
A New Toolbox in Experimental Embryology—Alternative Model Organisms for Studying Preimplantation Development


{kind=link}
{kind=link}
Abstract
1. Introduction
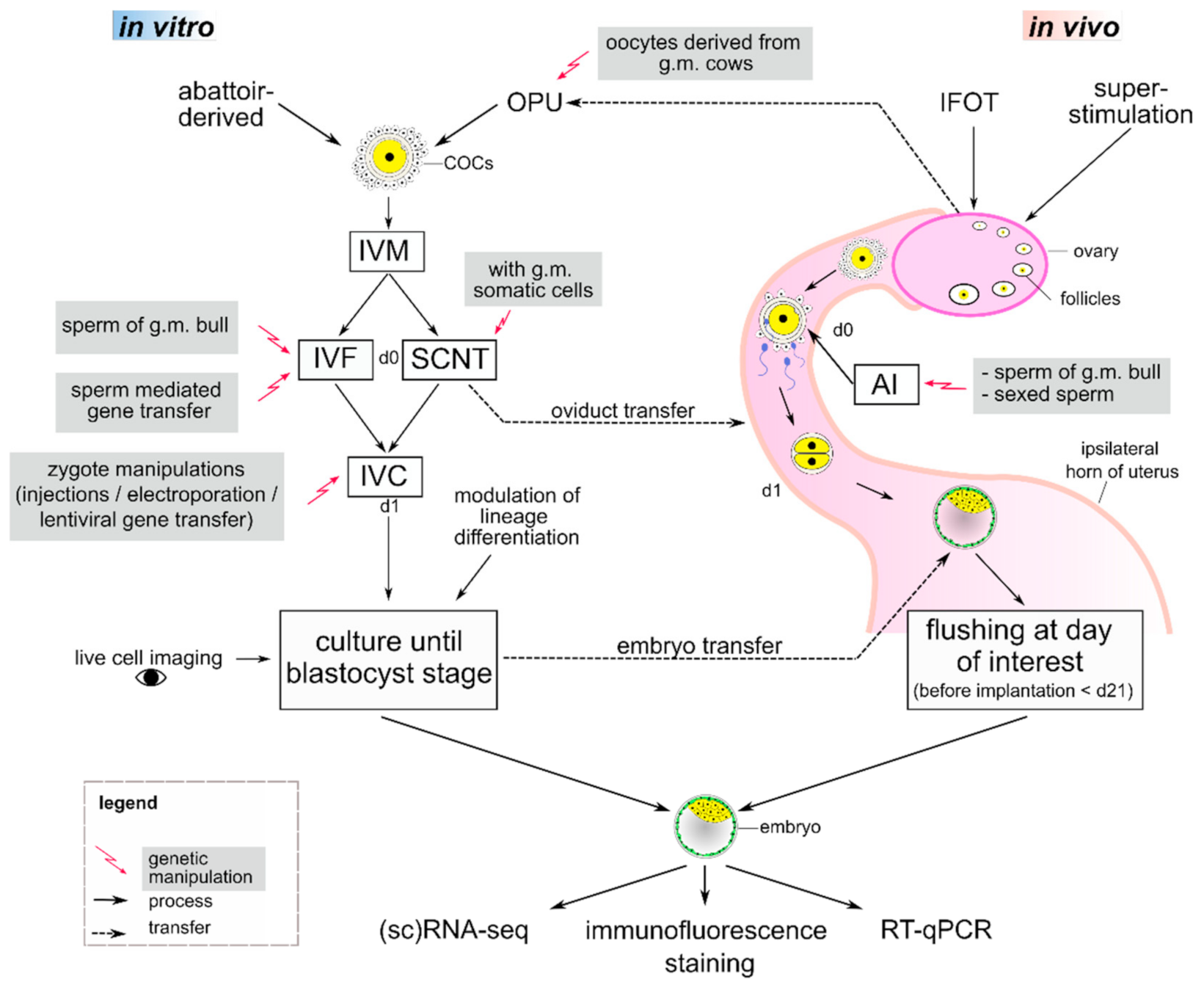
2. State of the ART—Assisted Reproductive Technologies in Cattle and Pigs
2.1. Superstimulation and Embryo Transfer
2.2. Ovum Pick-Up (OPU)
2.3. Intrafollicular Oocyte Transfer (IFOT)
2.4. Intracytoplasmic Sperm Injection (ICSI)
2.5. In Vitro Production (IVP) of Embryos
2.6. Somatic Cell Nuclear Transfer (SCNT)
3. Genetic Manipulations
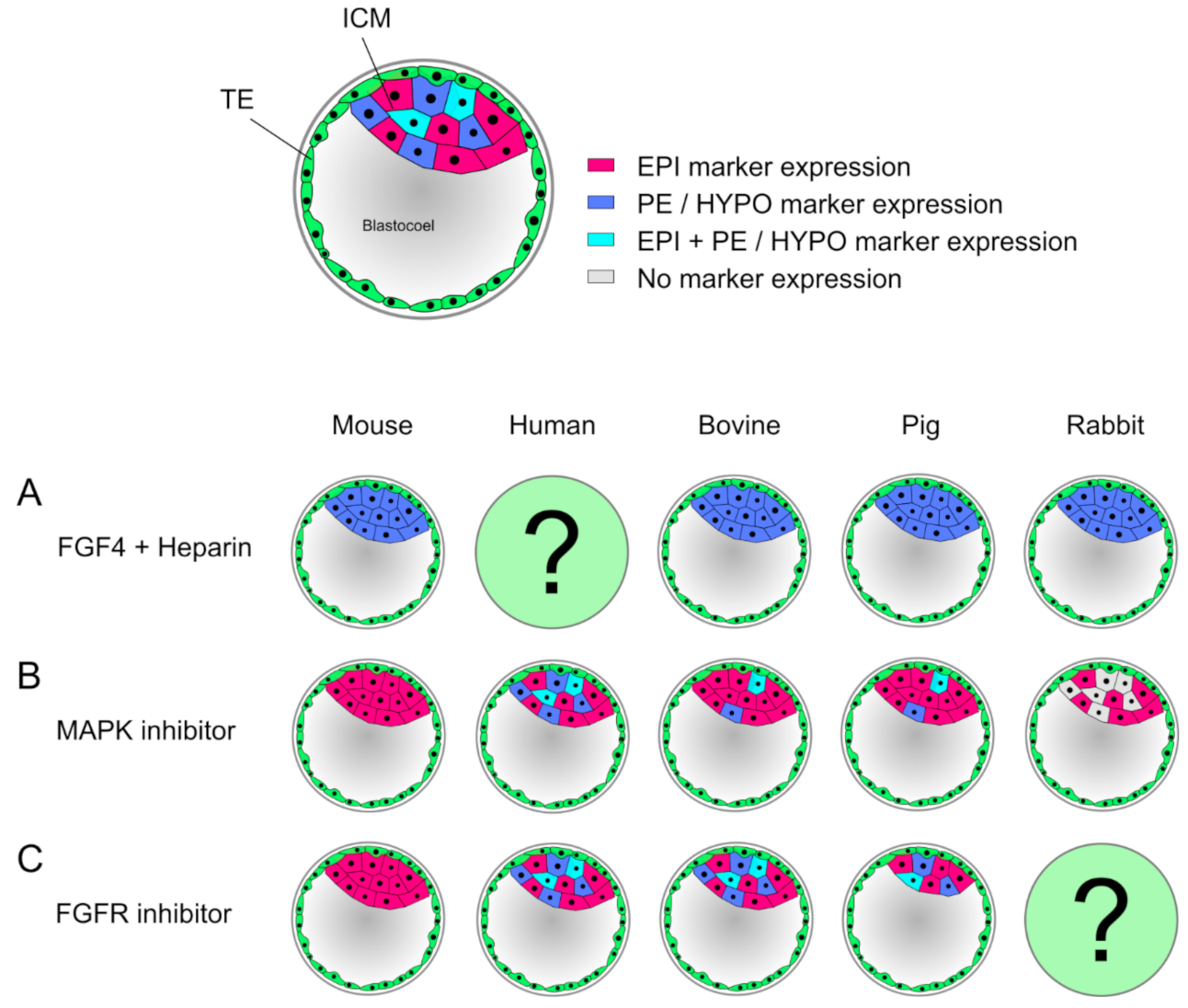
4. New Insights into Preimplantation Development from Alternative Model Organisms
5. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 3D | Three-dimensional |
| AETE | Association of Embryo Technology in Europe |
| AI | Artificial insemination |
| AMH | Anti-Müllerian hormone |
| ART | Assisted reproductive technology |
| BSA | Bovine serum albumin |
| COC | Cumulus-oocyte complex |
| d0 | Day 0 |
| DSB | Double strand break |
| EGF | Epidermal growth factor |
| EGFP | Enhanced green fluorescent protein |
| EPI | Epiblast |
| ESC | Embryonic stem cells |
| ET | Embryo transfer |
| FGFR | FGF-receptor |
| FSH | Follicle-stimulating hormone |
| GE | Genetically engineered |
| HDR | Homology-directed repair |
| HYPO | Hypoblast |
| ICM | Inner cell mass |
| ICSI | Intracytoplasmic sperm injection |
| IFOT | Intrafollicular oocyte transfer |
| IVF | In vitro fertilization |
| IVM | In vitro maturation |
| IVP | In vitro production |
| JAK/STAT | Janus kinase/signal transducer and activator of transcription |
| NHEJ | Non-homologous end joining |
| OPU | Ovum pick-up |
| PE | Primitive endoderm |
| PHD | Post-hatching development |
| RNA-seq | RNA-sequencing |
