
Differential Response of Two Tomato Genotypes, Wild Type cv. Ailsa Craig and Its ABA-Deficient Mutant flacca to Short-Termed Drought Cycles

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
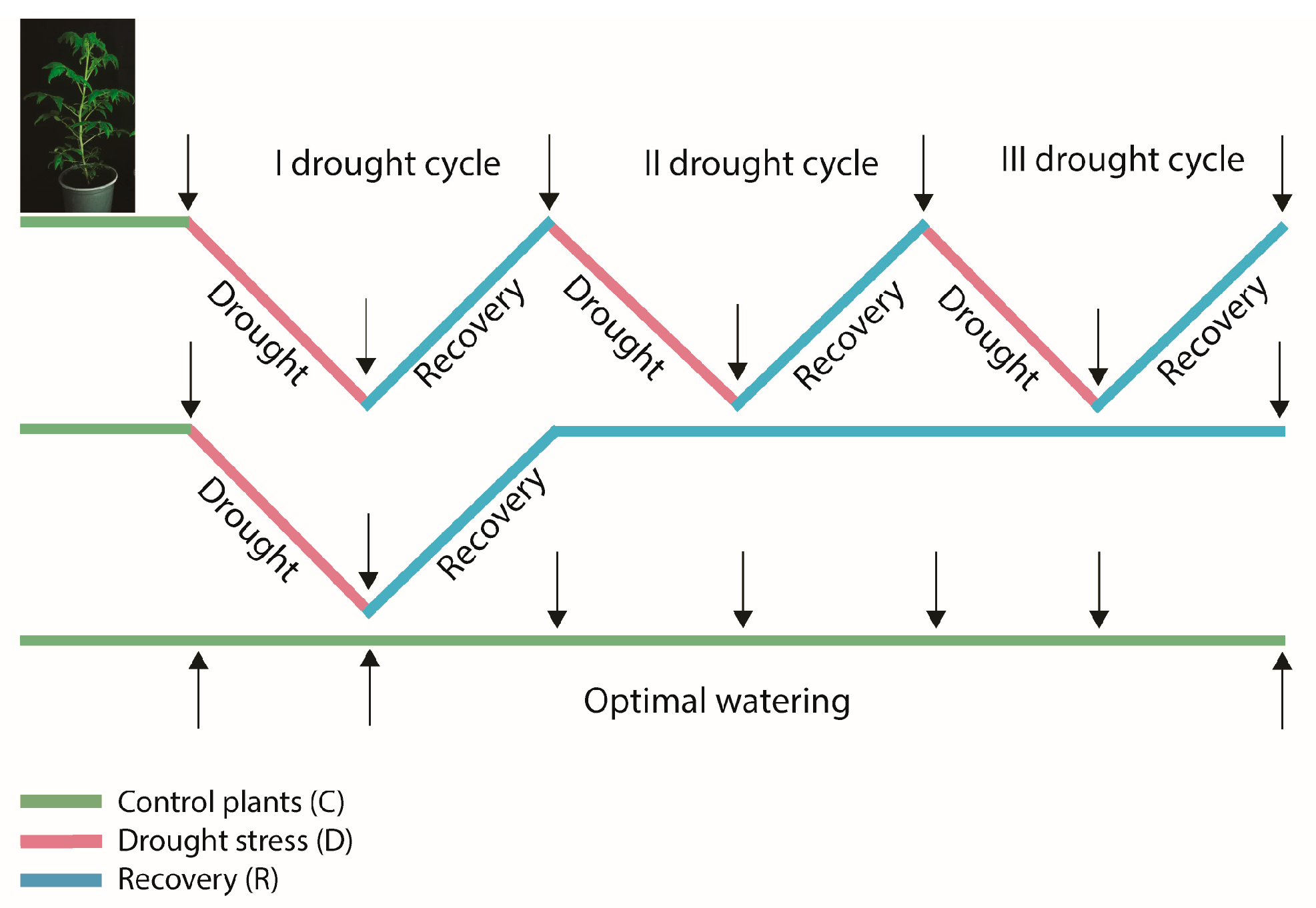
2.1. Drought Cycles
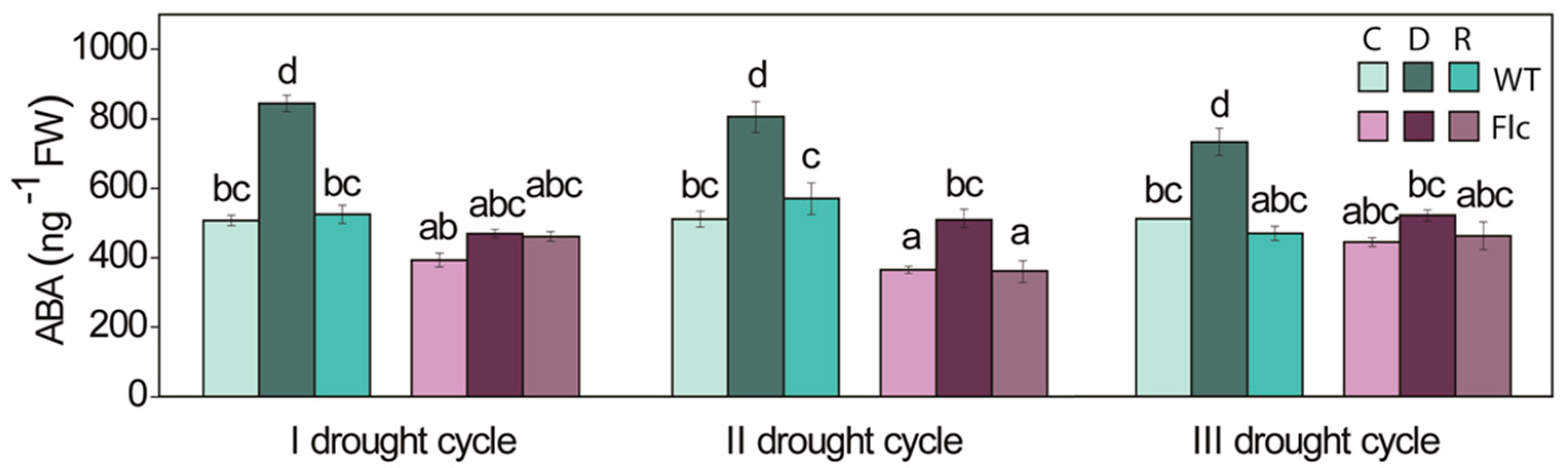
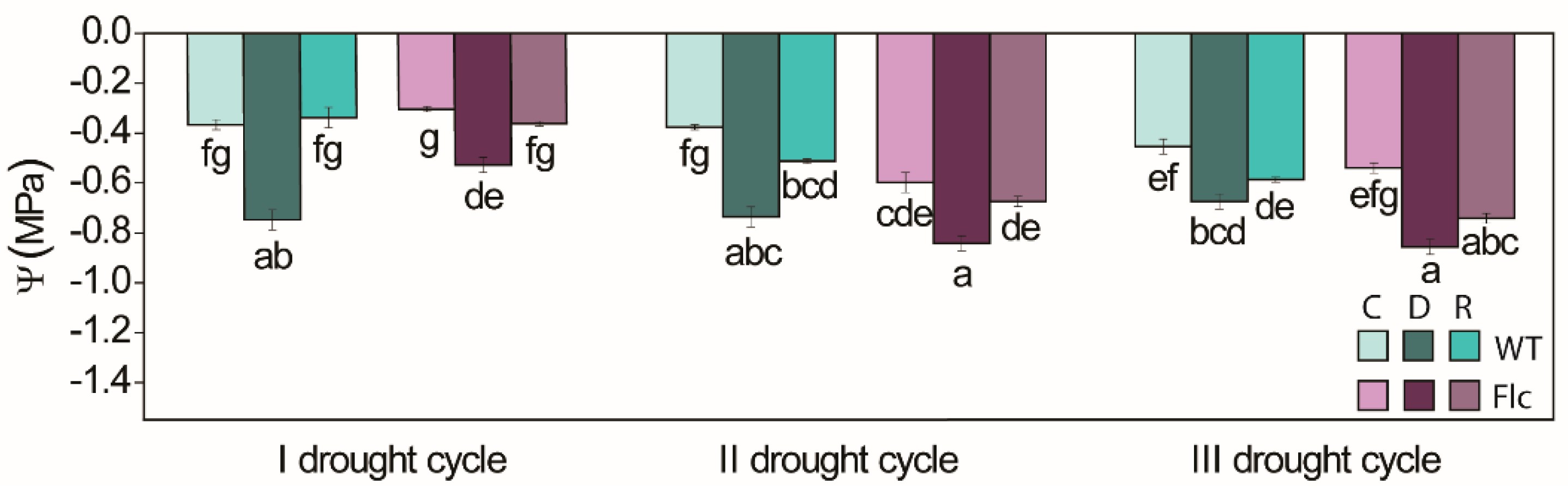
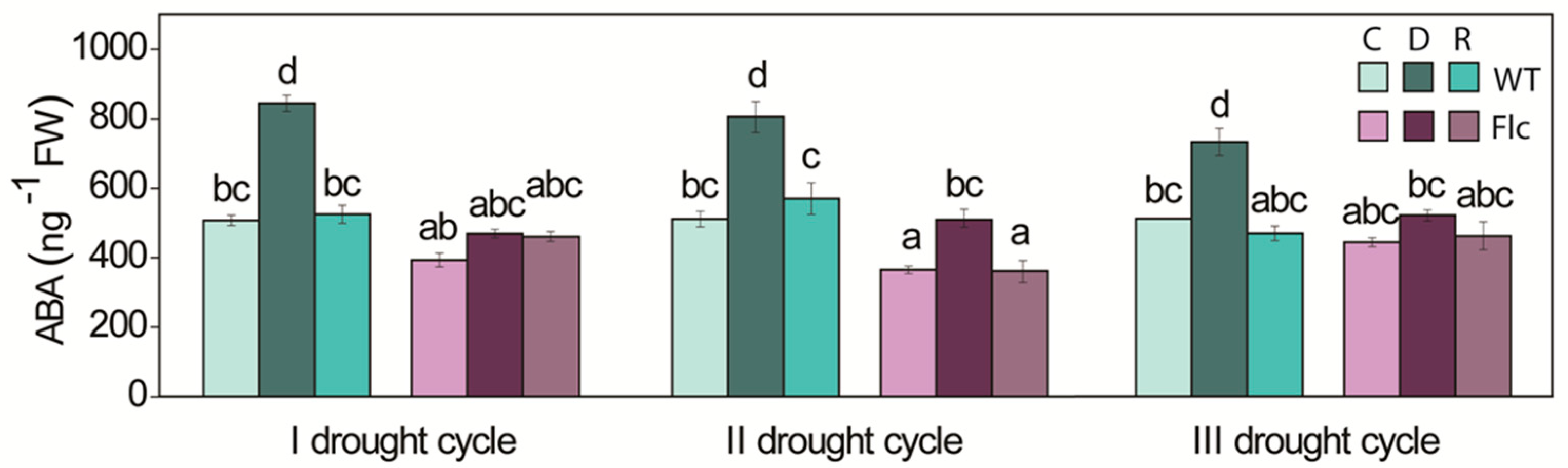
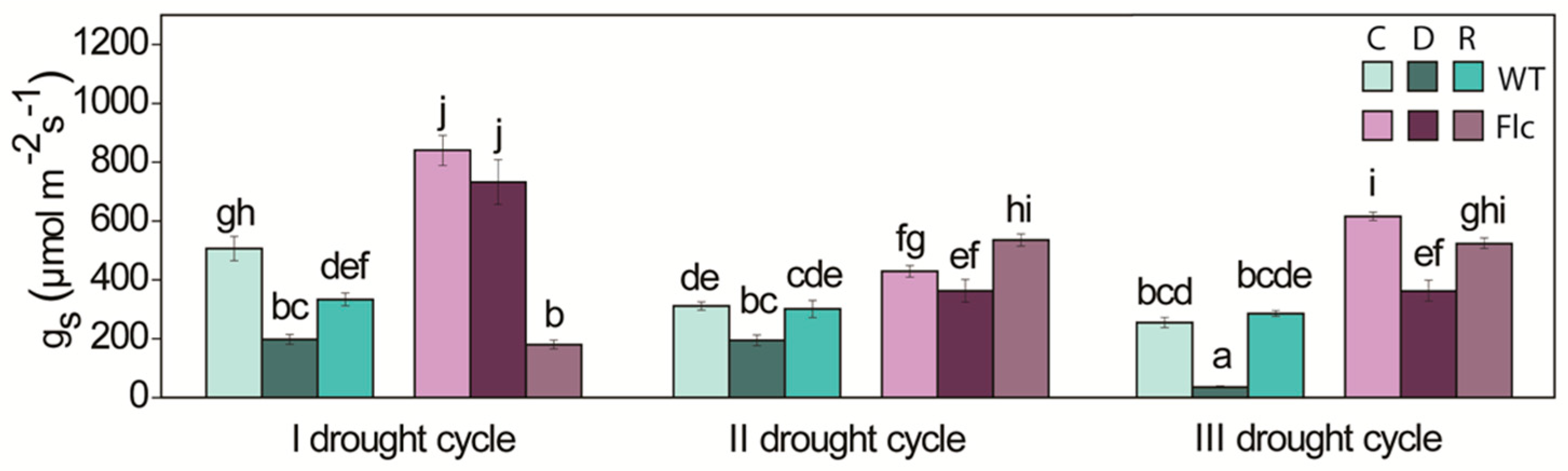
2.1.1. ABA Content, Stomata and Water Status
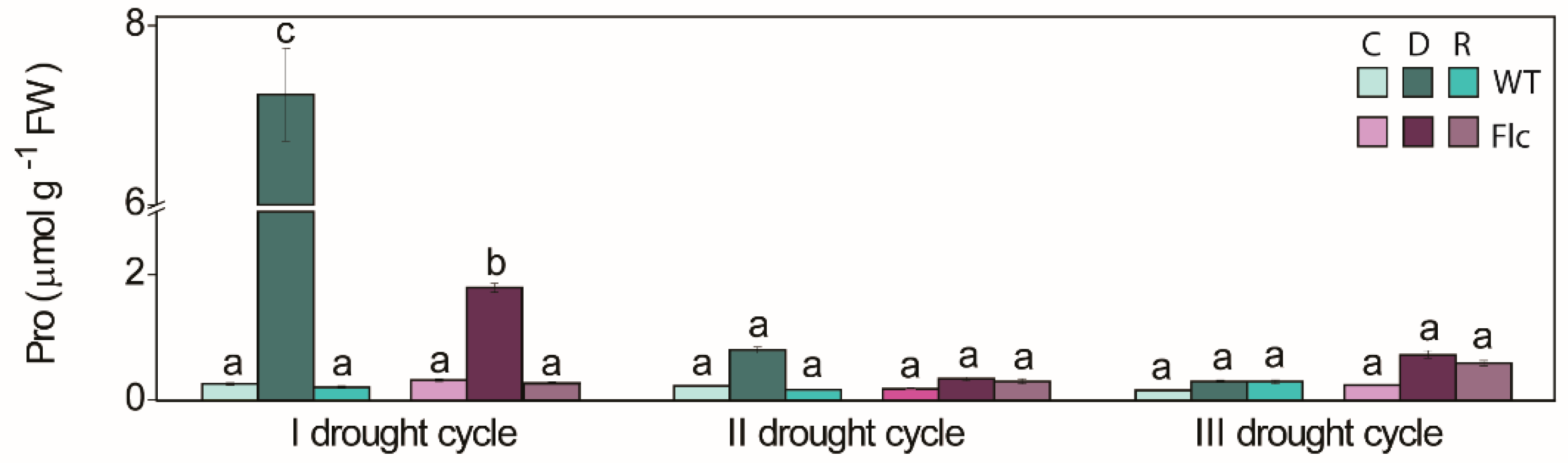
2.1.2. Drought-Induced Changes in Proline Content
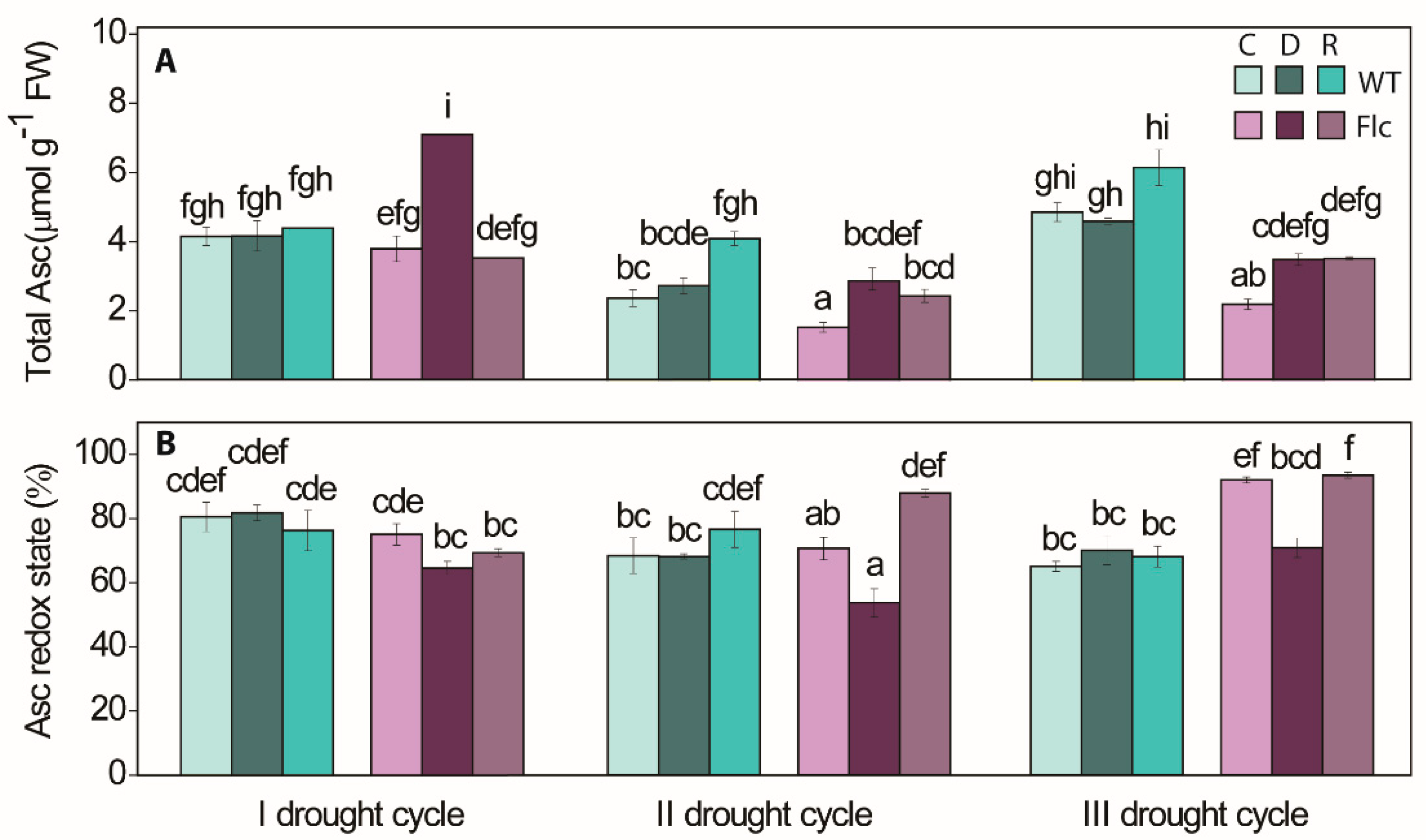
2.1.3. Drought-Induced Changes in Ascorbate Redox State
2.1.4. Drought-Induced Changes in ABA-Responsive Genes Expression
2.2. Prolonged Recovery after First Drought and Recovery after Third Drought
2.2.1. Effects of Prolonged Recovery after First Drought and Recovery after Third Drought on Growth Parameters in WT and flacca
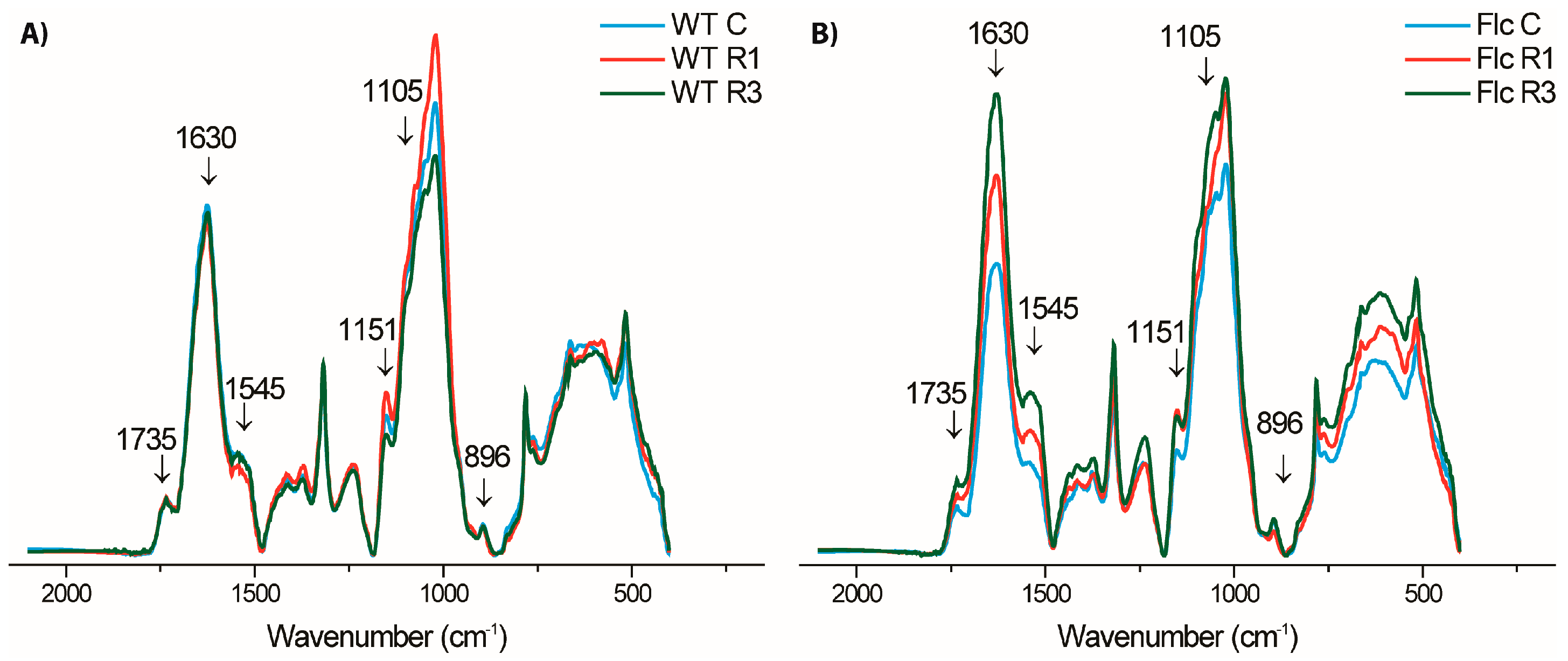
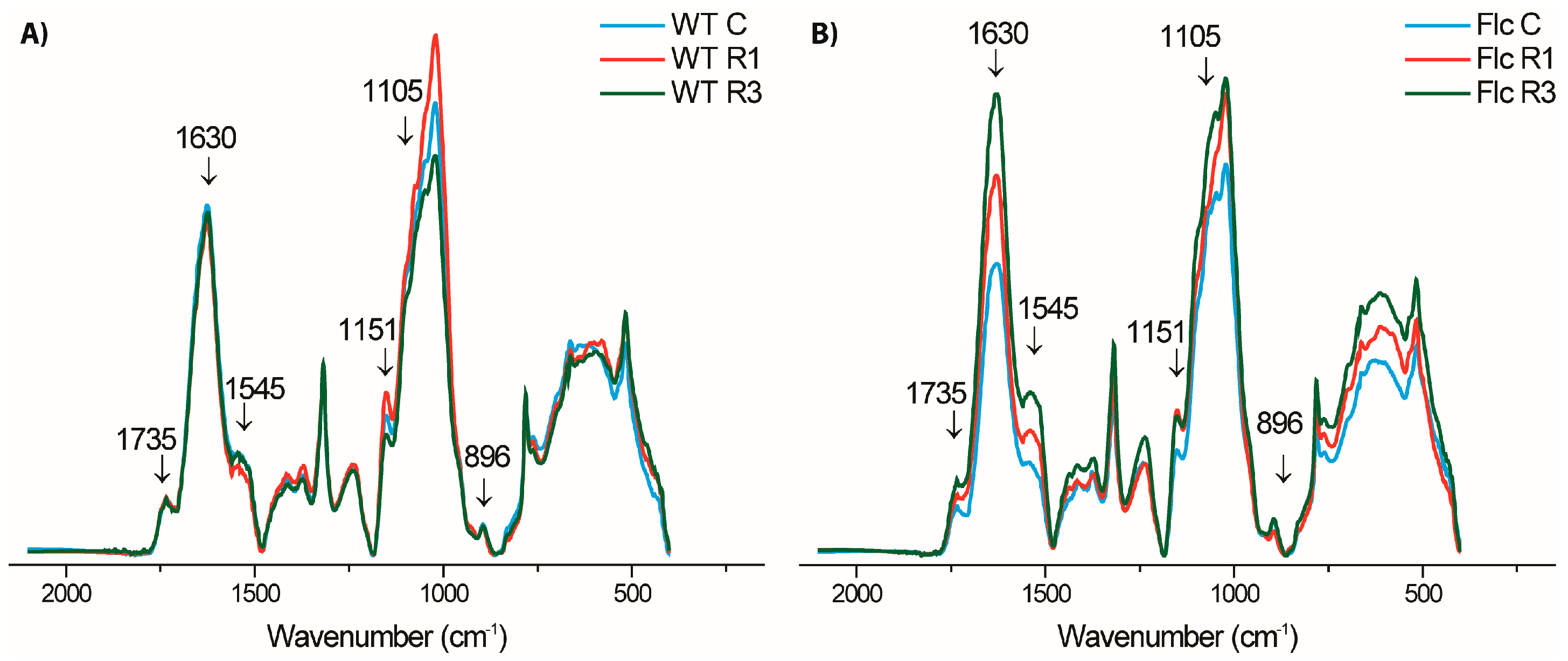
2.2.2. Effects of Prolonged Recovery after First Drought and Recovery after Third Drought on Cell Wall Composition in WT and flacca
3. Discussion
3.1. Low ABA Differences Result in flacca Traits
3.2. Stomatal Sensitivity Increases with Exposure to Recurrent Drought
3.3. Proline Accumulation Decreases with Repeating Drought Cycles
3.4. Drought-Induced Oxidative Stress
3.5. Drought Differentially Affected Growth and Cell Wall Compounds Accumulation in Two Genotypes
4. Materials and Methods
4.1. Plant Material and Experimental Setup
4.2. Measurements of Morphological and Physiological Parameters
