Breeding Buckwheat for Increased Levels and Improved Quality of Protein


Abstract
1. Introduction
2. Protein Levels in Buckwheat Grain and Milling Fractions
3. Quality of Buckwheat Protein Compared to Soybean and Cereals
4. Buckwheat Protein Digestibility
5. Bioactivity of Buckwheat Peptides
6. Buckwheat Breeding for High Protein Levels: Prospects for the Future
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holasova, M.; Fiedlerova, V.; Smrcinova, H.; Orsak, M.; Lachman, J.; Vavreinova, S. Buckwheat—The source of antioxidant activity in functional foods. Food Res. Int. 2002, 35, 207–211. [Google Scholar] [CrossRef]
- Kreft, M. Buckwheat phenolic metabolites in health and disease. Nutr. Res. Rev. 2016, 29, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Luthar, Z. Buckwheat genetic resources in Central Europe. In Buckwheat Germplasm in the World; Zhou, M., Kreft, I., Suvorova, G., Tang, Y., Woo, S.H., Eds.; Academic Press, An Imprint of Elsevier: London, UK, 2018; pp. 127–143. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Yan, J.; Zhou, M.; Woo, S.-H.; Weng, W.; Cheng, J.; Zhang, K. Tartary Buckwheat: An Under-utilized Edible and Medicinal Herb for Food and Nutritional Security. Food Rev. Int. 2020, 1–15. [Google Scholar] [CrossRef]
- Dziedzic, K.; Kurek, S.; Mildner–Szkudlarz, S.; Kreft, I.; Walkowiak, J. Fatty acids profile, sterols, tocopherol and squalene content in Fagopyrum tataricum seed milling fractions. J. Cereal Sci. 2020, 96, 103118. [Google Scholar] [CrossRef]
- Skrabanja, V.; Kreft, I. Resistant Starch Formation Following Autoclaving of Buckwheat (Fagopyrum esculentum Moench) Groats. An In Vitro Study. J. Agric. Food Chem. 1998, 46, 2020–2023. [Google Scholar] [CrossRef]
- Eggum, B.O. The protein quality of buckwheat in comparison with other protein sources of plant or animal origin. In Buckwheat: Genetics, Plant Breeding, Utilization; Kreft, I., Javornik, B., Dolinšek, B., Eds.; Biotechnical Faculty: Ljubljana, Slovenia, 1980; pp. 115–120. [Google Scholar]
- Skrabanja, V.; Kreft, I.; Golob, T.; Modic, M.; Ikeda, S.; Ikeda, K.; Kreft, S.; Bonafaccia, G.; Knapp, M.; Kosmelj, K. Nutrient Content in Buckwheat Milling Fractions. Cereal Chem. J. 2004, 81, 172–176. [Google Scholar] [CrossRef]
- Bonafaccia, G.; Marocchini, M.; Kreft, I. Composition and technological properties of the flour and bran from common and tartary buckwheat. Food Chem. 2003, 80, 9–15. [Google Scholar] [CrossRef]
- Vombergar, B.; Luthar, Z. The concentration of flavonoids, tannins and crude proteins in grain fractions of common buckwheat (Fagopyrum esculentum Moench) and Tartary buckwheat (Fagopyrum tataricum Gaertn.). Folia Boil. Geol. 2019, 59, 101–157. [Google Scholar] [CrossRef]
- Kump, P.; Rupnik, P.; Budnar, M.; Kreft, I. Analysis of sulphur in plant material. Nucl. Instrum. Methods Phys. Res. B 1977, 142, 205–208. [Google Scholar] [CrossRef]
- Vogel-Mikuš, K.; Pelicon, P.; Vavpetič, P.; Kreft, I.; Regvar, M. Elemental analysis of edible grains by micro-PIXE: Common buckwheat case study. Nucl. Instrum. Methods Phys. Res. B 2009, 267, 2884–2889. [Google Scholar] [CrossRef]
- Kaulich, B.; Gianoncelli, A.; Beran, A.; Eichert, D.; Kreft, I.; Pongrac, P.; Regvar, M.; Vogel-Mikuš, K.; Kiskinova, M. Low-energy X-ray fluorescence microscopy opening new opportunities for bio-related research. J. R. Soc. Interface 2009, 6, S641–S647. [Google Scholar] [CrossRef]
- Pongrac, P.; Vogel-Mikuš, K.; Regvar, M.; Vavpetič, P.; Pelicon, P.; Kreft, I. Improved Lateral Discrimination in Screening the Elemental Composition of Buckwheat Grain by Micro-PIXE. J. Agric. Food Chem. 2011, 59, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Pongrac, P.; Kelemen, M.; Vavpetič, P.; Vogel-Mikuš, K.; Regvar, M.; Pelicon, P. Application of micro-PIXE (particle induced X-ray emission) to study buckwheat grain structure and composition. Fagopyrum 2020, 37, 5–10. [Google Scholar] [CrossRef]
- Chrungoo, N.; Dohtdong, L.; Chettry, U. Diversity in Seed Storage Proteins and Their Genes in Buckwheat. In Molecular Breeding and Nutritional Aspects of Buckwheat; Academic Press is an Imprint of Elsevier: London, UK, 2016; pp. 387–399. [Google Scholar]
- Škrabanja, V.; Kreft, I. Nutritional Value of Buckwheat Proteins and Starch. In Molecular Breeding and Nutritional Aspects of Buckwheat; Academic Press is an Imprint of Elsevier: London, UK, 2016; pp. 169–176. [Google Scholar]
- Radović, S.R.; Maksimović, V.R.; Varkonyi-Gasic, E. Characterization of Buckwheat Seed Storage Proteins. J. Agric. Food Chem. 1996, 44, 972–974. [Google Scholar] [CrossRef]
- Joshi, D.C.; Zhang, K.; Wang, C.; Chandora, R.; Khurshid, M.; Li, J.; He, M.; Georgiev, M.I.; Zhou, M. Strategic enhancement of genetic gain for nutraceutical development in buckwheat: A genomics-driven perspective. Biotechnol. Adv. 2020, 39, 107479. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Khatkar, B.S. Pseudocereals nutritional composition functional properties and food applications. In Food Bioactives: Functionality and Applications in Human Health; Deka, S.C., Seth, D., Hulle, N.R.S., Eds.; Apple Academic Press: Cambridge, MA, USA, 2019; pp. 129–148. [Google Scholar]
- Sytar, O.; Brestic, M.; Zivcak, M.; Tran, L.-S.P. The Contribution of Buckwheat Genetic Resources to Health and Dietary Diversity. Curr. Genom. 2016, 17, 193–206. [Google Scholar] [CrossRef]
- Sytar, O.; Chrenková, M.; Ferencová, J.; Polačiková, M.; Rajský, M.; Brestič, M. Nutrient capacity of amino acids from buckwheat seeds and sprouts. J. Food Nutr. Res. 2018, 57, 38–47. [Google Scholar]
- Sytar, O.; Biel, W.; Smetanska, I.; Brestic, M. Bioactive Compounds and Their Biofunctional Properties of Different Buckwheat Germplasms for Food Processing. In Buckwheat Germplasm in the World; Zhou, M.L., Kreft, I., Suvorova, G., Tang, Y., Woo, S.H., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 191–204. [Google Scholar]
- Krumina-Zemture, G.; Beitane, I.; Gramatin, I. Amino acid and dietary fibre content of pea and buckwheat flours. Res. Rural Dev. 2016, 1, 84–90. [Google Scholar]
- Dziedzic, K.; Górecka, D.; Szwengiel, A.; Olejnik, A.; Rychlik, J.; Kreft, I.; Drożdżyńska, A.; Walkowiak, J. The cytotoxic effect of artificially digested buckwheat products on HT-29 colon cancer cells. J. Cereal Sci. 2018, 83, 68–73. [Google Scholar] [CrossRef]
- Fotschki, B.; Juśkiewicz, J.; Jurgoński, A.; Amarowicz, R.; Opyd, P.; Bez, J.; Muranyi, I.; Petersen, I.L.; Laparra, J.M. Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats. Nutrients 2020, 12, 2781. [Google Scholar] [CrossRef]
- Wang, C.; Yuan, S.; Zhang, W.; Ng, T.B.; Ye, X. Buckwheat Antifungal Protein with Biocontrol Potential To Inhibit Fungal (Botrytis cinerea) Infection of Cherry Tomato. J. Agric. Food Chem. 2019, 67, 6748–6756. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, Y.; Li, J.; Wang, Y.; Yang, X. Enzyme-assisted development of biofunctional polyphenol-enriched buckwheat protein: Physicochemical properties, in vitro digestibility, and antioxidant activity. J. Sci. Food Agric. 2019, 99, 3176–3185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, R.; Li, Y.M.; Liang, N.; Zhao, Y.; Zhu, H.; He, Z.; Liu, J.; Hao, W.; Jiao, R.; et al. Cholesterol-Lowering Activity of Tartary Buckwheat Protein. J. Agric. Food Chem. 2017, 65, 1900–1906. [Google Scholar] [CrossRef]
- Kayashita, J.; Shimaoka, I.; Yamazaki, M.; Kato, N. Buckwheat protein extract ameliorates atropine-induced constipation in rats. Curr. Adv. Buckwheat Res. 1995, 5, 941–946. [Google Scholar]
- Kayashita, J.; Shimaoka, I.; Nakajoh, M.; Kato, N. Feeding of buckwheat protein extract reduces hepatic triglyceride concentration, adipose tissue weight, and hepatic lipogenesis in rats. J. Nutr. Biochem. 1996, 7, 555–559. [Google Scholar] [CrossRef]
- Kayashita, J.; Shimaoka, I.; Nakajoh, M.; Kishida, N.; Kato, N. Consumption of a Buckwheat Protein Extract Retards 7,12-Dimethylbenz[α] anthracene-Induced Mammary Carcinogenesis in Rats. Biosci. Biotechnol. Biochem. 1999, 63, 1837–1839. [Google Scholar] [CrossRef]