| SCNT | Somatic cell nuclear transfer |
| scRNA-seq | Single-cell RNA-sequencing |
| ZGA | Zygotic genome activation |
| ZI | Zygote injection |
References
- Gerri, C.; Menchero, S.; Mahadevaiah, S.K.; Turner, J.M.A.; Niakan, K.K. Human Embryogenesis: A Comparative Perspective. Annu. Rev. Cell Dev. Biol. 2020, 36, 411–440. [Google Scholar] [CrossRef] [PubMed]
- Madeja, Z.E.; Pawlak, P.; Piliszek, A. Beyond the mouse: Non-rodent animal models for study of early mammalian development and biomedical research. Int. J. Dev. Biol. 2019, 63, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Piliszek, A.; Grabarek, J.B.; Frankenberg, S.R.; Plusa, B. Cell fate in animal and human blastocysts and the determination of viability. Mol. Hum. Reprod. 2016, 22, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Knaus, W. Dairy cows trapped between performance demands and adaptability. J. Sci. Food Agric. 2009, 89, 1107–1114. [Google Scholar] [CrossRef]
- Koketsu, Y.; Tani, S.; Iida, R. Factors for improving reproductive performance of sows and herd productivity in commercial breeding herds. Porc. Health Manag. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Willett, E.L.; Black, W.G.; Casida, L.E.; Stone, W.H.; Buckner, P.J. Successful transplantation of a fertilized bovine ovum. Science 1951, 113, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, M.C.; Kappen, K.A.; Kruip, T.A.; Taverne, M.A. Aspiration of bovine oocytes during transvaginal ultrasound scanning of the ovaries. Theriogenology 1988, 30, 751–762. [Google Scholar] [CrossRef]
- Hansen, P.J. Implications of Assisted Reproductive Technologies for Pregnancy Outcomes in Mammals. Annu. Rev. Anim. Biosci. 2020, 8, 395–413. [Google Scholar] [CrossRef] [PubMed]