4.3. Extraction and Analysis of Abscisic Acid Content
4.4. Determination of Leaf Proline Content
4.5. Determination of Total Leaf Ascorbate Content and Ascorbate Redox State
4.6. RNA Isolation and Real Time PCR
4.7. Cell Wall Isolation and Purification
4.8. FTIR Spectroscopy
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Naumann, G.; Cammalleri, C.; Mentaschi, L.; Feyen, L. Increased economic drought impacts in Europe with anthropo-genic warming. Nat. Clim. Chang. 2021, 11, 485–491. [Google Scholar] [CrossRef]
- Rojas, O. Agricultural extreme drought assessment at global level using the FAO-Agricultural Stress Index System (ASIS). Weather Clim. Extremes 2018, 27, 100184. [Google Scholar] [CrossRef]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. [Google Scholar]
- Pinheiro, C.; Chaves, M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Huseynova, I.M. Photosynthetic characteristics and enzymatic antioxidant capacity of leaves from wheat cultivars exposed to drought. Biochim. Biophys. Acta Bioenerg. 2012, 1817, 1516–1523. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Sanglard, L.M.V.P.; Avila, R.; Morais, L.E.; Martins, S.C.V.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.; Fernie, A.R.; et al. Photosynthetic and metabolic acclimation to repeated drought events play key roles in drought tolerance in coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.; Jentsch, A.; Beierkuhnlein, C.; Kreyling, J. Ecological stress memory and cross stress tolerance in plants in the face of climate extremes. Environ. Exp. Bot. 2013, 94, 3–8. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noctor, G.; Mhamdi, A.; Foyer, C. The Roles of Reactive Oxygen Metabolism in Drought: Not So Cut and Dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyngrope, S.; Bhoomika, K.; Dubey, R. Reactive oxygen species, ascorbate–glutathione pool, and enzymes of their me-tabolism in drought-sensitive and tolerant indica rice (Oryza sativa L.) seedlings subjected to progressing levels of water deficit. Protoplasma 2013, 250, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Gruszka, D.; Janeczko, A.; Dziurka, M.; Pociecha, E.; Fodor, J. Non-enzymatic antioxidant accumulations in BR-deficient and BR-insensitive barley mutants under control and drought conditions. Physiol. Plant. 2017, 163, 155–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesten, C.; Menna, A.; Sánchez-Rodríguez, C. Regulation of cellulose synthesis in response to stress. Curr. Opin. Plant Biol. 2017, 40, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.; Kim, S.H.; Jung, M.S.; Kim, M.S.; Oh, J.E.; Ju, H.W.; Kim, K.i.; Vierling, E.; Lee, H.; Hong, S.W. Arabidopsis hot2 encodes an endochitinase-like protein that is essential for tolerance to heat, salt and drought stresses. Plant J. 2007, 49, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q.; Dai, S. Drought-Responsive Mechanisms in Plant Leaves Revealed by Proteomics. Int. J. Mol. Sci. 2016, 17, 1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.-S.; Sun, L.; Dong, X.; Lu, S.-J.; Tian, W.; Liu, J.