
Landscape Is the Main Driver of Weed Assemblages in Field Margins but Is Outperformed by Crop Competition in Field Cores


Abstract
:1. Introduction
2. Results
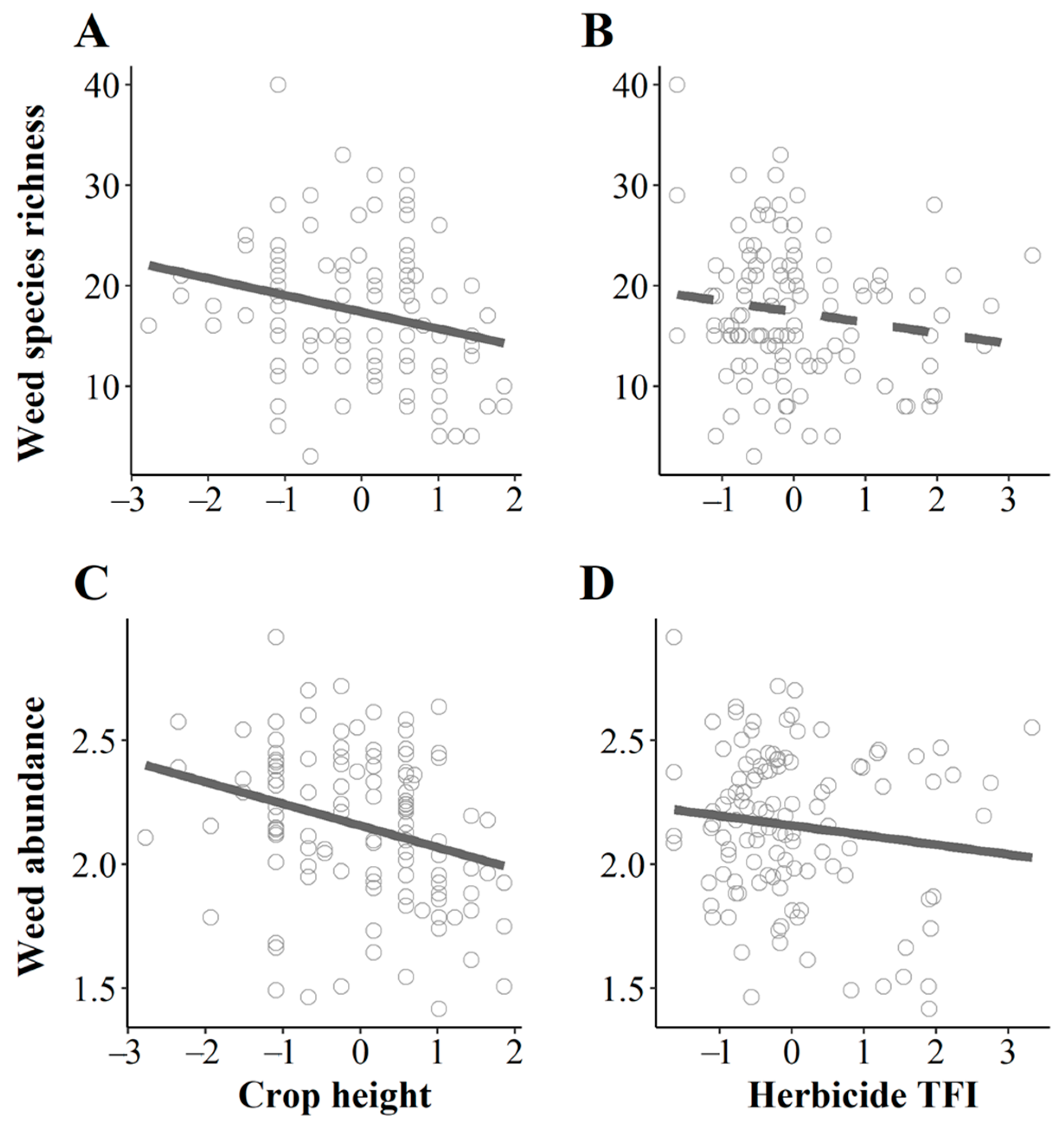
2.1. Competition and Weed Management Highly Affect Weed Species Richness in Field Core
2.2. Competition and Weed Management Strongly Affect Weed Abundance in Field Cores
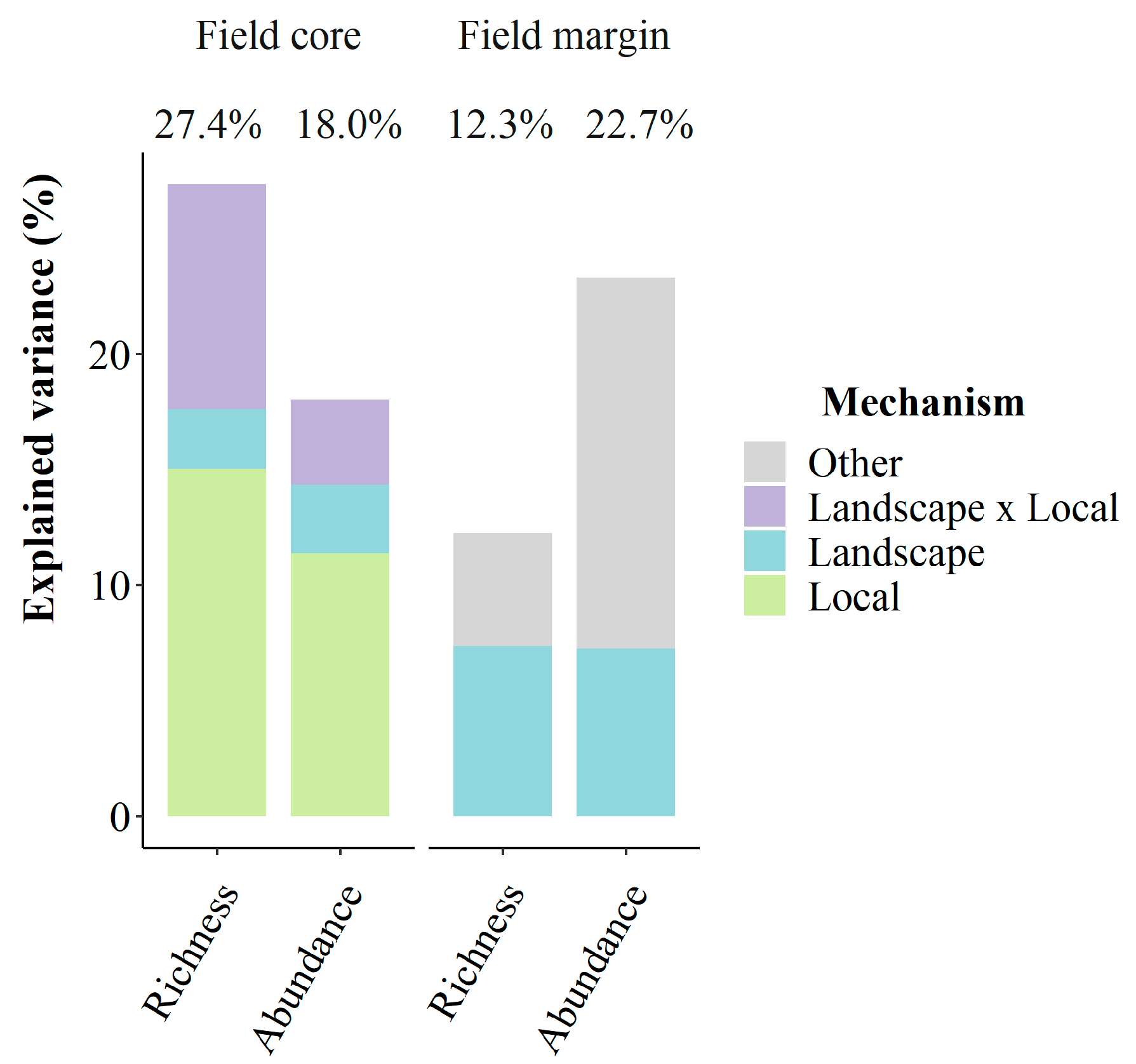
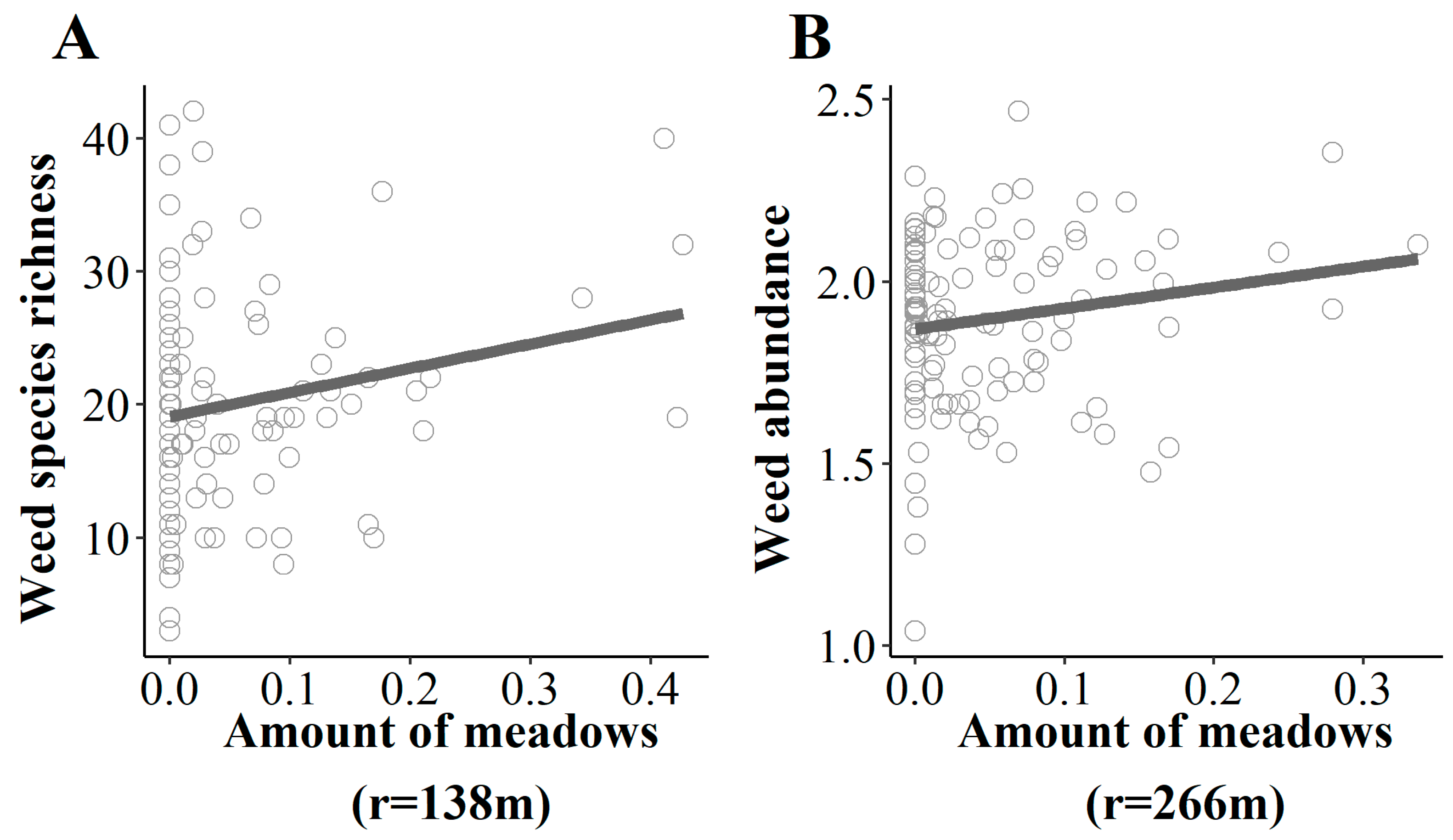
2.3. Landscape Is a Major Driver of Weed Assemblages in Field Margins
2.4. Multiscale Processes Shape Weed Assemblages in Field Cores
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Weed Sampling
4.3. Local, Landscape and Environmental Variables
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altieri, M.A.; Nicholls, C.I. Scaling up Agroecological Approaches for Food Sovereignty in Latin America. Development 2008, 51, 472–480. [Google Scholar] [CrossRef]
- Gliessman, S.R. 3 Sustainable agriculture: An agroecological perspective. In Advances in Plant Pathology; Andrews, J.H., Tommerup, I.C., Eds.; Academic Press: Cambridge, MA, USA, 1995; Volume 11, pp. 45–57. ISBN 0736-4539. [Google Scholar]
- Oerke, E.-C. Crop Losses to Pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Richner, N.; Holderegger, R.; Linder, H.P.; Walter, T. Reviewing change in the arable flora of Europe: A meta-analysis. Weed Res. 2015, 55, 1–13. [Google Scholar] [CrossRef]
- Albrecht, H.; Cambecèdes, J.; Lang, M.; Wagner, M. Management options for the conservation of rare arable plants in Europe. Bot. Lett. 2016, 163, 389–415. [Google Scholar] [CrossRef] [Green Version]
- Bretagnolle, V.; Gaba, S. Weeds for Bees? A Review. Agron. Sustain. Dev. 2015, 35, 891–909. [Google Scholar] [CrossRef] [Green Version]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Gaba, S.; Cheviron, N.; Perrot, T.; Piutti, S.; Gautier, J.-L.; Bretagnolle, V. Weeds enhance multifunctionality in arable lands in south-west of France. Front. Sustain. Food Syst. 2020, 4, 71. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef]
