Next Chapter in the Legend of Silphion: Preliminary Morphological, Chemical, Biological and Pharmacological Evaluations, Initial Conservation Studies, and Reassessment of the Regional Extinction Event


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
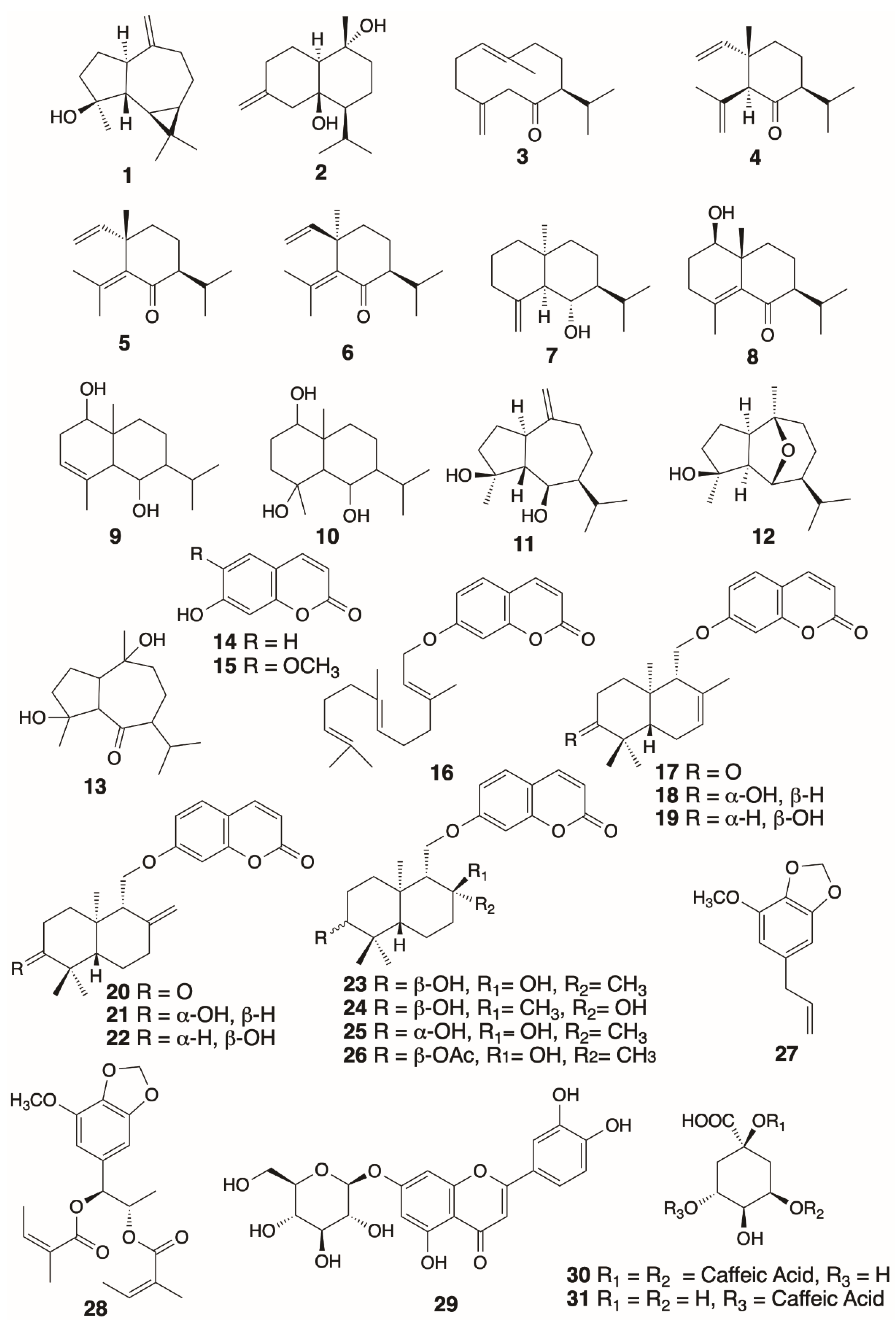
2.2. General Experimental Procedures, Extraction and Isolation of Secondary Metabolies
3. Results and Discussions
3.1. Comparison of Morphological/Organoleptic Characteristics
3.1.1. General Appearance
3.1.2. Fruits (Phyllon)
3.1.3. Oleo-Gum-Resin (Silphion)
3.1.4. Roots
3.1.5. Leaves (Maspetum)
3.2. Phylogenetic Classification Analyses of Ferula drudeana
3.3. Preliminary Chemical, Biological and Pharmacological Evaluations
3.4. Archaeobotanical Connections
3.5. Initial Conservation Studies
3.6. Reassessment of the Silphion Extinction Event in the Cyrenaic Region
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gemmill, C.L. Silphium. Bull. Hist. Med. 1966, 40, 295–313. Available online: https://www.jstor.org/stable/44447186 (accessed on 20 December 2020). [PubMed]
- Koerper, H.; Kolls, A.L. The Silphium motif adorning ancient Libyan coinage: Marketing a medicinal plant. Econ. Bot. 1999, 53, 133–143. [Google Scholar] [CrossRef]
- Bostock, J.; Riley, H.T. Book XXII, The Properties of Plants and Fruits, Chapter 49; Laser: Thirty-nine Remedies. In Pliny the Elder, Natural History; Collected Works of Pliny the Elder; Delphi Publishing Ltd., Delphi Classics, Hastings: East Sussex, UK, 2015. [Google Scholar]
- Rosengarten, F., Jr. The Book of Spices; Livingston Publishing Co.: Wynnewood, PA, USA, 1969; pp. 27–28. [Google Scholar]
- Bury, J.B. The Expansion of Greece, 7. Cyrene. In A History of Greece to the Death of Alexander the Great; Cambridge University Press: Cambridge, UK, 2015; pp. 116–117. [Google Scholar]
- Fage, J.D.; Oliver, R. The Greek Colonization of Cyrenaica; The Battiadai (c. 630 B.C. to c. 439 B.C.) and The Republic (c. 439 B.C. to c. 322 B.C.). In The Cambridge History of Africa, Volume 2, c. 500 B.C. to 1050 A.D.; Cambridge University Press: Cambridge, UK, 2008; pp. 107–114. [Google Scholar]
- Richardson, S. Libya Domestica: Libyan Trade and Society on the Eve of the Invasion of Egypt. J. Am. Res. Cent. Egypt 1999, 36, 149–164. [Google Scholar] [CrossRef]
- Totelin, L.M.V. Hippocratic Recipes, Oral and Written Transmission of Pharmacological Knowledge in Fifth- and Fourth-Century Greece; Brill: Leiden, The Netherlands, 2009; pp. 158–161. [Google Scholar]
- Hort, A.F. Book VI of Under-Shrubs [3.2–3.4]. In Theophrastus, Enquiry into Plants; Collected Works of Theophrastus; Delphi Publishing Ltd., Delphi Classics, Hastings: East Sussex, UK, 2019. [Google Scholar]
- Adams, C.D. Hippocrates, On Fistulae [9]. In Collected Works of Hippocrates; Delphi Publishing Ltd., Delphi Classics, Hastings: East Sussex, UK, 2015. [Google Scholar]
- Temkin, O. Book I, XIX. Whether One Ought to Make Use of Abortives and Contraceptives and How? In Soranus’ Gynecology; The Johns Hopkins University Press: Baltimore, MD, USA, 1956; p. 63. [Google Scholar]
- Scarborough, J. The drug lore of Asclepiades of Bithynia. Pharm. Hist. 1975, 17, 43–57. Available online: https://www.jstor.org/stable/41108902 (accessed on 20 December 2020). [PubMed]
- Bostock, J.; Riley, H.T. Book XIX, The Nature and Cultivation of Flax, and an Account of Various Garden Plants, Chapter 15, Laserpitium, Laser and Maspetum. In Pliny the Elder, Natural History; Collected Works of Pliny the Elder; Delphi Publishing Ltd., Delphi Classics, Hastings: East Sussex, UK, 2015. [Google Scholar]
- Gunther, R.T. The Greek Herbal of Dioscorides; Hafner Publishing Co.: London, UK; New York, NY, USA, 1968; pp. 323, 328–332. [Google Scholar]
- Osbaldeston, T.A.; Wood, R.P.A. 3-94. Silphion. In Dioscorides, De Materia Medica, a New Indexed Version in Modern English; Ibidis Press: Johannesburg, South Africa, 2000; pp. 475–479. [Google Scholar]
- Lykoudis, M. In Search of Silphion, Reading 17-1. 2008. Available online: https://hort.purdue.edu/newcrop/Hort_306/reading/Reading%2017-1.pdf (accessed on 20 December 2020).