-X. Cellulose synthesis genes CESA6 and CSI1 are important for salt stress tolerance in Arabidopsis. J. Integr. Plant Biol. 2015, 58, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.B.; Methenni, K.; Nouairi, I.; Zarrouk, M.; Youssef, N.B. Drought priming improves subsequent more severe drought in a drought-sensitive cultivar of olive cv. Chétoui. Sci. Horticult. 2017, 221, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, T.J.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Fleta-Soriano, E.; Pinto-Marijuan, M.; Munné-Bosch, S. Evidence of Drought Stress Memory in the Facultative CAM, Apteniacordifolia: Possible Role of Phytohormones. PLoS ONE 2015, 10, e0135391. [Google Scholar] [CrossRef] [Green Version]
- Tombesi, S.; Frioni, T.; Poni, S.; Palliotti, A. Effect of water stress “memory” on plant behavior during subsequent drought stress. Environ. Exp. Bot. 2018, 150, 106–114. [Google Scholar] [CrossRef]
- Wang, X.; Mao, Z.; Zhang, J.; Hemat, M.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Osmolyte accumulation plays important roles in the drought priming induced tolerance to post-anthesis drought stress in winter wheat (Triticum aestivum L.). Environ. Exp. Bot. 2019, 166, 103804. [Google Scholar] [CrossRef]
- Lukić, N.; Kukavica, B.; Davidović-Plavšić, B.; Hasanagić, D.; Walter, J. Plant stress memory is linked to high levels of anti-oxidative enzymes over several weeks. Environ. Exp. Bot. 2020, 178, 104166. [Google Scholar] [CrossRef]
- Sabir, N.; Singh, B. Protected cultivation of vegetables in global arena: A review. Indian J. Agric. Sci. 2013, 83, 123–135. [Google Scholar]
- Food and Agriculture Organization. 2019. Available online: http://www.fao.org/faostat/en/#home (accessed on 26 December 2019).