- MacLaren, C.; Storkey, J.; Menegat, A.; Metcalfe, H.; Dehnen-Schmutz, K. An ecological future for weed science to sustain crop production and the environment. A review. Agron. Sustain. Dev. 2020, 40, 24. [Google Scholar] [CrossRef]
- Alignier, A.; Solé-Senan, X.O.; Robleño, I.; Baraibar, B.; Fahrig, L.; Giralt, D.; Gross, N.; Martin, J.; Recasens, J.; Sirami, C.; et al. Configurational crop heterogeneity increases within-field plant diversity. J. Appl. Ecol. 2020, 57, 654–663. [Google Scholar] [CrossRef]
- Bourgeois, B.; Gaba, S.; Plumejeaud, C.; Bretagnolle, V. Weed diversity is driven by complex interplay between multi-scale dispersal and local filtering. Proc. R. Soc. B Biol. Sci. 2020, 287, 20201118. [Google Scholar] [CrossRef]
- Gaba, S.; Fried, G.; Kazakou, E.; Chauvel, B.; Navas, M.-L. Agroecological weed control using a functional approach: A review of cropping systems diversity. Agron. Sustain. Dev. 2014, 34, 103–119. [Google Scholar] [CrossRef] [Green Version]
- Mahaut, L.; Fried, G.; Gaba, S. Patch dynamics and temporal dispersal partly shape annual plant communities in ephemeral habitat patches. Oikos 2018, 127, 147–159. [Google Scholar] [CrossRef]
- Metcalfe, H.; Milne, A.E.; Coleman, K.; Murdoch, A.J.; Storkey, J. Modelling the effect of spatially variable soil properties on the distribution of weeds. Ecol. Model. 2019, 396, 1–11. [Google Scholar] [CrossRef]
- Fried, G.; Norton, L.R.; Reboud, X. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 2008, 128, 68–76. [Google Scholar] [CrossRef]
- Gunton, R.M.; Petit, S.; Gaba, S. Functional traits relating arable weed communities to crop characteristics: Traits relating weed communities to crops. J. Veg. Sci. 2011, 22, 541–550. [Google Scholar] [CrossRef]
- Gandía, M.L.; Casanova, C.; Sánchez, F.J.; Tenorio, J.L.; Santín-Montanyá, M.I. Arable weed patterns according to temperature and latitude gradient in central and southern Spain. Atmosphere 2020, 11, 853. [Google Scholar] [CrossRef]