- Riddle, J.M.; Worth Estes, J. Oral Contraceptives in Ancient and Medieval Times. Am. Sci. 1992, 80, 226–233. Available online: http://www.jstor.org/stable/29774642 (accessed on 20 December 2020).
- Parejko, K. Pliny the Elder’s Silphium: First Recorded Species Extinction. Conserv. Biol. 2003, 17, 925–927. [Google Scholar] [CrossRef]
- Roques, D. Synésios de Cyrène et le Silphion de Cyrénaïque. Rev. Études Grecques 1984, 97, 218–231. Available online: https://www.jstor.org/stable/44263808 (accessed on 20 December 2020). [CrossRef]
- Kiehn, M.S. Silphion: Eine in der Antike Ausgestorbene Nutzpflanze? Master’s Thesis, Universität Wien, Wien, Austria, 2006. [Google Scholar]
- Strantz, E. Zur Silphionfrage. Ph.D. Thesis, Universität Zürich, Zürich, Switzerland, 1909. [Google Scholar]
- Asciutti, V. The Silphium Plant: Analysis of Its Ancient Sources. Master’s Thesis, Durham University, Durham, UK, 2004. [Google Scholar]
- Kiehn, M. ΣΙΛΦΙOΝ—Silphion “...Multis iam Annis in ea Terra Non Invenitur...” Thessaloniki: European Botanic Gardens in a Changing World: Insights into Eurogard VI. 2014, pp. 33–41. Available online: http://www.botanicgardens.eu/eurogard/eurogard6/kiehn_silphion.pdf (accessed on 20 November 2020).
- Baytop, A. Plant collectors in Anatolia (Turkey). Phytol. Balc. 2010, 16, 187–213. Available online: http://www.bio.bas.bg/~phytolbalcan/PDF/16_2/16_2_03_Baytop.pdf (accessed on 20 December 2020).
- Korovin, E. Generis Ferula (Tourn.) L. Monographia Illustrate; Academia Scientiarum UzRSS: Taschkent, Uzbekistan, 1947. [Google Scholar]
- Sağıroğlu, M.; Duman, H. Rediscovery of Ferula anatolica and Ferula drudeana (Apiaceae) from Turkey. Biol. Divers. Conserv. 2011, 4, 191–197. Available online: https://dergipark.org.tr/en/pub/biodicon/issue/55889/765862 (accessed on 20 December 2020).
- Ellis, R.G. Ferula—Communis and Beyond. Bot. Soc. Br. Isles (BSBI) News 1998, 79, 64–66. Available online: http://archive.bsbi.org.uk/BSBINews79.pdf (accessed on 20 December 2020).
- Miski, M. Taxonomic Implications of the Sesquiterpenes of Ferula species. In Proceedings of the Book of Abstracts of 8th Apiales Symposium, Istanbul, Turkey, 4–8 August 2014; p. 16. [Google Scholar]
- Koerper, H.; Moerman, D.E. Coinage of Greek Cyrenaica, The Silphium Economy, and Exaggerated Advertising. Herb. Gram 2000, 48, 46–49. Available online: http://cms.herbalgram.org/herbalgram/pdfs/HG48-online.pdf (accessed on 20 November 2020).
- Piwczynski, M.; Wyborska, D.; Golebiewska, J.; Puchalka, R. Phylogenetic positions of seven poorly known species of Ferula (Apiaceae) with remarks on the phylogenetic utility of the plastid trnH-psbA, trnS-trnG, and atpB-rbcL intergenic spacers. Syst. Biodivers. 2018, 16, 428–440. [Google Scholar] [CrossRef]
- Inagaki, F.; Abe, A. Analysis of 1H and 13C Nuclear Magnetic Resonance Spectra of Spathulenol by Two-dimensional Methods. J. Chem. Soc. Perkin Trans. II 1985, 11, 1773–1778. [Google Scholar] [CrossRef]
- Ziaei, A.; Ramezani, M.; Wright, L.; Paetz, C.; Schneider, B.; Amirghofran, Z. Identification of Spathulenol in Salvia mirzayanii and the immunomodulatory effects. Phytother. Res. 2011, 25, 557–562. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, K.F.; Moreira, F.M.F.; Santos, J.A.; Kassuya, C.A.L.; Croda, J.H.R.; Cardoso, C.A.L.; do Carmo Vieira, M.; Ruiz, A.L.T.G.; Foglio, M.A.; de Carvalho, J.E.; et al. Antioxidant, anti-inflammatory, antiproliferative and antimycobacterial activities of the essential oil of Psidium guinness Sw. and spathulenol. J. Ethnopharmacol. 2018, 210, 351–358. [Google Scholar] [CrossRef]
- Martins, A.; Hajdu, Z.; Vasas, A.; Csupor-Löffler, B.; Molnar, J.; Hohmann, J. Spathulenol inhibit the human ABCB1 efflux pump. Planta Med. 2010, 76, 608. [Google Scholar] [CrossRef]
- Yamamura, S.; Iguchi, A.; Nishiyama, A.; Niwa, M.; Koyama, H.; Hirata, Y. Sesquiterpenes from Acorus calamus L. Tetrahedron 1971, 27, 5419–5431. [Google Scholar] [CrossRef]
- Zdero, C.; Bohlmann, F.; Solomon, J.C.; King, R.M.; Robinson, H. Ent-clerodane and other constituents from Bolivian Baccharis species. Phytochemistry 1989, 28, 531–542. [Google Scholar] [CrossRef]
- Niwa, M.; Terada, Y.; Iguchi, M.; Yamamura, S. Stereochemical studies on the elemene-type sesquiterpenes from Acorus calamus L. Chem. Lett. 1977, 6, 1415–1418. [Google Scholar] [CrossRef]
- Thomas, A.F.; Ozainne, M.; Decorzant, R.; Näf, F.; Lukacs, G. 10-Epijunenol, a New cis-Eudesmane Sesquiterpenoid. Tetrahedron 1976, 32, 2261–2264. [Google Scholar] [CrossRef]
- Thomas, A.F.; Ozainne, M. 272. New sesquiterpene alcohols from Galbanum resin: The occurrence of C(10)-epi-sesquiterpenoids. Helv. Chim. Acta 1978, 61, 2874–2880. [Google Scholar] [CrossRef]
- Nawakami, K.; Kuroyanagi, M. Sesquiterpenoids from Acorus calamus as germination inhibitors. Phytochemistry 1996, 43, 1175–1182. [Google Scholar] [CrossRef]
- Zheng, M.S.; Yang, J.H.; Li, Y.; Li, X.; Chang, H.W.; Son, J.-K. Anti-inflammatory activity of constituents isolated from Ulmus davidiana var. japonica. Biomol. Ther. 2010, 18, 321–328. [Google Scholar] [CrossRef][Green Version]
- Bruno, M.; De La Torre, M.C.; Rodriguez, B.; Omar, A.A. Guaiane sesquiterpenes from Teucrium leucocladum. Phytochemistry 1993, 34, 245–247. [Google Scholar] [CrossRef]
- Ahmed, A.A.; Hegazy, M.-E.F.; Hassan, N.M.; Wojcinska, M.; Karchesy, J.; Pare, P.W.; Mabry, T.J. Constituents of Chrysothamnus viscidiflorus. Phytochemistry 2006, 67, 1547–1553. [Google Scholar] [CrossRef]
- Ono, M.; Yamashita, M.; Mori, K.; Masuoka, C.; Ero, M.; Kinjo, J.; Ikeda, T.; Yoshimitsu, H.; Nohara, T. Sesquiterpenoids, triterpenoids, and flavonoids from the fruits of Schinus mole. Food Sci. Technol. Res. 2008, 14, 499–508. Available online: https://www.jstage.jst.go.jp/article/fstr/14/5/14_5_499/_pdf (accessed on 31 December 2020). [CrossRef]
- Roselli, S.; Maggio, A.; Bellone, G.; Formisano, C.; Basile, A.; Cicala, C.; Alfieri, A.; Mascolo, N.; Bruno, M. Antibacterial and anticoagulant activities of coumarins isolated from the flowers of Magydaris tomentosa. Planta Med. 2006, 72, 116–120. [Google Scholar] [CrossRef]
- Iranshahi, M.; Shahverdi, A.R.; Mirjani, R.; Amin, G.; Shafiee, A. Umbelliprenin from Ferula persica roots inhibits the red pigment production in Serratia marcescens. Z. Naturforsch. 2004, 59, 506–508. [Google Scholar] [CrossRef]
- Iranshahi, M.; Barthomeuf, C.; Bayet-Robert, M.; Chollet, P.; Davoodi, D.; Piacante, S.; Rezaee, R.; Sahebkar, A. Drimane-type sesquiterpene coumarins from Ferula gummosa fruits enhance doxorubicin uptake in doxorubicin-resistant human breast cancer cell line. J. Tradit. Complement. Med. 2014, 4, 118–125. [Google Scholar] [CrossRef]
- Tosun, F.; Beutler, J.A.; Ransom, T.T.; Miski, M. Anatolicin, a highly potent and selective cytotoxic sesquiterpene coumarin fron the root extract of Heptaptera anatolica. Molecules 2019, 24, 1153. [Google Scholar] [CrossRef]
- Tashkhodzaev, B.; Turgunov, K.K.; Izotova, L.Y.; Kamoldinov, K.S. Stereochemistry of samarcandin-type sesquiterpenoid coumarins. Crystal structures of feshurin and nevskin. Chem. Nat. Compd. 2015, 51, 242–246. [Google Scholar] [CrossRef]
- Malikov, V.M.; Saidkhodzhaev, A.I.; Aripov, K.N. Coumarins: Plants, structure, properties. Chem. Nat. Compd. 1998, 34, 202–264. [Google Scholar] [CrossRef]
- Zheng, G.-Q.; Kenney, P.M.; Lam, L.K.T. Myristicin: A potential cancer chemopreventive agent from parsley leaf oil. J. Agric. Food Chem. 1992, 40, 107–110. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, W. Anti-inflammatory effect of myristicin on RAW 264.7 macrophages stimulated with polyinosinic-polycytidylic acid. Molecules 2011, 16, 7132–7142. [Google Scholar] [CrossRef] [PubMed]
- Barrero, A.F.; Herrador, M.M.; Arteaga, P. Sesquiterpenes and phenylpropanoids from Seseli vayredanum. Phytochemistry 1992, 31, 203–207. [Google Scholar] [CrossRef]
- Miski, M.; Kürkçüoğlu, M.; İşcan, G.; Göger, F.; Tosun, F. Biological Activities of the Essential Oil, Fruit and Root Extracts of Ferula drudeana Korovin, 2013 ASP Annual Meeting, Abstract. Planta Med. 2013, 79, PN30. Available online: https://www.thieme-connect.com/products/ejournals/abstract/10.1055/s-0033-1348711 (accessed on 20 December 2020).
- Ziaei, A.; Amirghofran, Z.; Zapp, J.; Ramezani, M. Immunoinhibitory effect of teuclatriol a guaiane sesquiterpene from Salvia mirzayanii. Iran. J. Immunol. 2011, 8, 226–235. Available online: https://iji.sums.ac.ir/article_17030_d1ee180bd7518a26a0bc9248b1bc546b.pdf (accessed on 20 December 2020). [PubMed]
- Adhami, H.-R.; Fritz, V.; Lubich, A.; Kaehlig, H.; Zehl, M.; Krenn, L. Acetylcholinesterase inhibitors from galbanum, the oleo gum-resin of Ferula gummosa Boiss. Phytochem. Lett. 2014, 10, 82–87. [Google Scholar] [CrossRef]
- Bustanji, Y.; Issa, A.; Mohammad, M.; Hudaib, M.; Tawah, K.; Alkhatib, H.; Almasri, I.; Al-Khalidi, B. Inhibition of hormone sensitive lipase and pancreatic lipase by Rosmarinus officinalis extract and selected phenolic constituents. J. Med. Plant Res. 2010, 4, 2235–2242. [Google Scholar] [CrossRef]
- Zaugg, J.; Eickmeier, E.; Ebrahimi, S.N.; Baburin, I.; Hering, S.; Hamburger, M. Positive GABAA Receptor Modulators from Acorus calamus and Structural Analysis of (+)-Dioxosarcoguaiacol by 1D and 2D NMR and Moleculer Modeling. J. Nat. Prod. 2011, 74, 1437–1443. [Google Scholar] [CrossRef]
- Ziaei, A.; Hoppstadter, J.; Kiemer, A.K.; Ramezani, M.; Amirghofran, Z.; Diesel, B. Inhibitory effects of teuclatriol, a sesquiterpene from Salvia mirzayanii, on nuclear factor-kB activation and expression of inflammatory mediators. J. Ethnopharmacol. 2015, 160, 94–100. [Google Scholar] [CrossRef]
- Park, Y.J.; Zheng, H.; Kwak, J.H.; Chung, K.H. Sesquiterpenes from Cyperus rotundus and 4a,5a-oxido-eudesm-11-en-3-one as a potential selective estrogen receptor modulator. Biomed. Pharmacother. 2019, 109, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- Swetha, D.; Harika, K.; Preetha, C.; Vamsi Krishna, C.; Puzahenthi, R. A Review on the cardio protective activity of artichoke. J. Int. Pharm. Sci. 2017, 4, 44–51. Available online: http://jipsjournal.in/articles/JIPSDEC2017V4-I201.pdf (accessed on 20 November 2020).
- Olas, B.; Brys, M. Is it safe to use Acorus calamus as a source of promising bioactive compounds in prevention and treatment of cardiovascular diseases? Chem. Biol. Interact. 2018, 281, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Alqarni, M.H.; Soliman, G.A.; Salkini, M.A.A.; Alam, P.; Yusufoglu, H.S.; Baykan, S.; Öztürk, B.; Abdel-Kader, M.S. The Potential Aphrodisiac Effect of Ferula drudeana Korovin Extracts and Isolated Sesquiterpene Coumarins in Male Rats. Pharmacogn. Mag. 2020, 16, 404. [Google Scholar] [CrossRef]
- Dos Santos, E.; Radai, J.A.S.; do Nascimento, K.F.; Formagio, A.S.N.; de Matos Balsalobre, N.; Ziff, E.B.; Konkiewitz, E.C.; Kassuya, C.A.L. Contribution of spathulenol to the anti-nociceptive effects of Psidium guineense. Nutr. Neurosci. 2020, 1815330. [Google Scholar] [CrossRef]
- Cavar, S.; Maksimovic, M.; Šolic, M.E.; Jerkovic-Mujkic, A.; Bešta, R. Chemical composition and antioxidant and antimicrobial activity of two Satureja essential oils. Food Chem. 2008, 111, 648–653. [Google Scholar] [CrossRef]
- Lou, L.-L.; Li, W.; Zhou, B.-H.; Chen, L.; Weng, H.-Z.; Zou, Y.-H.; Tang, G.-H.; Bu, X.-Z.; Yin, S. (+)-Isobicyclogermacrenal and spathulenol from Aristolochia yunnanensis alleviate cardiac fibrosis by inhibiting transforming growth factor b/small mother against decapentaplegic signaling pathway. Phytother. Res. 2019, 33, 214–223. [Google Scholar] [CrossRef]
- Menichini, F.; Conforti, F.; Rigano, D.; Formisano, C.; Piozzi, F.; Senatore, F. Phytochemical composition, anti-inflammatory and antitumour activities of four Teucrium essential oils from Greece. Food Chem. 2009, 115, 679–686. [Google Scholar] [CrossRef]
- Chen, H.-P.; Yang, K.; Zheng, L.-S.; You, C.-X.; Cai, Q.; Wang, C.-F. Repellant and insecticidal activities of shyobunone and isoshyobunone derived from the essential oil of Acorus calamus rhizomes. Pharmacogn. Mag. 2015, 11, 675–681. [Google Scholar] [CrossRef]