- Živanović, B.; Vidović, M.; MilićKomić, S.; Jovanović, L.; Kolarž, P.; Morina, F.; Veljović Jovanović, S. Contents of phenolics and carotenoids in tomato grown under polytunnels with different UV-transmission rates. Turk. J. Agric. For. 2017, 41, 113–120. [Google Scholar] [CrossRef]
- Tamburino, R.; Vitale, M.; Ruggiero, A.; Sassi, M.; Sannino, L.; Arena, S.; Costa, A.; Batelli, G.; Zambrano, N.; Scaloni, A.; et al. Chloroplast proteome response to drought stress and recovery in tomato (Solanum lycopersicum L.). BMC Plant Biol. 2017, 17, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar Rai, G.; Parveen, A.; Jamwal, G.; Basu, U.; Ranjan Kumar, R.; Rai, P.K.; Sharma, J.P.; Alalawi, A.I.; Al-Duais, M.A.; Hossain, M.A.; et al. Leaf Proteome response to droughts and antioxidant potential in tomato (Solanum lycopersicum L.). Atmosphere 2021, 12, 1021. [Google Scholar]
- Sant’Ana, D.V.P.; Lefsrud, M. Tomato proteomics: Tomato as a model for crop proteomics. Sci. Horticult. 2018, 239, 224–233. [Google Scholar] [CrossRef]
- Iovieno, P.; Punzo, P.; Guida, G.; Mistretta, C.; van Oosten, M.J.; Nurcato, R.; Bostan, H.; Colatuono, C.; Costa, A.; Bagnaresi, P.; et al. Transcriptomic changes drive physiological responses to progressive drought stress and rehydration in tomato. Frontiers Plant Sci. 2016, 7, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuteja, N. Abscisic Acid and Abiotic Stress Signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic Acid Signaling and Abiotic Stress Tolerance in Plants: A Review on Current Knowledge and Future Prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, Q.; Xie, J.; Huang, M.; Cai, J.; Zhou, Q.; Dai, T.; Jiang, D. Abscisic acid and jasmonic acid are involved in drought priming-induced tolerance to drought in wheat. Crop. J. 2020, 9, 120–132. [Google Scholar] [CrossRef]
- Li, P.; Yang, H.; Wang, L.; Liu, H.; Huo, H.; Zhang, C.; Liu, A.; Zhu, A.; Hu, J.; Lin, Y.; et al. Physiological and Transcriptome Analyses Reveal Short-Term Responses and Formation of Memory Under Drought Stress in Rice. Front. Genet. 2019, 10, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrentino, G.; Haworth, M.; Wahbi, S.; Mahmood, T.; Zuomin, S.; Centritto, M. Abscisic Acid Induces Rapid Reductions in Mesophyll Conductance to Carbon Dioxide. PLoS ONE 2016, 11, e0148554. [Google Scholar] [CrossRef] [Green Version]
- Kollist, H.; Nuhkat, M.; Roelfsema, M.R.G. Closing gaps: Linking elements that control stomatal movement. New Phytol. 2014, 203, 44–62. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Fromm, M.E.; Avramova, Z. Multiple exposures to drought ‘train’ transcriptional responses in Arabidopsis. Nat. Commun. 2012, 3, 740. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.; Farrona, S. Plant Epigenetic Stress Memory Induced by Drought: A Physiological and Molecular Perspective. Plant Epigenetics Epigenomics 2020, 2093, 243–259. [Google Scholar] [CrossRef]
- Virlouvet, L.; Fromm, M. Physiological and transcriptional memory in guard cells during repetitive dehydration stress. New Phytol. 2014, 205, 596–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Zhu, J.-K. Regulation of Abscisic Acid Biosynthesis. Plant Physiol. 2003, 133, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartung, W.; Wilkinson, S.; Davies, W.J. Factors that regulate abscisic acid concentrations at the primary site of action at the guard cell. J. Exp. Bot. 1998, 49, 361–367. [Google Scholar] [CrossRef]
- Sagi, M.; Fluhr, R.; Lips, S.H. Aldehyde oxidase and xanthine dehydrogenase in a flacca tomato mutant with deficient abscisic acid and wilty phenotype. Plant Physiol. 1999, 120, 571–578. [Google Scholar]
- Sagi, M.; Scazzocchio, C.; Fluhr, R. The absence of molybdenum cofactor sulfuration is the primary cause of the flacca phenotype in tomato plants. Plant J. 2002, 31, 305–317. [Google Scholar] [CrossRef]
- Chen, G.; Shi, Q.; Lips, S.; Sagi, M. Comparison of growth of flacca and wild-type tomato grown under conditions diminishing their differences in stomatal control. Plant Sci. 2003, 164, 753–757. [Google Scholar] [CrossRef]
- Cornish, K.; Zeevaart, J.A.D.; Cohn, M.A.; Butera, D.L.; Hughes, J.A. Phenotypic Expression of Wild-Type Tomato and Three Wilty Mutants in Relation to Abscisic Acid Accumulation in Roots and Leaflets of Reciprocal Grafts. Plant Physiol. 1988, 87, 190–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imber, D.; Tal, M. Phenotypic Reversion of Flacca, a Wilty Mutant of Tomato, by Abscisic Acid. Science 1970, 169, 592–593. [Google Scholar] [CrossRef]
- Taylor, I.B.; Tarr, A.R. Phenotypic interactions between abscisic acid deficient tomato mutants. Theor. Appl. Genet. 1984, 68, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Živanović, B.; MilićKomić, S.; Tosti, T.; Vidović, M.; Prokić, L.; VeljovićJovanović, S. Leaf soluble sugars and free amino acids as important components of abscisic acid—Mediated drought response in tomato. Plants 2020, 9, 1147. [Google Scholar] [CrossRef]
- Lim, C.W.; Baek, W.; Jung, J.; Kim, J.-H.; Lee, S.C. Function of ABA in Stomatal Defense against Biotic and Drought Stresses. Int. J. Mol. Sci. 2015, 16, 15251–15270. [Google Scholar] [CrossRef] [Green Version]
- Dodd, I.C.; Theobald, J.C.; Richer, S.K.; Davies, W.J. Partial phenotypic reversion of ABA-deficient flacca tomato (Solanum lycopersicum) scions by a wild-type rootstock: Normalizing shoot ethylene relations promotes leaf area but does not diminish whole plant transpiration rate. J. Exp. Bot. 2009, 60, 4029–4039. [Google Scholar] [CrossRef]
- Fang, L.; Abdelhakim, L.; Hegelund, J.N.; Li, S.; Liu, J.; Peng, X.; Li, X.; Wei, Z.; Liu, F. ABA-mediated regulation of leaf and root hydraulic conductance in tomato grown at elevated CO2 is associated with altered gene expression of aquaporins. Hortic. Res. 2019, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntatsi, G.; Savvas, D.; Huntenburg, K.; Druege, U.; Hincha, D.K.; Zuther, E.; Schwarz, D. A study on ABA involvement in the response of tomato to suboptimal root temperature using reciprocal grafts with notabilis, a null mutant in the ABA-biosynthesis gene LeNCED1. Environ. Exp. Bot. 2014, 97, 11–21. [Google Scholar] [CrossRef]
- Poór, P.; Borbély, P.; Czékus, Z.; Takács, Z.; Ördög, A.; Popović, B.; Tari, I. Comparison of changes in water status and photosynthetic parameters in wild type and abscisic acid-deficient sitiens mutant of tomato (Solanum lycopersicum cv. RheinlandsRuhm) exposed to sublethal and lethal salt stress. J. Plant Physiol. 2018, 232, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.; Sharp, C.S.; Higgs, K.H. Growth and Water Relations of Wilty Mutants of Tomato (Lycopersicon esculentum Mill.). J. Exp. Bot. 1987, 38, 1848–1856. [Google Scholar] [CrossRef]
- Nagel, O.W.; Konings, H.; Lambers, H. Growth rate, plant development and water relations of the ABA-deficient tomato mutant sitiens. Physiol. Plant. 1994, 92, 102–108. [Google Scholar] [CrossRef]
- Sharp, R.E. Interaction with ethylene: Changing views on the role of abscisic acid in root and shoot growth responses to water stress. Plant Cell Environ. 2002, 25, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Giorio, P.; Guida, G.; Mistretta, C.; Sellami, M.H.; Oliva, M.; Punzo, P.; Iovieno, P.; Arena, C.; de Maio, A.; Grillo, S.; et al. Physiological, biochemical and molecular responses to water stress and rehydration in Mediterranean adapted tomato landraces. Plant Biol. 2018, 20, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Auler, P.A.; Amaral, M.N.D.; Rossatto, T.; Crizel, R.L.; Milech, C.; Chaves, F.C.; Souza, G.M.; Braga, E.J.B. Metabolism of abscisic acid in two contrasting rice genotypes submitted to recurrent water deficit. Physiol. Plant. 2020, 172, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Đurić, M.; Subotić, A.; Prokić, L.; Trifunović-Momčilov, M.; Cingel, A.; Vujičić, M.; Milošević, S. Morpho-physiological and molecular evaluation of drought and recovery in impatiens walleriana grown ex vitro. Plants 2020, 9, 1559. [Google Scholar] [CrossRef] [PubMed]
- Dörffling, K.; Streich, J.; Kruse, W.; Muxfeldt, B. Abscisic acid and the after-effect of water stress on stomatal opening potential. Z. Pflanzenphysiologie 1977, 81, 43–56. [Google Scholar] [CrossRef]
- Stålfelt, M.G. The Stomata as a Hydrophotic Regulator of the Water Deficit of the Plant. Physiol. Plant. 1955, 8, 572–593. [Google Scholar] [CrossRef]
- Milosavljevic, A.; Prokic, L.; Marjanovic, M.; Stikic, R.; Sabovljevic, A. The effects of drought on the expression of TAO1, NCED and EIL1 genes and ABA content in tomato wild-type and flacca mutant. Arch. Biol. Sci. 2012, 64, 297–306. [Google Scholar] [CrossRef]
- Muñoz-Espinoza, V.A.; López-Climent, M.F.; Casaretto, J.A.; Egomez-Cadenas, A. Water Stress Responses of Tomato Mutants Impaired in Hormone Biosynthesis Reveal Abscisic Acid, Jasmonic Acid and Salicylic Acid Interactions. Front. Plant Sci. 2015, 6, 997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAdam, E.L.; Brodribb, T.J.; McAdam, S.A. Does ozone increase ABA levels by non-enzymatic synthesis causing stomata to close? Plant Cell Environ. 2017, 40, 741–747. [Google Scholar] [CrossRef]
- Postiglione, A.E.; Muday, G.K. The Role of ROS Homeostasis in ABA-Induced Guard Cell Signaling. Front. Plant Sci. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed]
- Davies, W.J.; Kudoyarova, G.; Hartung, W. Long-distance ABA Signaling and Its Relation to Other Signaling Pathways in the Detection of Soil Drying and the Mediation of the Plant’s Response to Drought. J. Plant Growth Regul. 2005, 24, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Hartung, W.; Sauter, A.; Hose, E. Abscisic acid in the xylem: Where does it come from, where does it go to? J. Exp. Bot. 2002, 53, 27–32. [Google Scholar] [CrossRef]
- Yan, W.; Zheng, S.; Zhong, Y.; Shangguan, Z. Contrasting dynamics of leaf potential and gas exchange during progressive drought cycles and recovery in Amorph afruticosa and Robinia pseudoacacia. Sci. Rep. 2017, 7, 4470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, T.N. The control of stomata by water balance. New Phytol. 2005, 168, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Munemasa, S.; Hauser, F.; Park, J.; Waadt, R.; Brandt, B.; Schroeder, J.I. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr. Opin. Plant Biol. 2015, 28, 154–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.; John, G.P.; Pan, R.; Bartlett, M.K.; Fletcher, L.R.; Scoffoni, C.; Sack, L. A stomatal safety-efficiency trade-off constrains responses to leaf dehydration. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantin, F.; Monnet, F.; Jannaud, D.; Costa, J.M.; Renaud, J.; Muller, B.; Simonneau, T.; Genty, B. The dual effect of abscisic acid on stomata. New Phytol. 2012, 197, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Alguacil, M.D.M.; Vernieri, P.; Ruiz-Lozano, J.M. Plant Responses to Drought Stress and Exogenous ABA Application are Modulated Differently by Mycorrhization in Tomato and an ABA-deficient Mutant (Sitiens). Microb. Ecol. 2008, 56, 704–719. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Wang, Z. Abscisic Acid and Glycine Betaine Mediated Tolerance Mechanisms under Drought Stress and Recovery in Axonopuscompressus: A New Insight. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedipour, S. Relationship of Grain Yield, ABA and Proline Accumulation in Tolerant and Sensitive Wheat Cultivars as Affected by Water Stress. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2013, 83, 311–315. [Google Scholar] [CrossRef]
- Ábrahám, E.; Rigó, G.; Székely, G.; Nagy, R.; Koncz, C.; Szabados, L. Light-dependent induction of proline biosynthesis by abscisic acid and salt stress is inhibited by brassinosteroid in Arabidopsis. Plant Mol. Biol. 2003, 51, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Leufen, G.; Noga, G.; Hunsche, M. Drought stress memory in sugar beet: Mismatch between biochemical and physio-logical parameters. J. Plant Growth Regul. 2016, 35, 680–689. [Google Scholar] [CrossRef]
- Alves, R.D.F.B.; Menezes-Silva, P.E.; Sousa, L.F.; Loram-Lourenço, L.; Silva, M.L.F.; Almeida, S.E.S.; Silva, F.G.; de Souza, L.P.; Fernie, A.R.; Farnese, F.S. Evidence of drought memory in Dipteryxalata indicates differential acclimation of plants to savanna conditions. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Sharma, S.; Villamor, J.G.; Verslues, P.E. Essential Role of Tissue-Specific Proline Synthesis and Catabolism in Growth and Redox Balance at Low Water Potential. Plant Physiol. 2011, 157, 292–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verslues, P.E.; Bray, E.A. Role of abscisic acid (ABA) and Arabidopsis thaliana ABA-insensitive loci in low water potential-induced ABA and proline accumulation. J. Exp. Bot. 2005, 57, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Yoshiba, Y.; Kiyosue, T.; Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of levels of proline as an osmolyte in plants under water stress. Plant Cell Physiol. 1997, 38, 1095–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majer, P.; Vidović, M.; Czégény, G.; Jovanović, S.V.; Strid, Å.; Hideg, É. Evaluation of procedures for assessing anti-and pro-oxidants in plant samples. Anal. Methods 2016, 8, 5569–5580. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and Glutathione: The Heart of the Redox Hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hideg, É.; Jansen, M.A.; Strid, Å. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Petrović, I.; Jovanović, Z.; Stikić, R.; Marjanović, M.; Savić, S. Influence of severe drought on leaf response in ABA contrasting tomato genotypes (wild type and flacca Mutant). Biol. Life Sci. Forum 2021, 4, 96. [Google Scholar] [CrossRef]
- Li, X.; Ahammed, G.J.; Zhang, Y.Q.; Zhang, G.Q.; Sun, Z.H.; Zhou, J.; Zhou, Y.H.; Xia, X.J.; Yu, J.Q.; Shi, K. Carbon dioxide enrichment alleviates heat stress by improving cellular redox homeostasis through an ABA-independent process in tomato plants. Plant Biol. 2014, 17, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-W.; Wu, Z.-L.; Feng, L.-Y.; Dong, L.-H.; Song, A.-J.; Yuan, M.; Chen, Y.-E.; Zeng, J.; Chen, G.-D.; Yuan, S. Mg-Protoporphyrin IX Signals Enhance Plant’s Tolerance to Cold Stress. Front. Plant Sci. 2016, 7, 1545. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Shi, S.; Liu, Z.; Yang, F.; Yin, G. Drought tolerance in alfalfa (Medicago sativa L.) varieties is associated with enhanced antioxidative protection and declined lipid peroxidation. J. Plant Physiol. 2018, 232, 226–240. [Google Scholar] [CrossRef]
- Iyer, N.J.; Tang, Y.; Mahalingam, R. Physiological, biochemical and molecular responses to a combination of drought and ozone in Medicagotruncatula. Plant Cell Environ. 2012, 36, 706–720. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Drought Induces Oxidative Stress and Enhances the Activities of Antioxidant Enzymes in Growing Rice Seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Hasanagić, D.; Koleška, I.; Kojić, D.; Vlaisavljević, S.; Janjić, N.; Kukavica, B. Long term drought effects on tomato leaves: Anatomical, gas exchange and antioxidant modifications. Acta Physiol. Plant. 2020, 42, 1–14. [Google Scholar] [CrossRef]
- Chawla, S.; Jain, S.; Jain, V. Salinity induced oxidative stress and antioxidant system in salt-tolerant and salt-sensitive cultivars of rice (Oryza sativa L.). J. Plant Biochem. Biotechnol. 2012, 22, 27–34. [Google Scholar] [CrossRef]
- Gomes, F.P.; Oliva, M.A.; Mielke, M.S.; Almeida, A.-A.F.; Aquino, L.A. Osmotic adjustment, proline accumulation and cell membrane stability in leaves of Cocos nucifera submitted to drought stress. Sci. Hortic. 2010, 126, 379–384. [Google Scholar] [CrossRef]
- Erice, G.; Louahlia, S.; Irigoyen, J.J.; Sánchez-Díaz, M.; Avice, J.-C. Biomass partitioning, morphology and water status of four alfalfa genotypes submitted to progressive drought and subsequent recovery. J. Plant Physiol. 2010, 167, 114–120. [Google Scholar] [CrossRef]
- Sun, C.; Gao, X.; Chen, X.; Fu, J.; Zhang, Y. Metabolic and growth responses of maize to successive drought and rewatering cycles. Agric. Water Manag. 2016, 172, 62–73. [Google Scholar] [CrossRef]
- Chen, D.; Wang, S.; Xiong, B.; Cao, B.; Deng, X. Carbon/Nitrogen Imbalance Associated with Drought-Induced Leaf Senescence in Sorghum bicolor. PLoS ONE 2015, 10, e0137026. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Zhou, G.; Shimizu, H. Are plant growth and photosynthesis limited by pre-drought following rewatering in grass? J. Exp. Bot. 2009, 60, 3737–3749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellstein, C.; Poschlod, P.; Gohlke, A.; Chelli, S.; Campetella, G.; Rosbakh, S.; Canullo, R.; Kreyling, J.; Jentsch, A.; Beierkuhnlein, C. Effects of extreme drought on specific leaf area of grassland species: A meta-analysis of experimental studies in temperate and sub-Mediterranean systems. Glob. Chang. Biol. 2017, 23, 2473–2481. [Google Scholar] [CrossRef]
- Moore, J.P.; Vicré-Gibouin, M.; Farrant, J.; Driouich, A. Adaptations of higher plant cell walls to water loss: Drought vs. desiccation. Physiol. Plant. 2008, 134, 237–245. [Google Scholar] [CrossRef]
- Kačuráková, M.; Smith, A.C.; Gidley, M.J.; Wilson, R.H. Molecular interactions in bacterial cellulose composites studied by 1D FT-IR and dynamic 2D FT-IR spectroscopy. Carbohydr. Res. 2002, 337, 1145–1153. [Google Scholar] [CrossRef]
- Kalisz, G.; Gieroba, B.; Chrobak, O.; Suchora, M.; Starosta, A.; Sroka-Bartnicka, A. Vibrational Spectroscopic Analyses and Imaging of the Early Middle Ages Hemp BastFibres Recovered from Lake Sediments. Molecules 2021, 26, 1314. [Google Scholar] [CrossRef]
- Liang, C.Y.; Marchessault, R.H. Infrared spectra of crystalline polysaccharides: II Native celluloses in the region from 640 to 1700 cm−1. J. Polym. Sci. 1959, 39, 269–278. [Google Scholar] [CrossRef]
- Marchessault, R.H. Application of infra-red spectroscopy to cellulose and wood polysaccharides. Pure Appl. Chem. 1962, 5, 107–130. [Google Scholar] [CrossRef]
- Simonović, J.; Stevanic, J.; Djikanović, D.; Salmén, L.; Radotić, K. Anisotropy of cell wall polymers in branches of hardwood and softwood: A polarized FTIR study. Cellulose 2011, 18, 1433–1440. [Google Scholar] [CrossRef]
- Stewart, D. Fourier Transform Infrared Microspectroscopy of Plant Tissues. Appl. Spectrosc. 1996, 50, 357–365. [Google Scholar] [CrossRef]
- Szymanska-Chargot, M.; Zdunek, A. Use of FT-IR Spectra and PCA to the Bulk Characterization of Cell Wall Residues of Fruits and Vegetables along a Fraction Process. Food Biophys. 2012, 8, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef] [PubMed]
- Roig-Oliver, M.; Bresta, P.; Nadal, M.; Liakopoulos, G.; Nikolopoulos, D.; Karabourniotis, G.; Bota, J.; Flexas, J. Cell wall composition and thickness affect mesophyll conductance to CO2 diffusion in Helianthus annuus under water deprivation. J. Exp. Bot. 2020, 71, 7198–7209. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.M.; Peixoto-Junior, R.F.; Ribeiro, R.V.; Nobile, P.; Brito, M.S.; Marchiori, P.E.; Carlin, S.D.; Martins, A.P.B.; Goldman, M.H.S.; Llerena, J.P.P.; et al. Biomass Accumulation and Cell Wall Structure of Rice Plants Overexpressing a Dirigent-Jacalin of Sugarcane (ShDJ) Under Varying Conditions of Water Availability. Front. Plant Sci. 2019, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Wang, Y.; Xie, H.; Qiu, C.; Zhang, S.; Xiao, J.; Li, H.; Chen, L.; Li, X.; Ding, Z. Drought stress triggers proteomic changes involving lignin, flavonoids and fatty acids in tea plants. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Jiang, Y.; Yao, Y.; Wang, Y. Physiological Response, Cell Wall Components, and Gene Expression of Switchgrass under Short-Term Drought Stress and Recovery. Crop Sci. 2012, 52, 2718–2727. [Google Scholar] [CrossRef]
- Li, T.; Huang, Y.; Khadr, A.; Wang, Y.-H.; Xu, Z.-S.; Xiong, A.-S. DcDREB1A, a DREB-binding transcription factor from Daucus carota, enhances drought tolerance in transgenic Arabidopsis thaliana and modulates lignin levels by regulating lignin-biosynthesis-related genes. Environ. Exp. Bot. 2019, 169, 103896. [Google Scholar] [CrossRef]
- Leucci, M.R.; Lenucci, M.S.; Piro, G.; Dalessandro, G. Water stress and cell wall polysaccharides in the apical root zone of wheat cultivars varying in drought tolerance. J. Plant Physiol. 2008, 165, 1168–1180. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Moreno, M.J.; Gago, J.; Díaz-Vivancos, P.; Bernal, A.; Miedes, E.; Bresta, P.; Liakopoulos, G.; Fernie, A.R.; Hernández, J.A.; Flexas, J. The apoplastic antioxidant system and altered cell wall dynamics influence mesophyll conductance and the rate of photosynthesis. Plant J. 2019, 99, 1031–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roig-Oliver, M.; Nadal, M.; Clemente-Moreno, M.J.; Bota, J.; Flexas, J. Cell wall components regulate photosynthesis and leaf water relations of Vitis vinifera cv. Grenache acclimated to contrasting environmental conditions. J. Plant Physiol. 2019, 244, 153084. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-M.; Wang, F.; Wang, Y.-H.; Chen, T.; Hu, Y.-X.; Lin, J.-X. Anatomical and chemical characteristics of foliar vascular bundles in four reed ecotypes adapted to different habitats. Flora 2006, 201, 555–569. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Morina, F.; Jovanović, L.; Mojović, M.; Vidovic, M.; Pankovic, D.; Jovanovic, S.V. Zinc-induced oxidative stress in Verbascumthapsus L. is caused by an accumulation of reactive oxygen species and quinhydrone in the cell wall. Physiol. Plant. 2010, 140, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Vidović, M.; Morina, F.; Milić, S.; Albert, A.; Zechmann, B.; Tosti, T.; Winkler, J.B.; Jovanović, S.V. Carbon allocation from source to sink leaf tissue in relation to flavonoid biosynthesis in variegated Pelargonium zonale under UV-B radiation and high PAR intensity. Plant Physiol. Biochem. 2015, 93, 44–55. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Simonović-Radosavljević, J.; Pristov, J.B.; Mitrović, A.L.; Steinbach, G.; Mouille, G.; Tufegdžić, S.; Maksimović, V.; Mutavdžić, D.; Janošević, D.; Vuković, M. Parenchyma cell wall structure in twining stem of Dioscorea balcanica. Cellulose 2017, 24, 4653–4669. [Google Scholar]
- Bro, R.; Smilde, A.K. Principal component analysis. Anal. Methods 2014, 6, 2812–2831. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | |||
| C | R1 | R3 | |
| Leaves FW (g) | 31.86 ± 0.94 b | 40.75 ± 1.97 c | 19.89 ± 1.63 a |
| Stem FW (g) | 26.14 ± 0.59 ab | 23.84 ± 2.63 a | 33.47 ± 1.65 b |
| Plant FW (g) | 58.01 ± 1.53 a | 64.59 ± 4.60 a | 53.37 ± 3.3 a |
| Leaves DW (g) | 2.08 ± 0.02 b | 2.36 ± 0.05 c | 1.63 ± 0.07 a |
| Stem DW (g) | 2.10 ± 0.06 a | 2.17 ± 0.30 a | 1.70 ± 0.19 a |
| Plant DW (g) | 5.78 ± 0.11 ab | 6.09 ± 0.41 b | 4.50 ± 0.36 a |
| LA (cm2) | 1019. 9 ± 63.1 ab | 1137.9 ± 5.9 b | 880.8 ± 13.9 a |
| SLA (cm2 g−1DW) | 490.6 ± 26.8 a | 483.2 ± 7.7 a | 541.5 ± 16.1 a |
| flacca | |||
| C | R1 | R3 | |
| Leaves FW (g) | 27.26 ± 0.63 a | 38.49 ± 2.07 b | 27.08 ± 3.69 a |
| Stem FW (g) | 15.84 ± 0.60 b | 15.55 ± 0.71 ab | 13.01 ± 0.50 a |
| Plant FW (g) | 43.10 ± 0.1 ab | 54.1 ± 1.6 b | 40.1 ± 4.2 a |
| Leaves DW (g) | 1.13 ± 0.01 a | 2.23 ± 0.01 b | 1.97 ± 0.16 b |
| Stem DW (g) | 1.05 ± 0.02 a | 1.56 ± 0.01 b | 0.88 ± 0.09 a |
| Plant DW (g) | 2.91 ± 0.01 a | 5.28 ± 0.02 b | 4.16 ± 0.46 b |
| LA (cm2) | 630.1 ± 3.8 a | 963.9 ± 8.7 b | 544.1 ± 41.7 a |
| SLA (cm2 g−1DW) | 557.7 ± 3.8 c | 433.1 ± 3.2 b | 276.7 ± 1.1 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Živanović, B.; Milić Komić, S.; Nikolić, N.; Mutavdžić, D.; Srećković, T.; Veljović Jovanović, S.; Prokić, L. Differential Response of Two Tomato Genotypes, Wild Type cv. Ailsa Craig and Its ABA-Deficient Mutant flacca to Short-Termed Drought Cycles. Plants 2021, 10, 2308. https://doi.org/10.3390/plants10112308
Živanović B, Milić Komić S, Nikolić N, Mutavdžić D, Srećković T, Veljović Jovanović S, Prokić L. Differential Response of Two Tomato Genotypes, Wild Type cv. Ailsa Craig and Its ABA-Deficient Mutant flacca to Short-Termed Drought Cycles. Plants. 2021; 10(11):2308. https://doi.org/10.3390/plants10112308
Chicago/Turabian StyleŽivanović, Bojana, Sonja Milić Komić, Nenad Nikolić, Dragosav Mutavdžić, Tatjana Srećković, Sonja Veljović Jovanović, and Ljiljana Prokić. 2021. "Differential Response of Two Tomato Genotypes, Wild Type cv. Ailsa Craig and Its ABA-Deficient Mutant flacca to Short-Termed Drought Cycles" Plants 10, no. 11: 2308. https://doi.org/10.3390/plants10112308
APA StyleŽivanović, B., Milić Komić, S., Nikolić, N., Mutavdžić, D., Srećković, T., Veljović Jovanović, S., & Prokić, L. (2021). Differential Response of Two Tomato Genotypes, Wild Type cv. Ailsa Craig and Its ABA-Deficient Mutant flacca to Short-Termed Drought Cycles. Plants, 10(11), 2308. https://doi.org/10.3390/plants10112308