- Borgy, B.; Gaba, S.; Petit, S.; Reboud, X. Non-random distribution of weed species abundance in arable fields: Distribution of abundances in weed communities. Weed Res. 2012, 52, 383–389. [Google Scholar] [CrossRef]
- Melander, B.; Munier-Jolain, N.; Charles, R.; Wirth, J.; Schwarz, J.; van der Weide, R.; Bonin, L.; Jensen, P.K.; Kudsk, P. European perspectives on the adoption of nonchemical weed management in reduced-tillage systems for arable crops. Weed Technol. 2013, 27, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Kaur, R.; Chauhan, B.S. Understanding crop-weed-fertilizer-water interactions and their implications for weed management in agricultural systems. Crop Prot. 2018, 103, 65–72. [Google Scholar] [CrossRef]
- Gaba, S.; Caneill, J.; Nicolardot, B.; Perronne, R.; Bretagnolle, V. Crop competition in winter wheat has a higher potential than farming practices to regulate weeds. Ecosphere 2018, 9, e02413. [Google Scholar] [CrossRef] [Green Version]
- Sardana, V.; Mahajan, G.; Jabran, K.; Chauhan, B.S. Role of competition in managing weeds: An introduction to the special issue. Crop Prot. 2017, 95, 1–7. [Google Scholar] [CrossRef]
- Petit, S.; Gaba, S.; Grison, A.-L.; Meiss, H.; Simmoneau, B.; Munier-Jolain, N.; Bretagnolle, V. Landscape scale management affects weed richness but not weed abundance in winter wheat fields. Agric. Ecosyst. Environ. 2016, 223, 41–47. [Google Scholar] [CrossRef]
- Bohan, D.A.; Haughton, A.J. Effects of local landscape richness on in-field weed metrics across the Great Britain scale. Agric. Ecosyst. Environ. 2012, 158, 208–215. [Google Scholar] [CrossRef]
- Roschewitz, I.; Gabriel, D.; Tscharntke, T.; Thies, C. The effects of landscape complexity on arable weed species diversity in organic and conventional farming: Landscape complexity and weed species diversity. J. Appl. Ecol. 2005, 42, 873–882. [Google Scholar] [CrossRef]