- Ezzai, S.M.; Motaal, A.A. Isolation of new cytotoxic metabolites from Cleome droserifolia growing in Egypt. Z. Naturforsch. 2012, 67, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-L.; Yang, X.-W.; Wu, B.-F.; Shang, J.-H.; Liu, Y.-P.; Luo, Z.D.; Luo, X.-D. Anti-inflammatory effect of pomelo peel and its bioactive coumarins. J. Agric. Food Chem. 2019, 67, 8810–8818. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Hozayen, W.G.; Hasan, I.H.; Shaban, E.; Bin-Jumah, M. Umbelliferone ameliorates CCl4-induced liver fibrosis in rats by upregulating PPARg and attenuating oxidative stress, inflammation, and TGF-b1/Smad3 signaling. Inflammation 2019, 42, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.C.; Baek, J.M.; Lee, C.H.; Yoon, K.-H.; Lee, M.S.; Kim, J.-Y. Umbelliferone prevents lipopolysaccharide-induced bone loss and suppresses RANKL-induced osteoclastogenesis by attenuating Akt-c-Fos-NFATc1 signaling. Int. J. Biol. Sci. 2019, 15, 2427–2437. [Google Scholar] [CrossRef]
- Karakaya, S.; Yılmaz-Oral, D.; Kilic, C.S.; Gur, S. Umbelliferone isolated from Zosima absinthifolia roots partially restored erectile dysfunction in streptozicin-induced diabetic rats. Med. Chem. Res. 2019, 28, 1161–1167. [Google Scholar] [CrossRef]
- Singh, R.; Singh, B.; Singh, S.; Kumar, N.; Kumar, S.; Arora, S. Umbelliferone—An antioxidant isolated from Acacia nilotica (L.) Willd. Ex. Del. Food Chem. 2010, 120, 825–830. [Google Scholar] [CrossRef]
- Alam, M.; Khan, A.; Wadood, A.; Bashir, S.; Aman, A.; Farooq, U.; Khan, F.A.; Mabood, F.; Hussain, J.; Al-Harrasi, S.A. Bioassay-guided isolation of urease inhibitors from Ferula narthex Boiss. S. Afr. J. Bot. 2019, 120, 247–252. [Google Scholar] [CrossRef]
- Radha, G.V.; Sadhana, B.; Trideva Sastri, K.; Ganapaty, S. Bioactive umbelliferone and its derivatives: An update. J. Pharmacogn. Phytochem. 2019, 8, 59–66. Available online: https://www.phytojournal.com/archives/?year=2019&vol=8&issue=1&ArticleId=6701 (accessed on 31 December 2020).
- Pan, R.; Dai, Y.; Gao, X.-H.; Lu, D.; Xia, Y.-F. Inhibition of vascular endothelial growth factor-induced angiogenesis by scopoletin through interrupting the autophosphorylation of VEGF receptor 2 and its downstream signaling pathways. Vasc. Pharmacol. 2011, 54, 18–28. [Google Scholar] [CrossRef]
- Kang, S.Y.; Sung, S.H.; Park, J.H.; Kim, Y.C. Hepatoprotective activity of scopoletin, a constituent of Solanum lyratum. Arch. Pharm. Res. 1998, 21, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Gao, X.-H.; Lu, H.; Xu, X.-X.; Xia, Y.-F.; Dai, Y. Prevention of FGF-2-induced angiogenesis by scopoletin, a coumarin compound isolated from Erycibe obtusifolia Benth., and its mechanism of action. Int. Immunopharmacol. 2011, 11, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Adfa, M.; Yoshimura, T.; Komura, K.; Koketsu, M. Antitermite activities of coumarin derivatives and scopoletin from Protium javanicum Burm. f. J. Chem. Ecol. 2010, 36, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Napiroon, T.; Bacher, M.; Balslev, H.; Tawaitakham, K.; Santimaleeworagun, W.; Vajrodaya, S. Scopoletin from Lasianthus lucidus Blume (Rubiaceae): A potential antimicrobial against multi-drug resistant Pseudomonas aeruginosa. J. Appl. Pharm. Sci. 2018, 8, 1–6. [Google Scholar] [CrossRef]
- Luo, J.; Lai, T.; Guo, T.; Chen, F.; Linli, Z.; Ding, W.; Zhang, Y. Synthesis and acaricidal activities of scopoletin phenolic ether derivatives: QSAR, molecular docking study and in silico ADME predictions. Molecules 2018, 23, 995. [Google Scholar] [CrossRef] [PubMed]
- Pandy, V.; Narasingam, M.; Kunasegaran, T.; Murugan, D.D.; Mohamed, Z. Effect of Noni (Morinda citrifolia Linn.) fruit and its bioactive principles scopoletin and rutin on rat vas deferens contractility: An ex vivo study. Sci. World J. 2014, 909586. [Google Scholar] [CrossRef]
- Rollinger, J.M.; Hornick, A.; Langer, T.; Stuppner, H.; Prast, H. Acetylcholinesterase inhibitor activity of scopolin and scopoletin discovered by virtual screening of natural products. J. Med. Chem. 2004, 47, 6248–6254. [Google Scholar] [CrossRef]
- Tabana, Y.M.; Hassan, L.E.A.; Ahamed, M.B.K.; Dahham, S.S.; Iqbal, M.A.; Saeed, M.A.A.; Khan, M.S.S.; Sandai, D.; Majid, A.S.A.; Oon, C.E.; et al. Scopoletin, an active free principle of tree tobacco (Nicotiana glauca) inhibits human tumor vascularization in xenograft models and modulates ERK1, VEGF-A, and FGF-2 in computer model. Microvasc. Res. 2016, 107, 17–33. [Google Scholar] [CrossRef]
- Pandy, V.; Vijeepallam, K. Antipsychotic-like activity of scopoletin and rutin against the positive symptoms of schizophrenia in mouse models. Exp. Anim. 2017, 66, 417–423. Available online: https://www.jstage.jst.go.jp/article/expanim/66/4/66_17-0050/_pdf/-char/en (accessed on 30 December 2020). [CrossRef][Green Version]
- Choi, R.-Y.; Ham, J.R.; Lee, H.-I.; Cho, H.W.; Choi, M.-S.; Park, S.-K.; Lee, J.; Kim, M.-J.; Seo, K.-I.; Lee, M.-K. Scopoletin supplementation ameliorates steatosis and inflammation in diabetic mouse. Phytother. Res. 2017, 31, 1795–1804. [Google Scholar] [CrossRef]
- Mogana, R.; Adhikari, A.; Tzar, M.N.; Ramliza, R.; Viart, C. Antibacterial activities of the extracts, fractions and isolated compounds from Canarium patentinervium Miq. against bacterial clinical isolates. BMC Complement. Med. Ther. 2020, 20, 55. [Google Scholar] [CrossRef]
- Soltani, F.; Mosaffa, F.; Iranshahi, M.; Karimi, G.; Malekaneh, M.; Haghighi, F.; Behravan, J. Evaluation of antigenotoxicity effects of umbelliprenin on human peripheral lymphocytes exposed to oxidative stress. Cell Biol. Toxicol. 2009, 25, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Iranshahi, M.; Askari, M.; Sahebkar, A.; Hadjipavlou-Litina, D. Evaluation of antioxidant, anti-inflammatory and lipoxygenase inhibitory activities of the prenylated coumarin umbelliprenin. DARU J. Pharm. Sci. 2009, 17, 99–103. Available online: http://daru.tums.ac.ir/index.php/daru/article/view/530 (accessed on 30 December 2020).