- Mapletoft, R.J.; Hasler, J.F. Assisted reproductive technologies in cattle: A review. Rev. Sci. Tech. 2005, 24, 393–403. [Google Scholar] [CrossRef]
- Greve, T.; Xu, K.P.; Callesen, H.; Hyttel, P. In vivo development of in vitro fertilized bovine oocytes matured in vivo versus in vitro. J. Vitro Fert. Embryo Transf. 1987, 4, 281–285. [Google Scholar] [CrossRef]
- Brackett, B.G.; Bousquet, D.; Boice, M.L.; Donawick, W.J.; Evans, J.F.; Dressel, M.A. Normal development following in vitro fertilization in the cow. Biol. Reprod. 1982, 27, 147–158. [Google Scholar] [CrossRef]
- Rizos, D.; Ward, F.; Duffy, P.; Boland, M.P.; Lonergan, P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: Implications for blastocyst yield and blastocyst quality. Mol. Reprod. Dev. 2002, 61, 234–248. [Google Scholar] [CrossRef]
- Besenfelder, U.; Havlicek, V.; Mosslacher, G.; Brem, G. Collection of tubal stage bovine embryos by means of endoscopy. A technique report. Theriogenology 2001, 55, 837–845. [Google Scholar] [CrossRef]
- Putney, D.J.; Thatcher, W.W.; Drost, M.; Wright, J.M.; DeLorenzo, M.A. Influence of environmental temperature on reproductive performance of bovine embryo donors and recipients in the southwest region of the United States. Theriogenology 1988, 30, 905–922. [Google Scholar] [CrossRef]
- Ferraz, P.A.; Burnley, C.; Karanja, J.; Viera-Neto, A.; Santos, J.E.; Chebel, R.C.; Galvao, K.N. Factors affecting the success of a large embryo transfer program in Holstein cattle in a commercial herd in the southeast region of the United States. Theriogenology 2016, 86, 1834–1841. [Google Scholar] [CrossRef]
- Quinton, H. Commercial Embryo Transfer Activity in Europe. 2019. Available online: https://www.aete.eu/publications/statistics/ (accessed on 15 January 2021).