- Kayashita, J.; Shimaoka, I.; Nakajoh, M.; Kondoh, M.; Hayashi, K.; Kato, N. Muscle Hypertrophy in Rats Fed on a Buckwheat Protein Extract. Biosci. Biotechnol. Biochem. 1999, 63, 1242–1245. [Google Scholar] [CrossRef]
- Stibilj, V.; Kreft, I.; Smrkolj, P.; Osvald, J. Enhanced selenium content in buckwheat (Fagopyrum esculentum Moench) and pumpkin (Cucurbita pepo L.) seeds by foliar fertilisation. Eur. Food Res. Technol. 2004, 219, 142–144. [Google Scholar] [CrossRef]
- Breznik, B.; Germ, M.; Gaberscik, A.; Kreft, I. Combined effects of elevated UV-B radiation and the addition of selenium on common (Fagopyrum esculentum Moench) and tartary [Fagopyrum tataricum (L.) Gaertn.] buckwheat. Photosyntetica 2005, 43, 583–589. [Google Scholar] [CrossRef]
- Tadina, N.; Germ, M.; Kreft, I.; Breznik, B.; Gaberscik, A. Effects of water deficit and selenium on common buckwheat (Fagopyrum esculentum Moench.) plants. Photosyntetica 2007, 45, 472–476. [Google Scholar] [CrossRef]
- Smrkolj, P.; Germ, M.; Kreft, I.; Stibilj, V. Respiratory potential and Se compounds in pea (Pisum sativum L.) plants grown from Se-enriched seeds. J. Exp. Bot. 2006, 57, 3595–3600. [Google Scholar] [CrossRef] [PubMed]
- Golob, A.; Gadžo, D.; Stibilj, V.; Djikić, M.; Gavrić, T.; Kreft, I.; Germ, M. Sulphur interferes with selenium accumulation in Tartary buckwheat plants. Plant Physiol. Biochem. 2016, 108, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Golob, A.; Stibilj, V.; Kreft, I.; Vogel-Mikuš, K.; Gaberscik, A.; Germ, M. Selenium treatment alters the effects of UV radiation on chemical and production parameters in hybrid buckwheat. Acta Agric. Scand. Sect. B-Plant Soil Sci. 2017, 68, 1–11. [Google Scholar] [CrossRef]
- Fabjan, N.; Rode, J.; Košir, I.J.; Wang, Z.; Zhang, Z.; Kreft, I. Tartary Buckwheat (Fagopyrum tataricum Gaertn.) as a Source of Dietary Rutin and Quercitrin. J. Agric. Food Chem. 2003, 51, 6452–6455. [Google Scholar] [CrossRef]
- Kreft, S.; Štrukelj, B.; Gaberščik, A.; Kreft, I. Rutin in buckwheat herbs grown at different UV-B radiation levels: Comparison of two UV spectrophotometric and an HPLC method. J. Exp. Bot. 2002, 53, 1801–1804. [Google Scholar] [CrossRef]
- Ikeda, K.; Kishida, M. Digestibility of proteins in buckwheat seed. Fagopyrum 1993, 13, 21–24. [Google Scholar]
- Kreft, I.; Javornik, B. Chemical composition and protein quality of buckwheat (Fagopyrum esculentum Moench). Plant Foods Hum. Nutr. 1981, 30, 175–179. [Google Scholar] [CrossRef]
- Skrabanja, V.; Laerke, H.; Kreft, I. Effects of hydrothermal processing of buckwheat (Fagopyrum esculentum Moench) groats on starch enzymatic availability in vitro and in vivo in rats. J. Cereal Sci. 1998, 28, 209–214. [Google Scholar] [CrossRef]
- Skrabanja, V.; Lærke, H.N.; Kreft, I. Protein-polyphenol interactions and in vivo digestibility of buckwheat groat proteins. Pflügers Archiv. 2000, 440, R129–R131. [Google Scholar] [CrossRef]
- Kalinová, J.P.; Vrchotová, N.; Tříska, J. Phenolics levels in different parts of common buckwheat (Fagopyrum esculentum) achenes. J. Cereal Sci. 2019, 85, 243–248. [Google Scholar] [CrossRef]
- Ikeda, K.; Oku, M.; Kusano, T.; Yasumoto, K. Inhibitory Potency of Plant Antinutrients towards the In Vitro Digestibility of Buckwheat Protein. J. Food Sci. 1986, 51, 1527–1530. [Google Scholar] [CrossRef]
- Oparin, P.B.; Mineev, K.S.; Dunaevsky, Y.E.; Arseniev, A.S.; Belozersky, M.A.; Grishin, E.V.; Egorov, T.A.; Vassilevski, A. Buckwheat trypsin inhibitor with helical hairpin structure belongs to a new family of plant defence peptides. Biochem. J. 2012, 446, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Annor, G.A.; Tyl, C.; Marcone, M.; Ragaee, S.; Marti, A. Why do millets have slower starch and protein digestibility than other cereals? Trends Food Sci. Technol. 2017, 66, 73–83. [Google Scholar] [CrossRef]