- Shahverdi, A.R.; Saadat, F.; Khorramizadeh, M.R.; Iranshahi, M.; Khoshayand, M.R. Two matrix metalloproteinases inhibitors from Ferula persica var. persica. Phytomedicine 2006, 13, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Shahzadi, I.; Ali, Z.; Baek, S.H.; Mirza, B.; Ahn, K.S. Assessment of the antitumor potential of umbelliprenin, a naturally occurring sesquiterpene coumarin. Biomedicines 2020, 8, 126. [Google Scholar] [CrossRef]
- Valiahdi, S.M.; Iranshahi, M.; Sahebkar, A. Cytotoxic activities of phytochemicals from Ferula species. DARU J. Pharm. Sci. 2013, 21, 39. [Google Scholar] [CrossRef]
- Shakeri, A.; Iranshahi, M.; Iranshahi, M. Biological properties and molecular targets of umbelliprenin—A mini-review. J. Asian Nat. Prod. Res. 2014, 16, 884–889. [Google Scholar] [CrossRef]
- Fiorito, S.; Epifano, F.; Preziuso, F.; Cacciatore, I.; di Stefano, A.; Taddeo, V.A.; de Medina, P.; Genovese, S. Natural oxyprenylated coumarins are modulators of melanogenesis. Eur. J. Med. Chem. 2018, 152, 274–282. [Google Scholar] [CrossRef]
- Hashemzaei, M.; Dousti, T.; Tsarouhas, K.; Bagheri, G.; Nikolouzakis, T.K.; Rezaee, R.; Shakraki, J. Effect of umbelliprenin on blood pressure in high-fat diet hypertensive rats. Farmacia 2020, 68, 447–452. [Google Scholar] [CrossRef]
- Iranshahi, M.; Kalategi, F.; Rezaee, R.; Shahverdi, A.R.; Ito, C.; Furukawa, H.; Tokuda, H.; Itoigawa, M. Cancer chemopreventive activity of terpenoid coumarins from Ferula species. Planta Med. 2008, 74, 147–150. [Google Scholar] [CrossRef]
- Majnooni, M.B.; Fakhri, S.; Smeriglio, A.; Trombetta, D.; Croley, C.R.; Bhattacharyya, P.; Sobarzo-Sánchez, E.; Farzaei, M.H.; Bishayee, A. Antiangiogenic effects of coumarins against cancer: From chemistry to medicine. Molecules 2019, 24, 4278. [Google Scholar] [CrossRef]
- Rashidi, M.; Khalilnezhad, A.; Amani, D.; Jamshidi, H.; Muhammadnejad, A.; Bazi, A.; Ziai, S.A. Umbelliprenin shows antitumor, antiangiogenesis, antimetastatic, anti-inflammatory, and immunostimulatory activities in 4T1 tumor-bearing Balb/c mice. J. Cell. Physiol. 2018, 1–11. [Google Scholar] [CrossRef]
- Fazly Bazzaz, B.S.; Iranshahi, M.; Naderinasab, M.; Hajian, S.; Sabeti, Z.; Masumi, E. Evaluation of the effects of galbanic acid from Ferula szowitsiana and conferol from F. badrakema, as modulators of multi-drug resistance in clinical isolates of Escherichia coli and Staphylococcus aureus. Res. Pharm. Sci. 2010, 5, 21–28. Available online: http://www.rps.mui.ac.ir/index.php/jrps/article/view/72 (accessed on 30 December 2020). [PubMed]
- Bashir, S.; Alam, M.; Adhikari, A.; Shrestha, R.L.; Yousuf, S.; Ahmad, B.; Parveen, S.; Aman, A.; Choudhary, M.I. New antileishmanial sesquiterpene coumarins from Ferula narthex Boiss. Phytochem. Lett. 2014, 9, 46–50. [Google Scholar] [CrossRef]
- Lee, C.-L.; Chiang, L.C.; Cheng, L.H.; Liaw, C.-C.; Abd El-Razek, M.; Chang, F.-R.; Wu, Y.C. Influenza A (H1N1) antiviral and cytotoxic agents from Ferula assa-foetida. J. Nat. Prod. 2009, 72, 1568–1572. [Google Scholar] [CrossRef] [PubMed]
- Jabrane, A.; Ben Jannet, H.; Mighri, Z.; Mirjolet, J.-F.; Duchamp, O.; Harzallah-Skhiri, F.; Lacaille-Dubois, M.-A. Two New Sesquiterpene Derivatives from the Tunisian Endemic Ferula tunetana Pom. Chem. Biodivers. 2010, 7, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Collu, M. Endocannabinoid System Modulation by Natural Products from Ancient Medicinal Plants. Ph.D. Thesis, University of Cagliari, Cagliari, Italy, 2019. [Google Scholar]
- Özbek, H.; Güvenalp, Z.; Yılmaz, G.; Yerdelen, K.Ö.; Kazaz, C.; Demirezer, Ö. In vitro anticholinesterase activity and molecular docking studies of coumarin derivatives isolated from roots of Heptaptera cilicica. Med. Chem. Res. 2018, 27, 538–545. [Google Scholar] [CrossRef]
- Ghoran, S.H.; Atabaki, V.; Babaei, E.; Olfatkhah, S.R.; Dusek, M.; Eigner, V.; Soltani, A.; Khalaji, D. Isolation, spectroscopic characterization, X-ray, theoretical studies as well as in vitro cytotoxicity of samarcandin. Bioorg. Chem. 2016, 66, 27–32. [Google Scholar] [CrossRef]
- NCI Samarcandin Yeast Assay Results. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5459231#section=Biological-Test-Results (accessed on 31 December 2020).
- Sethur, A.S.; Naik, S.Y.; Skariyachan, S. Herbal lead as ideal bioactive compounds against probable drug targets of Ebola virus in comparison with known chemical analogue: A computational drug discovery perspective. Interdiscip. Sci. Comput. Life Sci. 2016, 9, 254–277. [Google Scholar] [CrossRef]
- Dapiaggi, F.; Pieraccini, S.; Potenza, D.; Vasile, F.; Podlipnik, C. Designing antiviral substances targeting the Ebola virus viral protein 24. In Emerging and Reemerging Viral Pathogens. Vol. 2; Ennaji, M.M., Ed.; Academic Press: London, UK, 2020; pp. 147–177. [Google Scholar] [CrossRef]
- Nilawati, A.; Atmajaningtyas, A.T.J.; Ansory, H.M. The influence of myristicin lost in Myristica fragrans volatile oils to antimicrobial activity against B. subtilis, E. coli and S. aureus. In Proceedings of the 2nd Bakti Tunas Husada-Health Science International Conference (BTH-HSIC 2019), Tasikmalaya, Indonesia, 5–6 October 2019. [Google Scholar] [CrossRef]
- Morita, T.; Jinno, K.; Kawagishi, H.; Arimoto, Y.; Suganuma, H.; Inakuma, T.; Sugiyama, K. Hepatoprotective effect of myristicin from nutmeg (Myristica fragrans) on lipopolysaccharide/D-galactosamine-induced liver injury. J. Agric. Food Chem. 2003, 51, 1560–1565. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, C.; Shen, X.; Xiao, L.; Wang, H.; Liu, P.; Wang, L.; Xu, H. Cytoprotective effects of myristicin against hypoxia-induced apoptosis and endoplasmic reticulum stress in rat dorsal root ganglion neurons. Mol. Med. Rep. 2017, 15, 2280–2288. [Google Scholar] [CrossRef]
- Qiburi, Q.; Ganbold, T.; Bao, Q.; Da, M.; Aoqier, A.; Temuqile, T.; Baigude, H. Bioactive components of ethnomedicine Eerdun Wurille regulate the transcription of pro-inflammatory cytokines in microglia. J. Ethnopharmacol. 2020, 246, 112241. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.A.; Andrade, L.N.; de Sousa, E.B.V.; de Sousa, D.P. Antitumor phenylpropanoids found in essential oils. Biomed. Res. Int. 2015, 392674. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, P.; Kumar, P.; Singh, V.K.; Singh, D.K. Enzyme inhibition by molluscicidal components of Myristica fragrans Houtt. In the nervous tissue of snail Lymnea acuminate. Enzym. Res. 2010, 478746. [Google Scholar] [CrossRef]
- Pandey, R.; Mahar, R.; Hasanain, M.; Shukla, S.K.; Sarkar, J.; Rameshkumar, K.B.; Kumar, B. Rapid screening and quantitative determination of bioactive compounds from fruit extracts of Myristica species and their in vitro antiproliferative activity. Food Chem. 2016, 211, 483–493. [Google Scholar] [CrossRef]
- Abou-Elnaga, Z.S. Insecticidal bioactivity of eco-friendly plant origin chemicals against Culex pipiens and Aedes aegypti (Diptera: Culicidae). J. Entomol. Zool. Stud. 2014, 2, 340–347. Available online: https://www.entomoljournal.com/archives/?year=2014&vol=2&issue=6&ArticleId=470 (accessed on 31 December 2020).