- Rico, C.; Fabre, S.; Médigue, C.; Clemente, N.D.; Clément, F.; Bontoux, M.; Touzé, J.-L.; Dupont, M.; Briant, E.; Rémy, B. Anti-Müllerian hormone is an endocrine marker of ovarian gonadotropin-responsive follicles and can help to predict superovulatory responses in the cow. Biol. Reprod. 2009, 80, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.Y.; Schmidt, K.T.; Kristensen, S.G.; Rosendahl, M.; Byskov, A.G.; Ernst, E. Concentrations of AMH and inhibin-B in relation to follicular diameter in normal human small antral follicles. Hum. Reprod. 2010, 25, 1282–1287. [Google Scholar] [CrossRef]
- Sartori, R.; Bastos, M.R.; Wiltbank, M.C. Factors affecting fertilisation and early embryo quality in single- and superovulated dairy cattle. Reprod. Fertil. Dev. 2010, 22, 151–158. [Google Scholar] [CrossRef]
- Gad, A.; Besenfelder, U.; Rings, F.; Ghanem, N.; Salilew-Wondim, D.; Hossain, M.M.; Tesfaye, D.; Lonergan, P.; Becker, A.; Cinar, U.; et al. Effect of reproductive tract environment following controlled ovarian hyperstimulation treatment on embryo development and global transcriptome profile of blastocysts: Implications for animal breeding and human assisted reproduction. Hum. Reprod. 2011, 26, 1693–1707. [Google Scholar] [CrossRef]
- Angel, M.A.; Gil, M.A.; Cuello, C.; Sanchez-Osorio, J.; Gomis, J.; Parrilla, I.; Vila, J.; Colina, I.; Diaz, M.; Reixach, J.; et al. The effects of superovulation of donor sows on ovarian response and embryo development after nonsurgical deep-uterine embryo transfer. Theriogenology 2014, 81, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Caárdenas, H.; Pope, W. Control of ovulation rate in swine. J. Anim. Sci. 2002, 80, E36–E46. [Google Scholar] [CrossRef]
- Besenfelder, U.; Brem, G. Tubal transfer of bovine embryos: A simple endoscopic method reducing long-term exposure of in vitro produced embryos. Theriogenology 1998, 50, 739–745. [Google Scholar] [CrossRef]
- Graf, A.; Krebs, S.; Zakhartchenko, V.; Schwalb, B.; Blum, H.; Wolf, E. Fine mapping of genome activation in bovine embryos by RNA sequencing. Proc. Natl. Acad. Sci. USA 2014, 111, 4139–4144. [Google Scholar] [CrossRef]
- Dean, W.; Santos, F.; Stojkovic, M.; Zakhartchenko, V.; Walter, J.; Wolf, E.; Reik, W. Conservation of methylation reprogramming in mammalian development: Aberrant reprogramming in cloned embryos. Proc. Natl. Acad. Sci. USA 2001, 98, 13734–13738. [Google Scholar] [CrossRef]
- Graf, A.; Krebs, S.; Heininen-Brown, M.; Zakhartchenko, V.; Blum, H.; Wolf, E. Genome activation in bovine embryos: Review of the literature and new insights from RNA sequencing experiments. Anim. Reprod. Sci. 2014, 149, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, M.; Rho, H.S.; Hemerich, D.; Henning, H.H.W.; van Tol, H.T.A.; Holker, M.; Besenfelder, U.; Mokry, M.; Vos, P.; Stout, T.A.E.; et al. An oviduct-on-a-chip provides an enhanced in vitro environment for zygote genome reprogramming. Nat. Commun. 2018, 9, 4934–4948. [Google Scholar] [CrossRef]
- Moore, N.W.; Shelton, J.N. Egg Transfer in Sheep. Effect of Degree of Synchronization between Donor and Recipient, Age of Egg, and Site of Transfer on the Survival of Transferred Eggs. J. Reprod. Fertil. 1964, 7, 145–152. [Google Scholar] [CrossRef]
- Jones, G.M.; Trounson, A.O.; Lolatgis, N.; Wood, C. Factors affecting the success of human blastocyst development and pregnancy following in vitro fertilization and embryo transfer. Fertil. Steril. 1998, 70, 1022–1029. [Google Scholar] [CrossRef]
- Kurome, M.; Kessler, B.; Wuensch, A.; Nagashima, H.; Wolf, E. Nuclear transfer and transgenesis in the pig. In Nuclear Reprogramming; Springer: Berlin/Heidelberg, Germany, 2015; pp. 37–59. [Google Scholar] [CrossRef]
- Wolf, E.; Braun-Reichhart, C.; Streckel, E.; Renner, S. Genetically engineered pig models for diabetes research. Transgenic Res. 2014, 23, 27–38. [Google Scholar] [CrossRef]
- Martinez, E.A.; Caamano, J.N.; Gil, M.A.; Rieke, A.; McCauley, T.C.; Cantley, T.C.; Vazquez, J.M.; Roca, J.; Vazquez, J.L.; Didion, B.A.; et al. Successful nonsurgical deep uterine embryo transfer in pigs. Theriogenology 2004, 61, 137–146. [Google Scholar] [CrossRef]
- Reichenbach, H.D.; Modl, J.; Brem, G. Piglets Born after Transcervical Transfer of Embryos into Recipient Gilts. Vet. Rec. 1993, 133, 36–39. [Google Scholar] [CrossRef]
- Galli, C.; Crotti, G.; Notari, C.; Turini, P.; Duchi, R.; Lazzari, G. Embryo production by ovum pick up from live donors. Theriogenology 2001, 55, 1341–1357. [Google Scholar] [CrossRef]
- Monteiro, F.M.; Batista, E.O.S.; Vieira, L.M.; Bayeux, B.M.; Accorsi, M.; Campanholi, S.P.; Dias, E.A.R.; Souza, A.H.; Baruselli, P.S. Beef donor cows with high number of retrieved COC produce more in vitro embryos compared with cows with low number of COC after repeated ovum pick-up sessions. Theriogenology 2017, 90, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Galli, C.; Duchi, R.; Colleoni, S.; Lagutina, I.; Lazzari, G. Ovum pick up, intracytoplasmic sperm injection and somatic cell nuclear transfer in cattle, buffalo and horses: From the research laboratory to clinical practice. Theriogenology 2014, 81, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Kassens, A.; Held, E.; Salilew-Wondim, D.; Sieme, H.; Wrenzycki, C.; Tesfaye, D.; Schellander, K.; Hoelker, M. Intrafollicular Oocyte Transfer (IFOT) of Abattoir-Derived and In Vitro-Matured Oocytes Results in Viable Blastocysts and Birth of Healthy Calves. Biol. Reprod. 2015, 92, 150–164. [Google Scholar] [CrossRef]
- Hoelker, M.; Kassens, A.; Salilew-Wondim, D.; Sieme, H.; Wrenzycki, C.; Tesfaye, D.; Neuhoff, C.; Schellander, K.; Held-Hoelker, E. Birth of healthy calves after intra-follicular transfer (IFOT) of slaughterhouse derived immature bovine oocytes. Theriogenology 2017, 97, 41–49. [Google Scholar] [CrossRef]
- Spricigo, J.F.; Sena Netto, S.B.; Muterlle, C.V.; Rodrigues Sde, A.; Leme, L.O.; Guimaraes, A.L.; Caixeta, F.M.; Franco, M.M.; Pivato, I.; Dode, M.A. Intrafollicular transfer of fresh and vitrified immature bovine oocytes. Theriogenology 2016, 86, 2054–2062. [Google Scholar] [CrossRef]
- Garcia-Rosello, E.; Garcia-Mengual, E.; Coy, P.; Alfonso, J.; Silvestre, M.A. Intracytoplasmic sperm injection in livestock species: An update. Reprod. Domest. Anim. 2009, 44, 143–151. [Google Scholar] [CrossRef]
- Martin, M.J. Development of in vivo-matured porcine oocytes following intracytoplasmic sperm injection. Biol. Reprod. 2000, 63, 109–112. [Google Scholar] [CrossRef]
- Goto, K.; Kinoshita, A.; Takuma, Y.; Ogawa, K. Fertilisation of bovine oocytes by the injection of immobilised, killed spermatozoa. Vet. Rec. 1990, 127, 517–520. [Google Scholar]
- Kolbe, T.; Holtz, W. Birth of a piglet derived from an oocyte fertilized by intracytoplasmic sperm injection (ICSI). Anim. Reprod. Sci. 2000, 64, 97–101. [Google Scholar] [CrossRef]
- Nakai, M.; Kashiwazaski, N.; Takizawa, A.; Hayashi, Y.; Nakatsukasa, E.; Fuchimoto, D.; Noguchi, J.; Kaneko, H.; Shino, M.; Kikuchi, K. Viable piglets generated from porcine oocytes matured in vitro and fertilized by intracytoplasmic sperm head injection. Biol. Reprod. 2003, 68, 1003–1008. [Google Scholar] [CrossRef]
- Probst, S.; Rath, D. Production of piglets using intracytoplasmic sperm injection (ICSI) with flowcytometrically sorted boar semen and artificially activated oocytes. Theriogenology 2003, 59, 961–973. [Google Scholar] [CrossRef]
- Lai, L.; Sun, Q.; Wu, G.; Murphy, C.N.; Kuhholzer, B.; Park, K.W.; Bonk, A.J.; Day, B.N.; Prather, R.S. Development of porcine embryos and offspring after intracytoplasmic sperm injection with liposome transfected or non-transfected sperm into in vitro matured oocytes. Zygote 2001, 9, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Leemans, B.; Stout, T.A.E.; De Schauwer, C.; Heras, S.; Nelis, H.; Hoogewijs, M.; Van Soom, A.; Gadella, B.M. Update on mammalian sperm capacitation: How much does the horse differ from other species? Reproduction 2019, 157, 181–197. [Google Scholar] [CrossRef]
- Gomez, E.; Rodriguez, A.; Munoz, M.; Caamano, J.N.; Hidalgo, C.O.; Moran, E.; Facal, N.; Diez, C. Serum free embryo culture medium improves in vitro survival of bovine blastocysts to vitrification. Theriogenology 2008, 69, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Adan, A.; Lonergan, P.; Rizos, D.; Ward, F.A.; Boland, M.P.; Pintado, B.; de la Fuente, J. Effect of the in vitro culture system on the kinetics of blastocyst development and sex ratio of bovine embryos. Theriogenology 2001, 55, 1117–1126. [Google Scholar] [CrossRef]
- Gordon, I. Laboratory Production of Cattle Embryos; CABI: Wallingford, UK, 2003; Volume 27. [Google Scholar]
- Wrenzycki, C. In vitro culture systems: How far are we from optimal conditions? Anim. Reprod. 2018, 13, 279–282. [Google Scholar] [CrossRef]
- Parrish, J.J. Bovine in vitro fertilization: In vitro oocyte maturation and sperm capacitation with heparin. Theriogenology 2014, 81, 67–73. [Google Scholar] [CrossRef]
- Lonergan, P.; Fair, T. In vitro-produced bovine embryos: Dealing with the warts. Theriogenology 2008, 69, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Brandao, D.O.; Maddox-Hyttel, P.; Lovendahl, P.; Rumpf, R.; Stringfellow, D.; Callesen, H. Post hatching development: A novel system for extended in vitro culture of bovine embryos. Biol. Reprod. 2004, 71, 2048–2055. [Google Scholar] [CrossRef]
- Vajta, G.; Alexopoulos, N.I.; Callesen, H. Rapid growth and elongation of bovine blastocysts in vitro in a three-dimensional gel system. Theriogenology 2004, 62, 1253–1263. [Google Scholar] [CrossRef]
- Machado, G.M.; Ferreira, A.R.; Pivato, I.; Fidelis, A.; Spricigo, J.F.; Paulini, F.; Lucci, C.M.; Franco, M.M.; Dode, M.A. Post-hatching development of in vitro bovine embryos from day 7 to 14 in vivo versus in vitro. Mol. Reprod. Dev. 2013, 80, 936–947. [Google Scholar] [CrossRef]
- Alexopoulos, N.I.; Vajta, G.; Maddox-Hyttel, P.; French, A.J.; Trounson, A.O. Stereomicroscopic and histological examination of bovine embryos following extended in vitro culture. Reprod. Fertil. Dev. 2005, 17, 799–808. [Google Scholar] [CrossRef]
- Ramos-Ibeas, P.; Lamas-Toranzo, I.; Martinez-Moro, A.; de Frutos, C.; Quiroga, A.C.; Zurita, E.; Bermejo-Alvarez, P. Embryonic disc formation following post-hatching bovine embryo development in vitro. Reproduction 2020, 160, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Whyte, J.J.; Meyer, A.E.; Spate, L.D.; Benne, J.A.; Cecil, R.; Samuel, M.S.; Murphy, C.N.; Prather, R.S.; Geisert, R.D. Inactivation of porcine interleukin-1beta results in failure of rapid conceptus elongation. Proc. Natl. Acad. Sci. USA 2018, 115, 307–312. [Google Scholar] [CrossRef]
- West, R.C.; Ming, H.; Logsdon, D.M.; Sun, J.; Rajput, S.K.; Kile, R.A.; Schoolcraft, W.B.; Roberts, R.M.; Krisher, R.L.; Jiang, Z.; et al. Dynamics of trophoblast differentiation in peri-implantation-stage human embryos. Proc. Natl. Acad. Sci. USA 2019, 116, 22635–22644. [Google Scholar] [CrossRef] [PubMed]
- Simintiras, C.A.; Sánchez, J.M.; McDonald, M.; Lonergan, P. The biochemistry surrounding bovine conceptus elongation. Biol. Reprod. 2019, 101, 328–337. [Google Scholar] [CrossRef]
- Blomberg, L.; Hashizume, K.; Viebahn, C. Blastocyst elongation, trophoblastic differentiation, and embryonic pattern formation. Reproduction 2008, 135, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Mor, A.; Reddy, I.; Nandi, S.; Gupta, P.; Mishra, A. In Vitro Embryo Production in Sheep. In Comparative Embryo Culture; Springer: Berlin/Heidelberg, Germany, 2019; pp. 131–140. [Google Scholar]
- Souza-Fabjan, J.M.G.; Batista, R.I.T.P.; Correia, L.F.L.; Paramio, M.T.; Fonseca, J.F.; Freitas, V.J.F.; Mermillod, P. In vitro production of small ruminant embryos: Latest improvements and further research. Reprod. Fertil. Dev. 2021, 33, 31–54. [Google Scholar] [CrossRef]
- Diskin, M.G.; Murphy, J.J.; Sreenan, J.M. Embryo survival in dairy cows managed under pastoral conditions. Anim. Reprod. Sci. 2006, 96, 297–311. [Google Scholar] [CrossRef]
- Crosier, A.E.; Farin, P.W.; Dykstra, M.J.; Alexander, J.E.; Farin, C.E. Ultrastructural morphometry of bovine blastocysts produced in vivo or in vitro. Biol. Reprod. 2001, 64, 1375–1385. [Google Scholar] [CrossRef]
- Fair, T.; Lonergan, P.; Dinnyes, A.; Cottell, D.C.; Hyttel, P.; Ward, F.A.; Boland, M.P. Ultrastructure of bovine blastocysts following cryopreservation: Effect of method of blastocyst production. Mol. Reprod. Dev. 2001, 58, 186–195. [Google Scholar] [CrossRef]
- Rizos, D.; Gutierrez-Adan, A.; Perez-Garnelo, S.; De La Fuente, J.; Boland, M.P.; Lonergan, P. Bovine embryo culture in the presence or absence of serum: Implications for blastocyst development, cryotolerance, and messenger RNA expression. Biol. Reprod. 2003, 68, 236–243. [Google Scholar] [CrossRef]
- Lazzari, G.; Wrenzycki, C.; Herrmann, D.; Duchi, R.; Kruip, T.; Niemann, H.; Galli, C. Cellular and molecular deviations in bovine in vitro-produced embryos are related to the large offspring syndrome. Biol. Reprod. 2002, 67, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.L.; Everts, R.E.; Sung, L.Y.; Du, F.; Page, R.L.; Henderson, B.; Rodriguez-Zas, S.L.; Nedambale, T.L.; Renard, J.P.; Lewin, H.A.; et al. Gene expression profiling of single bovine embryos uncovers significant effects of in vitro maturation, fertilization and culture. Mol. Reprod. Dev. 2009, 76, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, D.; Ponsuksili, S.; Wimmers, K.; Gilles, M.; Schellander, K. A comparative expression analysis of gene transcripts in post-fertilization developmental stages of bovine embryos produced in vitro or in vivo. Reprod. Domest. Anim. 2004, 39, 396–404. [Google Scholar] [CrossRef]
- Niemann, H.; Wrenzycki, C. Alterations of expression of developmentally important genes in preimplantation bovine embryos by in vitro culture conditions: Implications for subsequent development. Theriogenology 2000, 53, 21–34. [Google Scholar] [CrossRef]
- Mucci, N.; Aller, J.; Kaiser, G.G.; Hozbor, F.; Cabodevila, J.; Alberio, R.H. Effect of estrous cow serum during bovine embryo culture on blastocyst development and cryotolerance after slow freezing or vitrification. Theriogenology 2006, 65, 1551–1562. [Google Scholar] [CrossRef]
- Long, C.R.; Dobrinsky, J.R.; Johnson, L.A. In vitro production of pig embryos: Comparisons of culture media and boars. Theriogenology 1999, 51, 1375–1390. [Google Scholar] [CrossRef]
- Romar, R.; Canovas, S.; Matas, C.; Gadea, J.; Coy, P. Pig in vitro fertilization: Where are we and where do we go? Theriogenology 2019, 137, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Coy, P.; Romar, R. In vitro production of pig embryos: A point of view. Reprod. Fertil. Dev. 2002, 14, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Sherrer, E.S.; Rathbun, T.J.; Davis, D.L. Fertilization and blastocyst development in oocytes obtained from prepubertal and adult pigs. J. Anim. Sci. 2004, 82, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Machaty, Z.; Day, B.N.; Prather, R.S. Development of early porcine embryos in vitro and in vivo. Biol. Reprod. 1998, 59, 451–455. [Google Scholar] [CrossRef]
- Yoshioka, K.; Suzuki, C.; Tanaka, A.; Anas, I.M.; Iwamura, S. Birth of piglets derived from porcine zygotes cultured in a chemically defined medium. Biol. Reprod. 2002, 66, 112–119. [Google Scholar] [CrossRef]
- Miles, J.R.; Laughlin, T.D.; Sargus-Patino, C.N.; Pannier, A.K. In vitro porcine blastocyst development in three-dimensional alginate hydrogels. Mol. Reprod. Dev. 2017, 84, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T. The improvement of in vitro maturation systems for bovine and porcine oocytes. Theriogenology 2001, 55, 1291–1301. [Google Scholar] [CrossRef]
- Koo, D.B.; Kim, Y.J.; Yu, I.; Kim, H.N.; Lee, K.K.; Han, Y.M. Effects of in vitro fertilization conditions on preimplantation development and quality of pig embryos. Anim. Reprod. Sci. 2005, 90, 101–110. [Google Scholar] [CrossRef]
- Herrick, J.R.; Conover-Sparman, M.L.; Krisher, R.L. Reduced polyspermic fertilization of porcine oocytes utilizing elevated bicarbonate and reduced calcium concentrations in a single-medium system. Reprod. Fertil. Dev. 2003, 15, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Abeydeera, L.R.; Day, B.N. Fertilization and subsequent development in vitro of pig oocytes inseminated in a modified tris-buffered medium with frozen-thawed ejaculated spermatozoa. Biol. Reprod. 1997, 57, 729–734. [Google Scholar] [CrossRef]
- Steinborn, R.; Schinogl, P.; Zakhartchenko, V.; Achmann, R.; Schernthaner, W.; Stojkovic, M.; Wolf, E.; Muller, M.; Brem, G. Mitochondrial DNA heteroplasmy in cloned cattle produced by fetal and adult cell cloning. Nat. Genet. 2000, 25, 255–257. [Google Scholar] [CrossRef] [PubMed]
- Keefer, C.L. Artificial cloning of domestic animals. Proc. Natl. Acad. Sci. USA 2015, 112, 8874–8878. [Google Scholar] [CrossRef]
- Qu, P.; Wang, Y.; Zhang, C.; Liu, E. Insights into the roles of sperm in animal cloning. Stem Cell Res. Ther. 2020, 11, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Wells, K.D.; Prather, R.S. Genome-editing technologies to improve research, reproduction, and production in pigs. Mol. Reprod. Dev. 2017, 84, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Uh, K.; Farrell, K. Current progress of genome editing in livestock. Theriogenology 2020, 150, 229–235. [Google Scholar] [CrossRef]
- Tsunoda, Y.; Kato, Y.; Inui, A. Cloning in Cattle; