success of Aeranthes arachnitis (Orchidaceae). Lankesteriana 2007, 7, 509–514. [Google Scholar]
- Martins, D.; Johnson, S.D. Hawkmoth pollination of aerangoid orchids in Kenya, with special reference to nectar sugar concentration gradients in the floral spurs. Am. J. Bot. 2007, 94, 650–659. [Google Scholar] [CrossRef]
- Koopowitz, H.; Marchant, T.A. Postpollination nectar reabsorption in the African epiphyte Aerangis verdickii (Orchidaceae). Am. J. Bot. 1998, 85, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.A.; Jonsson, L.; Rason, L.; Randrianjohany, E. Monophily and pollination mechanisms in Angraecum arachnites Schltr. (Orchidaceae) in a guild of long-tongued hawk-moths (Sphingidae) in Madagascar. Biol. J. Linn. Soc. 1985, 26, 1–19. [Google Scholar] [CrossRef]
- Johnson, S.D.; Raguso, R.A. The long-tongued hawkmoth pollinator niche for native and invasive plants in Africa. Ann. Bot. 2016, 117, 25–36. [Google Scholar] [CrossRef]
- Micheneau, C.; Fournel, J.; Pailler, T. Bird Pollination in an Angraecoid Orchid on Reunion Island (Mascarene Archipelago, Indian Ocean). Ann. Bot. 2006, 97, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Micheneau, C.; Fournel, J.; Humeau, L.; Pailler, T. Orchid bird interactions: A case study from Angraecum (Vandeae, Angraecinae) and Zosterops (white-eyes, Zosteropidae) on Reunion Island. Botany 2008, 86, 1143–1151. [Google Scholar] [CrossRef]
- Balducci, M.G.; Van der Niet, T.; Johnson, S.D. Butterfly pollination of Bonatea cassidea (Orchidaceae): Solving a puzzle from the Darwin era. South Afr. J. Bot. 2019, 123, 308–316. [Google Scholar] [CrossRef]
- Balducci, M.G.; Van der Niet, T.; Johnson, S.D. Diel scent and nectar rhythms of an African orchid in relation to bimodal activity patterns of hawkmoth pollinators. Annals Bot. 2020, 126, 1155–1164. [Google Scholar] [CrossRef]
- Johnson, S.D.; Liltved, W. Hawkmoth pollination of Bonatea speciosa (Orchidaceae) in a South African coastal forest. Nord. J. Bot. 1997, 17, 5–10. [Google Scholar] [CrossRef]
- Ponsie, M.E.; Edwards, T.J.; Johnson, S.D. A taxonomic revision of Bonatea Willd. (Orchidaceae: Orchidoideae: Habenariinae). South Afr. J. Bot. 2007, 73, 1–21. [Google Scholar] [CrossRef]
- Larsen, M.W.; Peter, C.; Johnson, S.D.; Olesen, J.M. Comparative biology of pollination systems in the African-Malagasy genus Brownleea (Brownleeinae: Orchidaceae). Bot. J. Linn. Soc. 2008, 156, 65–78. [Google Scholar] [CrossRef]
- Reiter, N.; Bohman, B.; Flematti, G.R.; Phillips, R.D. Pollination by nectar-foraging thynnine wasps: Evidence of a new specialized pollination system for Australian orchids. Bot. J. Linn. Soc. 2018, 188, 327–337. [Google Scholar] [CrossRef]
- Reiter, N.; Bohman, B.; Batley, M.; Phillips, R.D. Pollination of an endangered Caladenia species (Orchidaceae) by nectar-foraging behaviour of a widespread species of colletid bee. Bot. J. Linn. Soc. 2019, 189, 83–98. [Google Scholar] [CrossRef]
- Ponert, J.; Trávníček, P.; Vuong, T.B.; Rybková, R.; Suda, J. A New Species of Cleisostoma (Orchidaceae) from the Hon Ba Nature Reserve in Vietnam: A Multidisciplinary Assessment. PLoS ONE 2016, 11, e0150631. [Google Scholar] [CrossRef]