- Yang, R.-L.; Yan, Z.-H.; Lu, Y. Cytotoxic phenylpropanoids from carrot. J. Agric. Food Chem. 2008, 56, 3024–3027. [Google Scholar] [CrossRef] [PubMed]
- Garg, C.; Khurana, P.; Garg, M. Molecular mechanism of skin photoaging and plant inhibitors. Int. J. Green Pharm. 2017, 11, S217. [Google Scholar] [CrossRef]
- Gebhardt, R. Choleretic and anticholestatic activities of flavonoids of artichoke (Cynara scolymus L. subsp. scolymus (L.) Hayek). Acta Hort. 2005, 681, 429–435. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Chirikova, N.K.; Vennos, C. Chemical composition, antioxidant and anticholinesterase activities of Gentianella azurea from Russian Federation. Nat. Prod. Commun. 2017, 12, 55–56. [Google Scholar] [CrossRef]
- Odontoya, G.; Hoult, J.R.S.; Houghton, P.J. Structure-activity relationship for anti-inflammatory effect of luteolin and its derived glycosides. Phytother. Res. 2005, 19, 782–786. [Google Scholar] [CrossRef]
- Chung, H.S. Inhibition of monoamine oxidase by a flavone and its glycoside from Ixeris dentata Nakai. Nutraceuticals Food 2003, 8, 141–144. [Google Scholar] [CrossRef]
- Brown, J.E.; Rice-Evans, C.A. Luteolin-Rich artichoke extract protects low density lipoprotein from oxidation in vitro. Free Rad. Res. 1998, 29, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhang, H.; Lo, R. Phenolic compounds from the leaf extract of artichoke (Cynara scolymus L.) and their antimicrobial activities. J. Agric. Food Chem. 2004, 52, 7272–7278. [Google Scholar] [CrossRef] [PubMed]
- Adzet, T.; Camarasa, J.; Laguna, J.C. Hepatoprotective activity of polyphenolic compounds from Cynara scolymus against CCl4 toxicity in isolated rat hepatocytes. J. Nat. Prod. 1987, 50, 612–617. [Google Scholar] [CrossRef] [PubMed]
- Hakkou, Z.; Maciuk, A.; Leblais, V.; Bounani, N.E.; Mekhfi, H.; Bnouham, M.; Aziz, M.; Ziyyat, A.; Rauf, A.; Hadda, T.B.; et al. Antihypertensive and vasodilator effects of methanolic extract of Inula viscosa: Biological evaluation and POM analysis of cynarin, chlorogenic acid as potential hypertensive. Biomed. Pharmacother. 2017, 93, 62–69. [Google Scholar] [CrossRef]
- Kirchhoff, R.; Beckers, C.; Kirchhoff, G.M.; Trinczek-Gärtner, H.; Petrowicz, O.; Reimann, H.J. Increase in choleresis by means of artichoke extract. Phytomedicine 1994, 1, 107–115. [Google Scholar] [CrossRef]
- Mileo, A.M.; Di Venere, D.; Linsalata, V.; Fraioli, R.; Miccadei, S. Artichoke polyphenols induce apoptosis and decrease the invasive potential of the human breast cancer cell line MDA-MB231. J. Cell. Physiol. 2012, 227, 3301–3309. [Google Scholar] [CrossRef]
- Hwang, S.J.; Kim, Y.-W.; Park, Y.; Lee, H.J.; Kim, K.-W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide-stimulated RAW 264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef]
- Jin, S.; Chang, C.; Zhang, L.; Liu, Y.; Huang, X.; Chen, Z. Chlorogenic acid improves late diabetes through adiponectin receptor signaling pathways in db/db mice. PLoS ONE 2015, 10, e0120842. [Google Scholar] [CrossRef]
- Wu, D.; Bao, C.; Li, L.; Fu, M.; Wang, D.; Xie, J.; Gong, X. Chlorogenic acid protects against cholestatic liver injury in rats. J. Pharm. Sci. 2015, 129, 177–182. [Google Scholar] [CrossRef]
- Kwon, S.-H.; Lee, H.-K.; Kim, J.-A.; Hong, S.-I.; Kim, H.-C.; Jo, T.-H.; Park, Y.-I.; Lee, C.-K.; Kim, Y.-B.; Lee, S.-Y.; et al. Neuroprotective effects of chlorogenic acid on scopolamine-induced amnesia via anti-acetylcholinesterase and anti-oxidative activities in mice. Eur. J. Pharmacol. 2010, 649, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Cao, Z.; Cao, L.; Ding, G.; Wang, Z.; Xiao, W. Antiviral activity of chlorogenic acid against influenza A (H1N1/H3N2) virus and its inhibition of neuraminidase. Sci. Rep. 2017, 7, 45723. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.W.; Hsu, A.; Tan, B.K.H. Anti-diabetic and anti-lipidemic effects of chlorogenic acid are mediated by ampk activation. Biochem. Pharmacol. 2013, 85, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Liu, N.; Hou, N.; Dong, L.; Li, J. Chlorogenic acid inhibits hepatocellular carcinoma in vitro and in vivo. J. Nutr. Biochem. 2017, 46, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Rammal, H.; Dicko, A.; Younos, C.; Soulimani, R. Chlorogenic acid, a polyphenol from Prunus domestica (Mirabelle), with coupled anxiolytic and antioxidant effects. J. Neurol. Sci. 2007, 262, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Bagdas, D.; Cinkilic, N.; Ozboluk, H.Y.; Ozyigit, M.O.; Gurun, M.S. Antihyperalgesic activity of chlorogenic acid in experimental neuropathic pain. J. Nat. Med. 2012, 67, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Mubarak, A.; Bondonno, C.P.; Liu, A.H.; Considine, M.J.; Rich, L.; Mas, E.; Croft, K.D.; Hodgson, J.M. Acute effects of chlorogenic acid on nitric oxide status, endothelial function, and blood pressure in healthy volunteers: A randomized trial. J. Agric. Food Chem. 2012, 60, 9130–9136. [Google Scholar] [CrossRef]
- Heitman, E.; Ingram, D.K. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, M.; Liu, D. Chlorogenic acid improves high fat diet-induced hepatic steatosis and insulin resistance in mice. Pharm. Res. 2015, 32, 1200–1209. [Google Scholar] [CrossRef]
- Wang, Z.; Lam, K.-L.; Hu, J.; Ge, S.; Zhou, A.; Zheng, B.; Zeng, S.; Lin, S. Chlorogenic acid alleviates obesity and modulates gut microbiota in high-fat-fed mice. Food Sci. Nutr. 2019, 7, 579–588. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, D.; Wan, X.; Bai, Y.; Yuan, C.; Wang, T.; Yuan, D.; Zhang, C.; Liu, C. Chlorogenic acid suppresses miR-155 and ameliorates ulcerative colitis through NF-kB/NLRP3 inflammasome pathway. Mol. Nutr. Food Res. 2020, 64, 2000452. [Google Scholar] [CrossRef] [PubMed]
- Xue, N.; Zhou, Q.; Ji, M.; Jin, J.; Lai, F.; Chen, J.; Zhang, M.; Jia, J.; Yang, H.; Zhang, J.; et al. Chlorogenic acid inhibits glioblastoma growth through repolarizating macrophage from M2 to M1 phenotype. Sci. Rep. 2017, 7, 39011. [Google Scholar] [CrossRef] [PubMed]
- Changizi, Z.; Moslehi, A.; Rohani, A.H.; Eidi, A. Chlorogenic acid induces 4T1 breast cancer tumor’s apoptosis via p53, Bax, Bcl-2, and caspase-3 signaling pathways in BALB/c mice. J. Biochem. Mol. Toxicol. 2020, e22642. [Google Scholar] [CrossRef] [PubMed]
- Jin, U.-H.; Lee, J.-Y.; Kang, S.-K.; Kim, J.-K.; Park, W.-H.; Kim, J.-G.; Moon, S.-K.; Kim, C.-H. A phenolic compound, 5-caffeoylquinic acid (chlorogenic acid), is a new type and strong matrix metalloproteinase-9 inhibitor: Isolation and identification from methanol extract of Euonymus alatus. Life Sci. 2005, 77, 2760–2769. [Google Scholar] [CrossRef] [PubMed]
- Kiehn, M. Neues zum Silphion der Antike. Z. Phytother. 2009, 30, 83–87. [Google Scholar] [CrossRef]
- Dokovic, D.D.; Bulatovic, V.M.; Bozic, B.D.; Kataranovski, M.V.; Zrakic, T.M.; Kovacevic, N.N. 3,5-Nonadiyne Isolated from the Rhizome of Cachrys ferulacea Inhibits Endogenous Nitric Oxide Release by Rat Peritoneal Macrophages. Chem. Pharm. Bull. 2004, 52, 853–854. [Google Scholar] [CrossRef]
- Miski, M.; Ulubelen, A.; Mabry, T.J.; Watson, W.H.; Vickovic, I.; Holub, M. A new sesquiterpene ester from Ferula tingitana. Tetrahedron 1984, 24, 5197–5201. [Google Scholar] [CrossRef]
- Miski, M.; Mabry, T.J. Daucane esters from Ferula communis subsp. communis. Phytochemistry 1985, 24, 1735–1741. [Google Scholar] [CrossRef]
- Miski, M.; Mabry, T.J. Fercolide, a type of sesquiterpene lactone from Ferula communis subsp. communis and the correct structure of vaginatin. Phytochemistry 1986, 25, 1673–1675. [Google Scholar] [CrossRef]
- Miski, M.; Mabry, T.J. New daucane esters from Ferula tingitana. J. Nat. Prod. 1986, 49, 657–660. [Google Scholar] [CrossRef]
- Miski, M.; Jakupovic, J. Cyclic farnesyl-coumarin and farnesyl-chromone derivatives from Ferula communis subsp. communis. Phytochemistry 1990, 29, 1995–1998. [Google Scholar] [CrossRef]
- Znati, M.; Filali, I.; Jabrane, A.; Casanova, J.; Bouajila, J.; Ben Jannet, H. Chemical composition and in vitro evaluation of antimicrobial antioxidant and antigerminative properties of the seed oil from the Tunisian endemic Ferula tunetana Pom. ex Batt. Chem. Biodivers. 2017, 14, e1600116. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A. Sesquiterpene coumarins and sesquiterpenes from Ferula sinaica. Phytochemistry 1999, 50, 109–112. [Google Scholar] [CrossRef]
- Valle, M.G.; Appendino, G.; Nano, G.M.; Picci, V. Prenylated Coumarins and Sesquiterpenoids from Ferula communis. Phytochemistry 1987, 26, 253–256. [Google Scholar] [CrossRef]
- Lamnaouer, D.; Bodo, B.; Martin, M.-T.; Molho, D. Ferulenol and ω-Hydroxyferulenol, Toxic Coumarins from Ferula communis var. genuina. Phytochemistry 1987, 26, 1613–1615. [Google Scholar] [CrossRef]
- Andersen, T.B.; Lopez, C.Q.; Manczak, T.; Martinez, K.; Simonsen, H.T. Thapsigargin—From Thapsia L. to Mipsagargin. Molecules 2015, 20, 6113–6127. [Google Scholar] [CrossRef]
- Benson, G.G.; Hemingway, S.R.; Leach, F.N. The analysis of wrappings of mummy 1770. In The Manchester Museum Mummy Project: Multidisciplinary Research on Ancient Egyptian Mummified Remains; David, A.R., Ed.; Manchester Museum: Manchester, UK, 1979; pp. 119–132. Available online: http://assets.mhs.manchester.ac.uk/Mummy1770/Articles/Multidisciplinary-Research-on-Ancient-Egyptian-Mummified-Remains.pdf (accessed on 20 December 2020).
- Oras, E.; Anderson, J.; Törv, M.; Vahur, S.; Rammo, R.; Remmer, S.; Mölder, M.; Malve, M.; Saag, L.; Saage, R.; et al. Multidisciplinary investigation of two Egyptian child mummies curated at the University of Tartu Art Museum, Estonia (Late/Graeco-Roman Periods). PLoS ONE 2020, 15, e0227446. [Google Scholar] [CrossRef]
- D’Agostino, A.; Gismondi, A.; Di Marco, G.; Castro, M.L.; Olevano, R.; Tiziano, C.; Leonardi, D.; Canini, A. Lifestyle of a Roman Imperial community: Ethnobotanical evidence from dental calculus of the Ager Curensis inhabitants. J. Ethnobiol. Ethnomed. 2019, 15, 62. [Google Scholar] [CrossRef]
- Benchabane, O. Chemical composition and insecticidal activities of essential oils of two Algerian endemic plants: Ferula vesceritensis Coss. et Dur. and Thymus pallescens de Noe. Int. J. Agric. Sci. Res. 2014, 4, 185–191. Available online: http://www.tjprc.org/publishpapers/2-50-1417685353-20.%20Agri%20Sci%20%20-%20IJASR%20-%20%20%20CHEMICAL%20COMPOSITION%20AND%20%20-%20Otmane%20BNCHABANE.pdf (accessed on 20 December 2020).
- Labed-Zouad, I.; Labed, A.; Laggoune, S.; Zahia, S.; Kabouche, A.; Kabouche, Z. Chemical compositions and antibacterial activity of four essential oils from Ferula vesceritensis Coss. & Dur. Against clinical isolated and food-borne pathogens. Rec. Nat. Prod. 2015, 9, 518–525. Available online: http://www.acgpubs.org/doc/2018080722431863-RNP-EO_1310-067.pdf (accessed on 20 December 2020).
- Bouraoua, A.; Ferhat, M.; Kabouche, A.; Laggoune, S.; Touzani, R.; Kabouche, Z. Comparative compositions of essential oils of Ferula. J. Mater. Environ. Sci. 2014, 5, 1214–1217. Available online: http://www.jmaterenvironsci.com/Document/vol5/vol5_N4/150-JMES-839-2014-Bouratoua.pdf (accessed on 31 December 2020).
- Dehak-Oughlissi, K.; Hammoudi, R.; Hadj-Mahammed, M.; Badjah-Hadj-Ahmed, Y.A. Analyse de L’Huile Essentielle des Parties Aeriennes de Ferula vesceritensis Coss. Et Dur. de la Localite de Sebseb. Ann. Sci. Technol. 2013, 167–173. Available online: http://docplayer.fr/83366872-Analyse-de-l-huile-essentielle-des-parties-aeriennes-de-ferula-vesceritensis-coss-et-dur-de-la-localite-de-sebseb.html (accessed on 20 December 2020).
- Zellagui, A.; Gherraf, N.; Rhouati, S. Chemical composition and antibacterial activitiy of the essential oils of F. vesceritensis Coss et Dur. leaves, endemic in Algeria. Org. Med. Chem. Lett. 2012, 2, 31. [Google Scholar] [CrossRef] [PubMed]
- Kanani, M.R.; Rahiminejad, M.R.; Sonboli, A.; Mozaffarian, V.; Osaloo, S.K.; Ebrahimi, S.N. Chemotaxonomic significance of the essential oils of 18 Ferula species (Apiaceae) from Iran. Chem. Biodivers. 2011, 8, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Baser, K.H.C.; Kırımer, N. Essential oils of Anatolian Apiaceae-A Profile. Nat. Vol. Essent. Oils 2014, 1, 1–50. Available online: https://nveo.org/wp-content/uploads/2019/03/NVEO-2014-Volume-1-Issue-1-Pages-1-50.pdf (accessed on 20 December 2020).