- Wieslander, G.; Fabjan, N.; Vogrincic, M.; Kreft, I.; Janson, C.; Spetz-Nyström, U.; Vombergar, B.; Tagesson, C.; Leanderson, P.; Norbäck, D. Eating Buckwheat Cookies Is Associated with the Reduction in Serum Levels of Myeloperoxidase and Cholesterol: A Double Blind Crossover Study in Day-Care Centre Staffs. Tohoku J. Exp. Med. 2011, 225, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Wieslander, G.; Fabjan, N.; Vogrinčič, M.; Kreft, I.; Vombergar, B.; Norbäck, D. Effects of common and Tartary buckwheat consumption on mucosal symptoms, headache and tiredness: A double-blind crossover intervention study. J. Food Agric. Environ. 2012, 10, 107–110. [Google Scholar]
- Tomotake, H.; Shimaoka, I.; Kayashita, J.; Yokoyama, F.; Nakajoh, M.; Kato, N. A Buckwheat Protein Product Suppresses Gallstone Formation and Plasma Cholesterol More Strongly than Soy Protein Isolate in Hamsters. J. Nutr. 2000, 130, 1670–1674. [Google Scholar] [CrossRef]
- Ma, Y.Y.; Xiong, Y.L. Antioxidant and bile acid binding activity of buckwheat protein in vitro digests. J. Agric. Food Chem. 2009, 57, 4372–4380. [Google Scholar] [CrossRef]
- Guo, X.-N.; Zhu, K.; Zhang, H.; Yao, H. Purification and Characterization of the Antitumor Protein from Chinese Tartary Buckwheat (Fagopyrum tataricum Gaertn.) Water-Soluble Extracts. J. Agric. Food Chem. 2007, 55, 6958–6961. [Google Scholar] [CrossRef]
- Guo, X.; Zhu, K.; Zhang, H.; Yao, H. Anti-Tumor Activity of a Novel Protein Obtained from Tartary Buckwheat. Int. J. Mol. Sci. 2010, 11, 5201–5211. [Google Scholar] [CrossRef]
- Leung, E.H.W.; Ng, T.B. A relatively stable antifungal peptide from buckwheat seeds with antiproliferative activity toward cancer cells. J. Pept. Sci. 2007, 13, 762–767. [Google Scholar] [CrossRef]
- Chen, X.-W.; Luo, D.-Y.; Chen, Y.-J.; Wang, J.-M.; Guo, J.; Yang, X. Dry fractionation of surface abrasion for polyphenol-enriched buckwheat protein combined with hydrothermal treatment. Food Chem. 2019, 285, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-M.; Ma, C.-Y. Extraction, purification and characterization of globulin from common buckwheat (Fagopyrum esculentum Moench) seeds. Food Res. Int. 2006, 39, 974–981. [Google Scholar] [CrossRef]
- Deng, Y.; Padilla-Zakour, O.; Zhao, Y.; Tao, S. Influences of High Hydrostatic Pressure, Microwave Heating, and Boiling on Chemical Compositions, Antinutritional Factors, Fatty Acids, In Vitro Protein Digestibility, and Microstructure of Buckwheat. Food Bioprocess Technol. 2015, 8, 2235–2245. [Google Scholar] [CrossRef]
- Fujimura, M.; Minami, Y.; Watanabe, K.; Tadera, K. Purification, Characterization, and Sequencing of a Novel Type of Antimicrobial Peptides, Fa-AMP1 andFa-AMP2, from Seeds of Buckwheat (Fagopyrum esculentum Moench.). Biosci. Biotechnol. Biochem. 2003, 67, 1636–1642. [Google Scholar] [CrossRef]
- Ahmed, A.; Khalid, N.; Ahmad, A.; Abbasi, N.A.; Latif, M.S.Z.; Randhawa, M.A. Phytochemicals and biofunctional properties of buckwheat: A review. J. Agric. Sci. 2013, 152, 349–369. [Google Scholar] [CrossRef]
- Nic Phiarais, B.P.; Schehl, B.D.; Arendt, E.K. Protein Changes during Malting of Buckwheat. J. Am. Soc. Brew. Chem. 2008, 66, 127–135. [Google Scholar] [CrossRef]
- Zhou, X. Toward a novel understanding of buckwheat self-defensive strategies during seed germination and preliminary investigation on the potential pharmacological application of its malting products. J. Med. Plants Res. 2011, 5, 6946–6954. [Google Scholar] [CrossRef]
- Wang, X.; Ullah, N.; Sun, X.; Guo, Y.; Chen, L.; Li, Z.; Feng, X. Development and characterization of bacterial cellulose reinforced biocomposite films based on protein from buckwheat distiller’s dried grains. Int. J. Biol. Macromol. 2017, 96, 353–360. [Google Scholar] [CrossRef]
- Pongrac, P.; Kump, P.; Budič, B.; Vogel-Mikuš, K. Magnesium and phosphorus distributions in developing tartary buckwheat cotyledons/Razporeditev magnezija in fosforja v razvijajočih se kličnih listih tatarske ajde. Folia Boil. Geol. 2017, 57, 45. [Google Scholar] [CrossRef]
- Pongrac, P.; Scheers, N.; Sandberg, A.-S.; Potisek, M.; Arčon, I.; Kreft, I.; Kump, P.; Vogel-Mikuš, K. The effects of hydrothermal processing and germination on Fe speciation and Fe bioaccessibility to human intestinal Caco-2 cells in Tartary buckwheat. Food Chem. 2016, 199, 782–790. [Google Scholar] [CrossRef]
- Pongrac, P.; Potisek, M.; Fraś, A.; Likar, M.; Budič, B.; Myszka, K.; Boros, D.; Nečemer, M.; Kelemen, M.; Vavpetič, P.; et al. Composition of mineral elements and bioactive compounds in tartary buckwheat and wheat sprouts as affected by natural mineral-rich water. J. Cereal Sci. 2016, 69, 9–16. [Google Scholar] [CrossRef]
- Ma, Q.; Zhao, Y.; Wang, H.-L.; Li, J.; Yang, Q.-H.; Gao, L.-C.; Murat, T.; Feng, B. Comparative study on the effects of buckwheat by roasting: Antioxidant properties, nutrients, pasting, and thermal properties. J. Cereal Sci. 2020, 95, 103041. [Google Scholar] [CrossRef]
- Koyama, M.; Hattori, S.; Amano, Y.; Watanabe, M.; Nakamura, K. Blood Pressure-Lowering Peptides from Neo-Fermented Buckwheat Sprouts: A New Approach to Estimating ACE-Inhibitory Activity. PLoS ONE 2014, 9, e105802. [Google Scholar] [CrossRef]
- Koyama, M.; Naramoto, K.; Nakajima, T.; Aoyama, T.; Watanabe, M.; Nakamura, K. Purification and Identification of Antihypertensive Peptides from Fermented Buckwheat Sprouts. J. Agric. Food Chem. 2013, 61, 3013–3021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Huang, L.; Tang, W.; Zhou, Y.; Wang, Q.; Li, Z. A novel buckwheat protein with a beneficial effect in atherosclerosis was purified from Fagopyrum tataricum (L.) Gaertn. Arch. Biol. Sci. 2013, 65, 767–772. [Google Scholar] [CrossRef]
- Zhou, X.; Wen, L.; Li, Z.; Zhou, Y.; Chen, Y.; Lu, Y. Advance on the benefits of bioactive peptides from buckwheat. Phytochem. Rev. 2015, 14, 381–388. [Google Scholar] [CrossRef]
- Tomotake, H.; Yamamoto, N.; Yanaka, N.; Ohinata, H.; Yamazaki, R.; Kayashita, J.; Kato, N. High protein buckwheat flour suppresses hypercholesterolemia in rats and gallstone formation in mice by hypercholesterolemic diet and body fat in rats because of its low protein digestibility. Nutrition 2006, 22, 166–173. [Google Scholar] [CrossRef]
- Tomotake, H.; Yamamoto, N.; Kitabayashi, H.; Kawakami, A.; Kayashita, J.; Ohinata, H.; Karasawa, H.; Kato, N. Preparation of Tartary Buckwheat Protein Product and Its Improving Effect on Cholesterol Metabolism in Rats and Mice Fed Cholesterol-Enriched Diet. J. Food Sci. 2007, 72, S528–S533. [Google Scholar] [CrossRef]
- Kayashita, J.; Nagai, H.; Kato, N. Buckwheat Protein Extract Suppression of the Growth Depression in Rats Induced by Feeding Amaranth (Food Red No. 2). Biosci. Biotechnol. Biochem. 1996, 60, 1530–1531. [Google Scholar] [CrossRef][Green Version]
- Kayashita, J.; Shimaoka, I.; Nakajoh, M.; Yamazaki, M.; Kato, N. Consumption of buckwheat protein lowers plasma cholesterol and raises fecal neutral sterols in cholesterol-Fed rats because of its low digestibility. J. Nutr. 1997, 127, 1395–1400. [Google Scholar] [CrossRef]
- Li, C.-H.; Matsui, T.; Matsumoto, K.; Yamasaki, R.; Kawasaki, T. Latent production of angiotensin I-converting enzyme inhibitors from buckwheat protein. J. Pept. Sci. 2002, 8, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Janssen, F.; Pauly, A.; Rombouts, I.; Jansens, K.J.; Deleu, L.J.; Delcour, J.A. Proteins of Amaranth (Amaranthus spp.), Buckwheat (Fagopyrum spp.), and Quinoa (Chenopodium spp.): A Food Science and Technology Perspective. Compr. Rev. Food Sci. Food Saf. 2017, 16, 39–58. [Google Scholar] [CrossRef]
- Jin, J.; Okagu, O.D.; Yagoub, A.E.A.; Udenigwe, C.C. Effects of sonication on the in vitro digestibility and structural properties of buckwheat protein isolates. Ultrason. Sonochem. 2021, 70, 105348. [Google Scholar] [CrossRef] [PubMed]
- Veličković, T.Ć.; Stanic-Vucinic, D.J. The Role of Dietary Phenolic Compounds in Protein Digestion and Processing Technologies to Improve Their Antinutritive Properties. Compr. Rev. Food Sci. Food Saf. 2017, 17, 82–103. [Google Scholar] [CrossRef]
- Xue, F.; Wu, Z.; Tong, J.; Zheng, J.; Li, C. Effect of combination of high-intensity ultrasound treatment and dextran glycosylation on structural and interfacial properties of buckwheat protein isolates. Biosci. Biotechnol. Biochem. 2017, 81, 1891–1898. [Google Scholar] [CrossRef]
- Yang, Z.-H.; Li, C.; Li, Y.-Y.; Wang, Z. Effects of Maillard reaction on allergenicity of buckwheat allergen Fag t 3 during thermal processing. J. Sci. Food Agric. 2012, 93, 1510–1515. [Google Scholar] [CrossRef]
- Lee, C.; In, S.; Han, Y.; Oh, S. Reactivity change of IgE to buckwheat protein treated with high-pressure and enzymatic hydrolysis. J. Sci. Food Agric. 2015, 96, 2073–2079. [Google Scholar] [CrossRef]
- Ličen, M.; Kreft, I. Buckwheat (Fagopyrum esculentum Moench) low molecular weight seed proteins are restricted to the embryo and are not detectable in the endosperm. Plant Physiol. Biochem. 2005, 43, 862–865. [Google Scholar] [CrossRef]
- Wieslander, G.; Norbäck, D. Buckwheat allergy. Allergy 2001, 56, 703–704. [Google Scholar] [CrossRef]
- Wang, T.-C.; Shyur, S.-D.; Wen, D.-C.; Kao, Y.-H.; Huang, L.-H. Buckwheat anaphylaxis: An unusual allergen in Taiwan. Asian Pac. J. Allergy Immunol. 2006, 24, 167–170. [Google Scholar]
- Jungewelter, S.; Airaksinen, L.; Pesonen, M. Occupational buckwheat allergy as a cause of allergic rhinitis, asthma, contact urticaria and anaphylaxis—An emerging problem in food-handling occupations? Am. J. Ind. Med. 2020, 63, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- De Arcangelis, E.; Cuomo, F.; Trivisonno, M.C.; Marconi, E.; Messia, M.C. Gelatinization and pasta making conditions for buckwheat gluten-free pasta. J. Cereal Sci. 2020, 95, 103073. [Google Scholar] [CrossRef]
- Clemente, A. Enzymatic protein hydrolysates in human nutrition. Trends Food Sci. Technol. 2000, 11, 254–262. [Google Scholar] [CrossRef]
- Kreft, I.; Fabjan, N.; Yasumoto, K. Rutin content in buckwheat (Fagopyrum esculentum Moench) food materials and products. Food Chem. 2006, 98, 508–512. [Google Scholar] [CrossRef]
- Tang, C.-H.; Wang, X.-Y.; Liu, F.; Wang, C.-S. Physicochemical and Conformational Properties of Buckwheat Protein Isolates: Influence of Polyphenol Removal with Cold Organic Solvents from Buckwheat Seed Flours. J. Agric. Food Chem. 2009, 57, 10740–10748. [Google Scholar] [CrossRef]
- Zhu, L.W.; Zhou, Y.; Cai, F.; Deng, J.; Huang, J.; Zhang, X.N.; Zhang, J.G.; Chen, Q.F. Quantitative analysis of perennial buckwheat leaves proteinand GABA using near infrared spectroscopy. Spectrosc. Spectr. Anal. 2020, 40, 2421–2426. [Google Scholar] [CrossRef]
- Nosworthy, M.G.; Franczyk, A.; Zimoch-Korzycka, A.; Appah, P.; Utioh, A.; Neufeld, J.; House, J.D. Impact of Processing on the Protein Quality of Pinto Bean (Phaseolus vulgaris) and Buckwheat (Fagopyrum esculentum Moench) Flours and Blends, As Determined by in Vitro and in Vivo Methodologies. J. Agric. Food Chem. 2017, 65, 3919–3925. [Google Scholar] [CrossRef]
- Tang, C.-H.; Peng, J.; Zhen, D.-W.; Chen, Z. Physicochemical and antioxidant properties of buckwheat (Fagopyrum esculentum Moench) protein hydrolysates. Food Chem. 2009, 115, 672–678. [Google Scholar] [CrossRef]
- Riedl, K.M.; Hagerman, A.E. Tannin−Protein Complexes as Radical Scavengers and Radical Sinks. J. Agric. Food Chem. 2001, 49, 4917–4923. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Zielinski, H. Buckwheat as a Functional Food and Its Effects on Health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef]
- Woo, S.H.; Roy, S.K.; Kwon, S.J.; Cho, S.W.; Sarker, K.; Lee, M.S.; Chung, K.Y.; Kim, H.H. Concepts, Prospects, and Potentiality in Buckwheat (Fagopyrum eculentum Moench): A research perspective. In Molecular Breeding and Nutritional Aspects of Buckwheat; Zhou, M., Kreft, I., Woo, S.H., Chrungoo, N., Wieslander, G., Eds.; Academic Press, an Imprint of Elsevier: London, UK, 2016; pp. 21–49. [Google Scholar]
- Yabe, S.; Iwata, H. Genomics-assisted breeding in minor and pseudo-cereals. Breed. Sci. 2020, 70, 19–31. [Google Scholar] [CrossRef]
- Vombergar, B.; Škrabanja, V.; Luthar, Z.; Germ, M. Starting points for the study of the effects of flavonoids, tannins and crude proteins in grain fractions of common buckwheat (Fagopyrum esculentum Moench) and Tartary buckwheat (Fagopyrum tataricum Gaertn.). Folia Biol. Geol 2017, 58, 101–145. [Google Scholar] [CrossRef]
- Jin, J.; Ohanenye, I.C.; Udenigwe, C.C. Buckwheat proteins: Functionality, safety, bioactivity, and prospects as alternative plant-based proteins in the food industry. Crit. Rev. Food Sci. Nut. 2020, 1–13. [Google Scholar] [CrossRef]
- Yanagihara, Y. Buckwheat hypersensitivity. Kansensho 1980, 10, 184–188. [Google Scholar]
- Yano, M.; Nakamura, R.; Hayakawa, S.; Torii, S. Purification and Properties of Allergenic Proteins in Buckwheat Seeds. Agric. Biol. Chem. 1989, 53, 2387–2392. [Google Scholar] [CrossRef]
- Urisu, A.; Kondo, Y.; Morita, Y.; Yagi, E.; Tsuruta, M.; Yasaki, T.; Yamada, K.; Kuzuya, H.; Suzuki, M.; Titani, K. Isolation and characterization of major allergen in buckwheat seed. In Current Advances in Buckwheat Research; Shinshu University Press: Matsumoto, Japan, 1995; pp. 965–974. [Google Scholar]
- Wang, Z.; Zhang, Z.; Zhao, Z.; Wieslander, G.; Norbäck, D.; Kreft, I. Purification and Characterization of a 24 kDa Protein from Tartary Buckwheat Seeds. Biosci. Biotechnol. Biochem. 2004, 68, 1409–1413. [Google Scholar] [CrossRef]
- Matsumoto, R.; Fujino, K.; Nagata, Y.; Hashiguchi, S.; Ito, Y.; Aihara, Y.; Takahashi, Y.; Maeda, K.; Sugimura, K. Molecular characterization of a 10-kDa buckwheat molecule reactive to allergic patients’ IgE. Allergy 2004, 59, 533–538. [Google Scholar] [CrossRef]
- Alonso-Miravalles, L.; Zannini, E.; Bez, J.; Arendt, E.K.; O’Mahony, J.A. Physical and flow properties of pseudocereal-based protein-rich ingredient powders. J. Food Eng. 2020, 281, 109973. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, Q.; Xia, Q.; Zha, B.; Sun, J.; Xu, B.; Shi, Y.-C. Intact endosperm cells in buckwheat flour limit starch gelatinization and digestibility in vitro. Food Chem. 2020, 330, 127318. [Google Scholar] [CrossRef]
- Zhou, X.L.; Yan, B.B.; Xiao, Y.; Zhou, Y.M.; Liu, T.Y. Tartary buckwheat protein prevented dyslipidemia in high-fat diet-fed mice associated with gut microbiota changes. Food Chem. Toxicol. 2018, 119, 296–301. [Google Scholar] [CrossRef]
- Ge, R.H.; Wang, H. Nutrient components and bioactive compounds in tartary buckwheat bran and flour as affected by thermal processing. Int. J. Food Prop. 2020, 23, 127–137. [Google Scholar] [CrossRef]
- Ji, X.; Han, L.; Liu, F.; Yin, S.; Peng, Q.; Wang, M. A mini-review of isolation, chemical properties and bioactivities of polysaccharides from buckwheat (Fagopyrum Mill). Int. J. Biol. Macromol. 2019, 127, 204–209. [Google Scholar] [CrossRef]
- Guo, X.; Xiong, Y.L. Characteristics and functional properties of buckwheat protein–sugar Schiff base complexes. LWT 2013, 51, 397–404. [Google Scholar] [CrossRef]
- Kreft, I.; Rutar, V.; Luthar, Z.; Javornik, B. Ugotavljanje velikosti kalčka in vsebnost olja z NMR pri žlahtnjenju visokolizinskega ječmena. Zb. BF UEK Ljubl. Slov. Agric. Issue 1986, 47, 25–31. [Google Scholar]
- Rolletschek, H.; Fuchs, J.; Friedel, S.; Börner, A.; Todt, H.; Jakob, P.M.; Borisjuk, L. A novel noninvasive procedure for high-throughput screening of major seed traits. Plant Biotechnol. J. 2014, 13, 188–199. [Google Scholar] [CrossRef]
- Ličen, M.; Dvořáček, V.; Čepková, P.; Michalová, A. Endosperm protein polymorphism of common and tartary buckwheat. Acta Agric. Slov. 2004, 83, 171–179. [Google Scholar]
- Luthar, Z.; Rogl, S.; Kump, B.; Javornik, B. 38–48 kDa subunits of buckwheat 13S globulins are controlled by a single locus. Plant Breed. 2008, 127, 322–324. [Google Scholar] [CrossRef]
- Kreft, I. Ajda; Kmečki Glas: Ljubljana, Slovenia, 1995; 112p. [Google Scholar]
- Adhikari, K.N.; Campbell, C. Natural outcrossing in common buckwheat. Euphytica 1998, 102, 233–237. [Google Scholar] [CrossRef]
- Luthar, Z. Genska banka ajde: Buckwheat genebank—A source of Slovenian genetic variability. Acta Agric. Slov. 2012, 99, 307–316. [Google Scholar] [CrossRef]
- Fang, X.; Zhang, Y.; Zhang, Y.; Huang, K.; Yang, W.; Li, X.; Zhang, Z.; Wu, K.; Xu, X.; Ruan, R.; et al. De novo transcriptome assembly and identification of genes related to seed size in common buckwheat (Fagopyrum esculentum M.). Breed. Sci. 2019, 69, 487–497. [Google Scholar] [CrossRef]
- Chen, P.; Guo, Y.F.; Yan, Q.; Li, Y.H. Molecular cloning and characterization of Fag t 2: A 16-kDa major allergen from Tartary buckwheat seeds. Allergy 2011, 66, 1393–1395. [Google Scholar] [CrossRef]
- Yasui, Y.; Hirakawa, H.; Ueno, M.; Matsui, K.; Katsube-Tanaka, T.; Yang, S.J.; Aii, J.; Sato, S.; Mori, M. Assembly of the draft genome of buckwheat and its applications in identifying agronomically useful genes. DNA Res. 2016, 23, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Yabe, S.; Hara, T.; Ueno, M.; Enoki, H.; Kimura, T.; Nishimura, S.; Yasui, Y.; Ohsawa, R.; Iwata, H. Potential of Genomic Selection in Mass Selection Breeding of an Allogamous Crop: An Empirical Study to Increase Yield of Common Buckwheat. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, R. Current status and prospects of common buckwheat breeding in Japan. Breed. Sci. 2020, 70, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Tomatsu, T.; Kinouchi, S.; Suzuki, T.; Sato, T. Identification of a gene encoding glutathione S-transferase that is related to anthocyanin accumulation in buckwheat (Fagopyrum esculentum). J. Plant Physiol. 2018, 231, 291–296. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, J.; Liu, X.; Geng, F.; Huang, Q.; Zhao, J.; Xiang, D.; Zhao, G. Analysis of tartary buckwheat (Fagopyrum tataricum) seed proteome using offline two-dimensional liquid chromatography and tandem mass spectrometry. J. Food Biochem. 2019, 43, e12863. [Google Scholar] [CrossRef]
- Yasui, Y. History of the progressive development of genetic marker systems for common buckwheat. Breed. Sci. 2020, 70, 13–18. [Google Scholar] [CrossRef]
- Suzuki, T.; Noda, T.; Morishita, T.; Ishiguro, K.; Otsuka, S.; Brunori, A. Present status and future perspectives of breeding for buckwheat quality. Breed. Sci. 2020, 70, 48–66. [Google Scholar] [CrossRef]
- Zhang, K.; He, M.; Fan, Y.; Zhao, H.; Gao, B.; Yang, K.; Li, F.; Tang, Y.; Gao, Q.; Lin, T.; et al. Resequencing of global Tartary buckwheat accessions reveals multiple domestication events and key loci associated with agronomic traits. Genome Biol. 2020. [Google Scholar] [CrossRef]



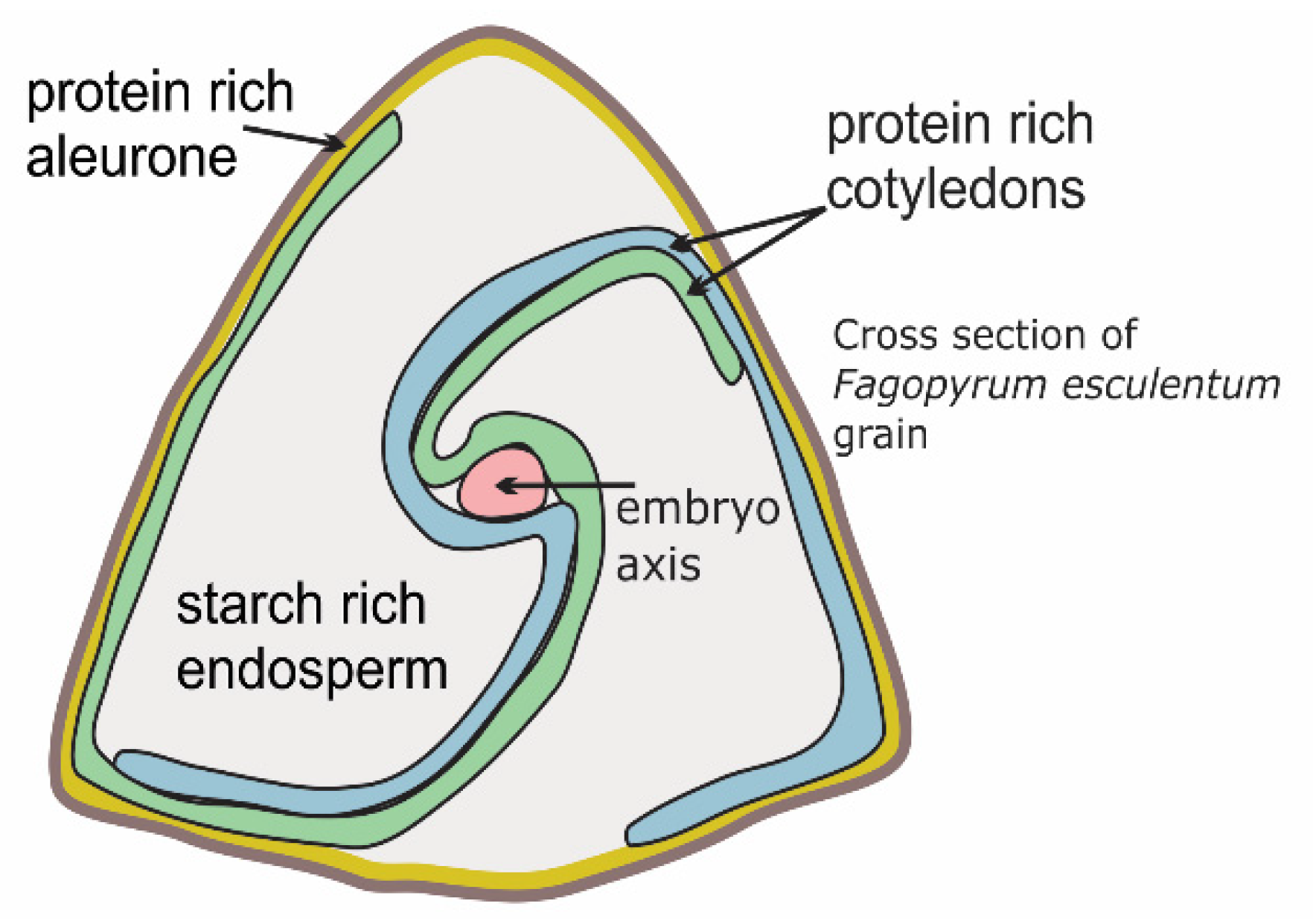{kind=link}
{kind=link}
| Protein Source | Amino Acid (g/16 g N) | Biological | ||
|---|---|---|---|---|
| Lysine | Methionine | Cysteine | Value (%) | |
| Buckwheat | 5.09 | 1.89 | 2.02 | 93.1 |
| Wheat | 2.55 | 1.81 | 1.79 | 62.5 |
| Soybean | 5.99 | 1.61 | 1.59 | 68.4 |
| Fava bean | 6.04 | 0.94 | 1.44 | 51.1 |
| Egg | 6.79 | 3.09 | 2.38 | 99.3 |
| Pork | 7.99 | 2.85 | 1.14 | 84.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luthar, Z.; Zhou, M.; Golob, A.; Germ, M. Breeding Buckwheat for Increased Levels and Improved Quality of Protein. Plants 2021, 10, 14. https://doi.org/10.3390/plants10010014
Luthar Z, Zhou M, Golob A, Germ M. Breeding Buckwheat for Increased Levels and Improved Quality of Protein. Plants. 2021; 10(1):14. https://doi.org/10.3390/plants10010014
Chicago/Turabian StyleLuthar, Zlata, Meiliang Zhou, Aleksandra Golob, and Mateja Germ. 2021. "Breeding Buckwheat for Increased Levels and Improved Quality of Protein" Plants 10, no. 1: 14. https://doi.org/10.3390/plants10010014
APA StyleLuthar, Z., Zhou, M., Golob, A., & Germ, M. (2021). Breeding Buckwheat for Increased Levels and Improved Quality of Protein. Plants, 10(1), 14. https://doi.org/10.3390/plants10010014