- Henckel, L.; Börger, L.; Meiss, H.; Gaba, S.; Bretagnolle, V. Organic fields sustain weed metacommunity dynamics in farmland landscapes. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150002. [Google Scholar] [CrossRef] [Green Version]
- Kovács-Hostyánszki, A.; Batáry, P.; Báldi, A.; Harnos, A. Interaction of local and landscape features in the conservation of hungarian arable weed diversity: Weed richness in Hungarian cereal fields. Appl. Veg. Sci. 2011, 14, 40–48. [Google Scholar] [CrossRef]
- Medeiros, H.R.; Thibes Hoshino, A.; Ribeiro, M.C.; de Oliveira Menezes Junior, A. Landscape complexity affects cover and species richness of weeds in brazilian agricultural environments. Basic Appl. Ecol. 2016, 17, 731–740. [Google Scholar] [CrossRef]
- Pallavicini, Y.; Bastida, F.; Hernández-Plaza, E.; Petit, S.; Izquierdo, J.; Gonzalez-Andujar, J.L. Local factors rather than the landscape context explain species richness and functional trait diversity and responses of plant assemblages of mediterranean cereal field margins. Plants 2020, 9, 778. [Google Scholar] [CrossRef]
- Carpentier, F.; Martin, O. Siland a R Package for Estimating the Spatial Influence of Landscape. Sci. Rep. 2021, 11, 7488. [Google Scholar] [CrossRef]
- Bouchet, A.-S.; Laperche, A.; Bissuel-Belaygue, C.; Snowdon, R.; Nesi, N.; Stahl, A. Nitrogen use efficiency in rapeseed. A review. Agron. Sustain. Dev. 2016, 36, 38. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reboud, X. Weed flora shifts and specialisation in winter oilseed rape in France. Weed Res. 2015, 55, 514–524. [Google Scholar] [CrossRef]
- Smith, R.G.; Mortensen, D.A.; Ryan, M.R. A new hypothesis for the functional role of diversity in mediating resource pools and weed–crop competition in agroecosystems. Weed Res. 2010, 50, 37–48. [Google Scholar] [CrossRef]
- Lemerle, D.; Verbeek, B.; Coombes, N. Losses in grain yield of winter crops from lolium rigidum competition depend on crop species, cultivar and season. Weed Res. 1995, 35, 503–509. [Google Scholar] [CrossRef]