- Cobb, M.A. The reception and consumption of eastern goods in Roman society. Greece Rome 2013, 60, 136–152. Available online: https://www.jstor.org/stable/43298108 (accessed on 20 December 2020). [CrossRef]
- Godley, A.D. Histories. In Herodotus Complete Works; Delphi Publishing Co., Delphi Classics, Hastings: East Sussex, UK, 2013. [Google Scholar]
- Aydogan, F.; Baykan, S.; Soliman, G.A.; Yusufoglu, H.; Bedir, E. Evaluation of the potential aphrodisiac activity of sesquiterpenoids from Ferula huber-morathii Peşmen in male rats. J. Ethnopharmacol. 2020, 257, 112868. [Google Scholar] [CrossRef]
- Eser, N.; Buyuknacar, H.S.; Cimentepe, O.O.; Gocmen, C.; Ucar, Y.; Erdogan, S.; Yoshiyama, M. The Effect of Ferula elaeochytris root extract on erectile dysfunction in streptozicin-induced diabetic rat. Int. J. Impot. Res. 2020, 32, 186–194. [Google Scholar] [CrossRef]
- Eser, N.; Yoldaş, A.; Yigin, A.; Yumusak, N.; Bozkurt, A.S.; Kokbas, U.; Aslan, M. The protective effect of Ferula elaeochytris on age-related erectile dysfunction. J. Ethnopharmacol. 2020, 258, 112921. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Metabolite | Biological Activities |
|---|---|
| Spathulenol (1) | Immunomodulator [32], anti-nociceptive [64], antimicrobial [65], alleviates cardiac fibrosis [66], antioxidant [33], antiproliferative, antimycobacterial [33], anti-inflammatory [33,67], antitumor [67]. |
| Preisocalamendiol (3) | Positive GABAA receptor modulator [58]. |
| Shyobunone (4) | Positive GABAA receptor modulator [58], insect repellant and insecticide [68]. |
| Isoshyobunone (5) | Positive GABAA receptor modulator [58], insect repellant and insecticide [68]. |
| Acorusnol (8) | Anti-inflammatory [41], germination inhibitor [40]. |
| Teucladiol (11) | Cytotoxic against MCF-7 (estrogen-responsive mammalian adenocarcinoma) [43,69], MDA-MB-435 (estrogen non-responsive mammalian cancer) [43], HCT116 (colon cancer) [69] cell lines. |
| Chrysothol (12) | Cytotoxic against MCF-7 (estrogen-responsive mammalian adenocarcinoma), MDA-MB-435 (estrogen non-responsive mammalian cancer) cell lines [43]. |
| Umbelliferone (14) | Anti-inflammatory [70,71], alleviates liver fibrosis [71], bone loss prevention [72], partial restoration of erectile dysfunction [73], antioxidant [74], urease inhibitor [75], anti-bacterial, anti-fungal, antidiabetic, neuroprotective, anti-cancer, molluscicidal [76]. |
| Scopoletin (15) | Inhibitor of VEGF-induced angiogenesis [77], hepatoprotective [78], prevention of FGF-2-induced angiogenesis [79], antitermite [80], antimicrobial against multi-drug resistant Pseudomonas aeruginosa [81], acaricidal [82], antidopaminergic and antiadrenergic [83], inhibitor of acetylcholinesterase [84], inhibitor of human tumor vascularization [85], antipsychotic [86], ameliorates steatosis and inflammation in diabetic mice [87], antibacterial against multi-drug resistant clinical isolate pathogen strains [88]. |
| Umbelliprenin (16) | Antigenotoxic [89], antioxidant, anti-inflammatory, lipoxygenase inhibitor [90], matrix metalloproteinase inhibitor [91], antitumor [92], cytotoxic activity against CH1 (ovarian), A549 (lung), SK-MEL-28 (melanoma) [93], M4Beu (metastatic pigmented malignant melanoma), QU-DB (large cell lung) [94], and UO31 (renal) [48] cancer cell lines, modulator of melanogenesis [95], antihypertension [96], cancer chemoprevention [97], antiangiogenic [98], antimetastatic and immunostimulatory [99]. |
| Conferone (17) | Urease inhibitor [75], cytotoxic activity against CH1 (ovarian), A549 (lung) and SK-MEL-28 (melanoma) cancer cell lines [93], cancer chemoprevention [97], antiangiogenic [98]. |
| Feselol (18) | Cancer chemoprevention [97], potential aphrodisiac [63]. |
| Conferol (19) | Urease inhibitor [75], modulators of multi-drug resistance in clinical isolates of Escherichia coli and Staphylococcus aureus [100], antileishmanial [101], antiviral against Influenza A (H1N1) virus [102], cytotoxic against HepG2 (hepatocellular carcinoma), Hep3B (hepatocellular carcinoma) and MCF-7 (estrogen-responsive mammalian adenocarcinoma) cancer cell lines [102]. |
| Badrakemone (20) | Cytotoxic against UO31 (renal) cancer cell line [48], cancer chemoprevention [97], weak matrix metalloproteinase inhibitor [91]. |
| Colladonin (21) | Cytotoxic against COLO205 (colon), KM12 (colon), A498 (kidney carcinoma), UO31 (renal), TC32 (Ewing’s sarcoma) [48], HCT116 (human colorectal), HT-29 (human colorectal) [103] cancer cell lines, endocannabinoid system modulator [104]. |
| Badrakemin (22) | Cytotoxic against KM12 (colon), A498 (kidney carcinoma), UO31 (renal) cancer cell lines [48], inhibitor of butyrylcholinesterase [105]. |
| Samarcandin (23) | Potential aphrodisiac [63], antifungal [54], cytotoxic against AGS (human gastric carcinoma), WEHI-164 (fibrosarcoma) cancer cell lines [106], active in NCI yeast anticancer drug screen assays [107], potential antiviral activity against Ebola virus [108,109]. |
| Samarcandin Acetate (26) | Potential aphrodisiac [63]. |
| Myristicin (27) | Cancer chemopreventive agent [51], anti-inflammatory [52], antimicrobial against B. subtilis, E. coli, S. aureus [110], hepatoprotective [111], cytoprotective against hypoxia-induced apoptosis and endoplasmic reticulum stress [112], down-regulates expression of pro-inflammatory cytokines [113], cytotoxic against SK-N-SH (human neuroblastoma) cancer cell line [114], acetylcholinesterase inhibitor [115], antiproliferative [116], insecticidal against Culex pipiens and Aedes aegypti [117]. |
| Laserine (28) | Weak cytotoxicity against HL-60 (acute promyelocytic leukemia) cancer cell line [118], inhibitor of skin photo-aging [119]. |
| Cynaroside (29) (Luteolin-7-β-d-glucoside) | Choleretic and anticholestatic [120], antioxidant and anticholinesterase [121], antibacterial against multi-drug resistant clinical isolate strains [88], anti-inflammatory [122], inhibitor of monoamine oxidase B [123], inhibitor of low-density lipoprotein (LDL) oxidation [124], antimicrobial [125], hepatoprotective [126]. |
| Cynarin (30) | Antimicrobial [125], hepatoprotective [126], antihypertensive, vasodilator [127], choleretic [128]. |
| Chlorogenic Acid (31) | Antimicrobial [125], hepatoprotective [126], antihypertensive, vasodilator [127], antitumor [129], anti-inflammatory [130], improves late diabetes [131], protects against cholestatic liver injury [132], neuroprotective [133], antiviral activity against influenza A (H1N1/H3N2) virus [134], anti-diabetic and anti-lipidemic [135], inhibits hepatocellular carcinoma [136], anxiolytic and antioxidant [137], antihyperalgesic [138], cardioprotective [139], neuroprotective and cognitive improvement [140], improves hepatic steatosis and insulin resistance [141], alleviates obesity and modulates gut microbiota [142], ameliorates ulcerative colitis [143], inhibits glioblastoma growth [144], induces 4T1 breast cancer tumor’s apoptosis [145], strong matrix metalloproteinase-9 inhibitor [146]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miski, M. Next Chapter in the Legend of Silphion: Preliminary Morphological, Chemical, Biological and Pharmacological Evaluations, Initial Conservation Studies, and Reassessment of the Regional Extinction Event. Plants 2021, 10, 102. https://doi.org/10.3390/plants10010102
Miski M. Next Chapter in the Legend of Silphion: Preliminary Morphological, Chemical, Biological and Pharmacological Evaluations, Initial Conservation Studies, and Reassessment of the Regional Extinction Event. Plants. 2021; 10(1):102. https://doi.org/10.3390/plants10010102
Chicago/Turabian StyleMiski, Mahmut. 2021. "Next Chapter in the Legend of Silphion: Preliminary Morphological, Chemical, Biological and Pharmacological Evaluations, Initial Conservation Studies, and Reassessment of the Regional Extinction Event" Plants 10, no. 1: 102. https://doi.org/10.3390/plants10010102
APA StyleMiski, M. (2021). Next Chapter in the Legend of Silphion: Preliminary Morphological, Chemical, Biological and Pharmacological Evaluations, Initial Conservation Studies, and Reassessment of the Regional Extinction Event. Plants, 10(1), 102. https://doi.org/10.3390/plants10010102