- Nilsson, L.A.; Rabakonandrianina, E.; Razananaivo, R.; Randriamanindry, J.J. Long pollinia on eyes: Hawk-moth pollination of Cynorkis uniflora Lindley (Orchidaceae) in Madagascar. Bot. J. Linn. Soc. 1992, 109, 145–160. [Google Scholar] [CrossRef]
- Luyt, R.P. Pollination and Evolution of the genus Mystacidium (Orchidaceae). Master of Science Thesis, School of Botany and Zoology, University of Natal, Pietermaritzburg, South Africa, 2002. [Google Scholar]
- Johnson, S.D. Observations of hawkmoth pollination in the South African orchid Disa cooperi. Nord. J. Bot. 1995, 15, 121–125. [Google Scholar] [CrossRef]
- Johnson, S.D. Pollination by long-proboscid flies in the endangered African orchid Disa scullyi. South Afr. J. Bot. 2006, 72, 24–27. [Google Scholar] [CrossRef][Green Version]
- Pais, M.S.S.; Neves, H.C.D. Sugar content of the nectary exudate of Epipactis atropurpurea Rafin. Apidologie 1980, 11, 39–45. [Google Scholar] [CrossRef]
- Sun, M.; Gross, K.; Schiestl, F.P. Floral adaptation to local pollinator guilds in a terrestrial orchid. Ann. Bot. 2014, 113, 289–300. [Google Scholar] [CrossRef]
- Xiong, Y.Z.; Jia, L.B.; Liu, C.Q.; Huang, S.Q. Effective pollinia transfer by settling moths’ legs in an orchid Habenaria aitchisonii: Pollinia transfer by moth’s legs. J. Syst. Evol. 2019, 58, 174–181. [Google Scholar] [CrossRef]
- Xiong, Y.Z.; Liu, C.Q.; Huang, S.Q. Mast fruiting in a hawkmoth pollinated orchid Habenaria glaucifolia: An 8-year survey. J. Plant Ecol. 2015, 8, 136–141. [Google Scholar] [CrossRef]
- Singer, R.; Cocucci, A.A. Eye attached hemipollinaria in the hawkmoth and settling moth pollination of Habenaria (Orchidaceae): A study on functional morphology in 5 species from subtropical South America. Bot. Acta. 1997, 110, 328–337. [Google Scholar] [CrossRef]
- Amorim, F.W.; Wyatt, G.E.; Sazima, M. Low abundance of long-tongued pollinators leads to pollen limitation in four specialized hawkmoth-pollinated plants in the Atlantic Rain forest, Brazil. Naturwissenschaften 2014, 101, 893–905. [Google Scholar] [CrossRef]
- Pedron, M.; Buzatto, C.R.; Singer, R.B.; Batista, J.A.N.; Moser, A. Pollination biology of four sympatric species of Habenaria (Orchidaceae: Orchidinae) from southern Brazil. Bot. J. Linn. Soc. 2012, 170, 141–156. [Google Scholar] [CrossRef]
- Tao, Z.B.; Ren, Z.X.; Bernhardt, P.; Wang, W.J.; Liang, H.; Li, H.D.; Wang, H. Nocturnal hawkmoth and noctuid moth pollination of Habenaria limprichtii (Orchidaceae) in sub-alpine meadows of the Yulong Snow Mountain (Yunnan, China). Bot. J. Linn. Soc. 2018, 187, 483–498. [Google Scholar] [CrossRef]
- Singer, R.B.; Breier, T.B.; Flach, A.; Farias-Singer, R. The Pollination Mechanism of Habenaria pleiophylla Hoehne & Schlechter (Orchidaceae: Orchidinae). Funct. Ecosyst. Comm. 2007, 1, 10–14. [Google Scholar]
- Chen, X.H.; Tan, S.T.; Liang, Y.L.; Huang, L.; Xiao, H.W.; Luo, H.L.; Xiong, D.J.; Yang, B.Y.; Ren, Z.X. The pollination of Habenaria rhodocheila (Orchidaceae) in South China: When butterflies take sides. Ecol. Evol. 2021, 11, 2849–2861. [Google Scholar] [CrossRef] [PubMed]