- Holman, J.D.; Bussan, A.J.; Maxwell, B.D.; Miller, P.R.; Mickelson, J.A. Spring wheat, canola, and sunflower response to persian darnel (Lolium Persicum) interference. Weed Technol. 2004, 18, 509–520. [Google Scholar] [CrossRef]
- Hashem, A.; Borger, C.P.D.; Riethmuller, G. Weed Suppression by Crop Competition in Three Crop Species in Western Australia; New Zealand Plant Protection Society: Christchurch, New Zealand, 2010; pp. 26–30. [Google Scholar]
- Rathke, G.; Behrens, T.; Diepenbrock, W. Integrated nitrogen management strategies to improve seed yield, oil content and nitrogen efficiency of winter oilseed rape (Brassica napus L.): A review. Agric. Ecosyst. Environ. 2006, 117, 80–108. [Google Scholar] [CrossRef]
- Fried, G.; Cordeau, S.; Metay, A.; Kazakou, E. Relative importance of environmental factors and farming practices in shaping weed communities structure and composition in French vineyards. Agric. Ecosyst. Environ. 2019, 275, 1–13. [Google Scholar] [CrossRef]
- Gaba, S.; Gabriel, E.; Chadœuf, J.; Bonneu, F.; Bretagnolle, V. Herbicides do not ensure for higher wheat yield, but eliminate rare plant species. Sci. Rep. 2016, 6, 30112. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.A.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12, 693–715. [Google Scholar] [CrossRef]
- Sullivan, E.R.; Powell, I.; Ashton, P.A. Long-term hay meadow management maintains the target community despite local-scale species turnover. Folia Geobot. 2018, 53, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Bourgeois, B.; Munoz, F.; Gaba, S.; Denelle, P.; Fried, G.; Storkey, J.; Violle, C. Functional biogeography of weeds reveals how anthropogenic management blurs trait–climate relationships. J. Veg. Sci. 2021, 32, e12999. [Google Scholar] [CrossRef]
- Bourgeois, B.; Munoz, F.; Fried, G.; Mahaut, L.; Armengot, L.; Denelle, P.; Storkey, J.; Gaba, S.; Violle, C. What makes a weed a weed? A large-scale evaluation of arable weeds through a functional lens. Am. J. Bot. 2019, 106, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Bretagnolle, V.; Berthet, E.; Gross, N.; Gauffre, B.; Plumejeaud, C.; Houte, S.; Badenhausser, I.; Monceau, K.; Allier, F.; Monestiez, P.; et al. Towards sustainable and multifunctional agriculture in farmland landscapes: Lessons from the integrative approach of a French LTSER platform. Sci. Total Environ. 2018, 627, 822–834. [Google Scholar] [CrossRef]
- Bretagnolle, V.; Berthet, E.; Gross, N.; Gauffre, B.; Plumejeaud, C.; Houte, S.; Badenhausser, I.; Monceau, K.; Allier, F.; Monestiez, P.; et al. Description of long-term monitoring of farmland biodiversity in a LTSER. Data Brief 2018, 19, 1310–1313. [Google Scholar] [CrossRef]
- Violle, C.; Garnier, E.; Lecoeur, J.; Roumet, C.; Podeur, C.; Blanchard, A.; Navas, M.-L. Competition, traits and resource depletion in plant communities. Oecologia 2009, 160, 747–755. [Google Scholar] [CrossRef]
- Möhring, N.; Gaba, S.; Finger, R. Quantity based indicators fail to identify extreme pesticide risks. Sci. Total Environ. 2019, 646, 503–523. [Google Scholar] [CrossRef] [Green Version]
- Boutin, C.; Baril, A.; Martin, P. Plant diversity in crop fields and woody hedgerows of organic and conventional farms in contrasting landscapes. Agric. Ecosyst. Environ. 2008, 123, 185–193. [Google Scholar] [CrossRef]
- Erwin, J. Temperature and light effects on seed germination. Minn. Flower Grow. Bull. 1991, 40, 16–23. [Google Scholar]
- Vidotto, F.; De Palo, F.; Ferrero, A. Effect of short-duration high temperatures on weed seed germination: High temperatures affecting weed seeds. Ann. Appl. Biol. 2013, 454–465. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.43.17. 2020. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 15 August 2021).
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Third Version; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 15 August 2021).
- Hertzog, L.R. How robust are structural equation models to model misspecification? A simulation study. arXiv 2019, arXiv:1803.06186v3 [stat.AP]. [Google Scholar]
- Lefcheck, J.S. PiecewiseSEM: Piecewise structural equation modeling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimated Buffer Radius (m) | Estimate | df | t-Value | p-Value | ||
|---|---|---|---|---|---|---|
| (A) Weed species richness | Intercept | 16.314 | 1 | 11.204 | <0.001 | |
| Crop height | −1.407 | 1 | −2.400 | 0.018 | ||
| Nitrogen | 1.058 | 1 | 1.090 | 0.278 | ||
| Herbicides | −1.787 | 1 | −1.788 | 0.077 | ||
| Hedge density | 17 | 48.506 | 2 | 1.418 | 0.159 | |
| Number of meadows | 500 | −4.439 | 2 | −0.366 | 0.715 | |
| Amount of organic farming | 777 | −6.513 | 2 | −0.979 | 0.330 | |
| Amount of oilseed rape | 600 | 6.913 | 2 | 0.714 | 0.477 | |
| Nitrogen × Herbicides | 2.017 | 1 | 3.316 | 0.001 | ||
| Nitrogen × Number of meadows | −47.839 | 1 | −3.061 | 0.003 | ||
| Nitrogen × Amount of organic farming | 0.420 | 1 | 0.099 | 0.921 | ||
| Herbicides × Number of meadows | −5.072 | 1 | −0.281 | 0.779 | ||
| Herbicides × Amount of organic farming | 16.105 | 1 | 2.051 | 0.043 | ||
| (B) Weed Abundance | ||||||
| Intercept | 2.103 | 1 | 42.803 | <0.001 | ||
| Crop Height | −0.088 | 1 | −3.101 | 0.002 | ||
| Herbicides | −0.075 | 1 | −2.223 | 0.028 | ||
| Hedge density | 68 | 6.262 | 2 | 1.345 | 0.181 | |
| Number of meadows | 26 | −0.167 | 2 | −0.429 | 0.669 | |
| Amount of organic farming | 140 | 0.159 | 2 | 0.814 | 0.418 | |
| Amount of oilseed rape | 26 | 0.199 | 2 | 1.075 | 0.285 | |
| Herbicides × Number of meadows | 0.938 | 1 | 1.674 | 0.097 | ||
| Herbicides × Amount of organic farming | 0.353 | 1 | 1.394 | 0.166 |
| (A) | Estimated Buffer Radius (m) | Estimate | df | t-Value | p-Value | |
|---|---|---|---|---|---|---|
| Intercept | 20.262 | 1 | 10.818 | <0.001 | ||
| Rainfall | −1.805 | 1 | −2.467 | 0.015 | ||
| Hedge density | 37 | −86.517 | 2 | −1.083 | 0.281 | |
| Number of meadows | 138 | 18.731 | 2 | 2.184 | 0.031 | |
| Amount of organic farming | 20 | 8.255 | 2 | 1.642 | 0.103 | |
| Amount of oilseed rape | 980 | −11.572 | 2 | −0.711 | 0.478 | |
| (B) | ||||||
| Intercept | 1.892 | 1 | 60.673 | <0.001 | ||
| Rainfall | −0.087 | 1 | −4.060 | <0.001 | ||
| Temperature | 0.051 | 1 | 2.310 | 0.023 | ||
| Hedge density | 5 | −0.990 | 2 | −1.892 | 0.061 | |
| Number of meadows | 266 | 0.664 | 2 | 1.991 | 0.049 | |
| Amount of organic farming | 22 | 0.168 | 2 | 1.244 | 0.216 | |
| Amount of oilseed rape | 8 | −0.099 | 2 | −0.755 | 0.452 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berquer, A.; Martin, O.; Gaba, S. Landscape Is the Main Driver of Weed Assemblages in Field Margins but Is Outperformed by Crop Competition in Field Cores. Plants 2021, 10, 2131. https://doi.org/10.3390/plants10102131
Berquer A, Martin O, Gaba S. Landscape Is the Main Driver of Weed Assemblages in Field Margins but Is Outperformed by Crop Competition in Field Cores. Plants. 2021; 10(10):2131. https://doi.org/10.3390/plants10102131
Chicago/Turabian StyleBerquer, Adrien, Olivier Martin, and Sabrina Gaba. 2021. "Landscape Is the Main Driver of Weed Assemblages in Field Margins but Is Outperformed by Crop Competition in Field Cores" Plants 10, no. 10: 2131. https://doi.org/10.3390/plants10102131
APA StyleBerquer, A., Martin, O., & Gaba, S. (2021). Landscape Is the Main Driver of Weed Assemblages in Field Margins but Is Outperformed by Crop Competition in Field Cores. Plants, 10(10), 2131. https://doi.org/10.3390/plants10102131