- Micheneau, C.; Fournel, J.; Gauvin-Bialecki, A.; Pailler, T. Auto-pollination in a long-spurred endemic_orchid Jumellea stenophylla on Reunion Island Mascarene Archipelago Indian Ocean. Plant Syst. Evol. 2008, 272, 11–22. [Google Scholar] [CrossRef]
- Vakhrameeva, M.G.; Tatarenko, I.V.; Varlygina, T.I.; Torosyan, G.K.; Zagulski, M.N. Orchids of Russia and Adjacent Countries (within the Borders of the Former USSR); A.R.G. Gantner: Ruggell, Liechtenstein, 2008; p. 690. [Google Scholar]
- Peter, C.I.; Venter, N. Generalist, settling moth pollination in the endemic South African twig epiphyte, Mystacidium pusillum Harv. (Orchidaceae). Flora 2016, 232, 16–21. [Google Scholar] [CrossRef]
- Luyt, R.; Johnson, S. Hawkmoth pollination of the African epiphytic orchid Mystacidium venosum, with special reference to flower and pollen longevity. Plant Syst. Evol. 2001, 228, 49–62. [Google Scholar] [CrossRef]
- Aguiar, J.M.R.B.V.; Pansarin, E.R. Does Oeceoclades maculata (Orchidaceae) reabsorb nectar? Eur. J. Env. Sci. 2013, 3, 113–118. [Google Scholar] [CrossRef][Green Version]
- Gonzales-Dias, N.; Ackerman, J.D. Pollination, fruit set, and seed production in the orchid, Oeceoclades maculata. Lindleyana 1998, 3, 150–155. [Google Scholar]
- Stpiczyńska, M.; Pielecki, J. Sekrecja, resorbcja i skład chemiczny nektaru podkolana zielonawego Platanthera chlorantha (Custer) Rchb. (Orchidaceae) (Secretion, resorption and nectar chemistry of Greater Butterfly Orchid Platanthera chlorantha (Custer) Rchb. (Orchidaceae)). Ann UMCS Sectio. EEE Hortic. 2002, 10, 173–179. [Google Scholar]
- Robertson, J.L.; Wyatt, R. Reproductive biology of the yellow-fringed orchid, Platanthera ciliaris. Am. J. Bot. 1990, 77, 388–398. [Google Scholar] [CrossRef]
- Westwood, A.R.; Borkowsky, C.L.; Budnick, K.E. Seasonal variation in the nectar sugar concentration and nectar quantity in the Western Prairie Fringed Orchid, Platanthera praeclara (Orchidaceae). Rhodora 2011, 113, 201–219. [Google Scholar] [CrossRef]
- Patt, J.M.; Merchant, M.W.; Williams, D.R.E.; Meeuse, B.J.D. Pollination Biology of Platanthera stricta (Orchidaceae) in Olympic National Park, Washington. Am. J. Bot. 1989, 76, 1097–1106. [Google Scholar] [CrossRef]
- Buragohain, B.; Chaturvedi, S.K.; Puro, N. Biotic Pollination in Rhynchostylis retusa (L.) Bl. (Orchidaceae). Int. J. Plant Repr. Biol. 2015, 7, 78–83. [Google Scholar] [CrossRef]
- Carvalho, R.; Machado, I.C. Rodriguezia bahiensis Rchb. F.: Biologia floral, polinizadores e primeiro registro de polinização por moscas Acroceridae em Orchidaceae. Braz. J. Bot. 2006, 29, 461–470. [Google Scholar] [CrossRef]
- Ospina-Calderón, N.H.; Duque-Buitrago, C.A.; Tremblay, R.L.; Tupac Otero, J. Pollination ecology of Rodriguezia granadensis (Orchidaceae). Lankesteriana 2015, 15, 129–139. [Google Scholar] [CrossRef]
- Pansarin, E.R.; Bergamo, P.J.; Ferraz, L.J.C.; Pedro, S.R.M.; Ferreira, A.W.C. Comparative reproductive biology reveals two distinct pollination strategies in Neotropical twig-epiphyte orchids. Plant Syst. Evol. 2018, 304, 793–806. [Google Scholar] [CrossRef]
- Johnson, S.D. Bird pollination in South African species of Satyrium (Orchidaceae). Plant Syst. Evol. 1996, 203, 91–98. [Google Scholar] [CrossRef]
- Ellis, A.G.; Johnson, S.D. Do pollinators determine hybridization patterns in sympatric Satyrium (Orchidaceae) species? Plant Syst. Evol. 1999, 219, 137–150. [Google Scholar] [CrossRef]
- Van der Niet, T.; Jürgens, A.; Johnson, S.D. Pollinators, floral morphology and scent chemistry in the southern African orchid genus Schizochilus. South Afr. J. Bot. 2010, 76, 726–738. [Google Scholar] [CrossRef]
- Johnson, S.D.; Peter, C.I.; Ellis, A.G.; Boberg, E.; Botes, C.; van der Niet, T. Diverse pollination systems of the twin-spurred orchid genus Satyrium in African grasslands. Plant Syst. Evol. 2011, 292, 95–103. [Google Scholar] [CrossRef]
- Arakaki, N.; Yasuda, K.; Kanayama, S.; Jitsino, S.; Oike, M.; Wakamura, S. Attraction of males of the cupreous polished chafer Protaetia pryeri pryeri (Coleoptera: Scarabaeidae) for pollination by an epiphytic orchid Luisia teres (Asparagales: Orchidaceae). Appl. Entomol. Zool. 2016, 51, 241–246. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Log (Sugar Concentration) | |||
|---|---|---|---|
| Predictors | Estimates | CI | p |
| (Intercept) | 1.77 | 0.77–2.76 | 0.001 |
| Continent [Asia] | 1.16 | 0.30–2.02 | 0.009 |
| Continent [Europe] | 0.68 | −0.29–1.66 | 0.166 |
| Continent [N. America] | 0.74 | −0.29–1.78 | 0.158 |
| Continent [S. America] | 0.33 | 0.08–0.58 | 0.012 |
| Climate zone [southern temperate] | 1.06 | 0.13–1.99 | 0.027 |
| Climate zone [tropics] | 0.69 | −0.19–1.57 | 0.124 |
| Pollinators [bees] | 0.62 | 0.25–0.99 | 0.001 |
| Pollinators [beetles] | −2.41 | −3.62–−1.20 | <0.001 |
| Pollinators [butterflies] | 0.45 | −0.02–0.92 | 0.060 |
| Pollinators [flies] | 0.45 | −0.04–0.94 | 0.073 |
| Pollinators [hummingbirds] | 0.13 | −0.25–0.50 | 0.500 |
| Pollinators [moths] | 0.27 | −0.09–0.64 | 0.137 |
| Spur category [no spur, open nectaries] | 0.05 | −0.23–0.33 | 0.744 |
| Spur category [below 1 cm] | 0.07 | −0.19–0.34 | 0.576 |
| Spur category [over 5 cm] | −0.09 | −0.37–0.20 | 0.553 |
| Observations | 106 | ||
| R2/R2 adjusted | 0.491/0.406 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brzosko, E.; Mirski, P. Floral Nectar Chemistry in Orchids: A Short Review and Meta-Analysis. Plants 2021, 10, 2315. https://doi.org/10.3390/plants10112315
Brzosko E, Mirski P. Floral Nectar Chemistry in Orchids: A Short Review and Meta-Analysis. Plants. 2021; 10(11):2315. https://doi.org/10.3390/plants10112315
Chicago/Turabian StyleBrzosko, Emilia, and Paweł Mirski. 2021. "Floral Nectar Chemistry in Orchids: A Short Review and Meta-Analysis" Plants 10, no. 11: 2315. https://doi.org/10.3390/plants10112315
APA StyleBrzosko, E., & Mirski, P. (2021). Floral Nectar Chemistry in Orchids: A Short Review and Meta-Analysis. Plants, 10(11), 2315. https://doi.org/10.3390/plants10112315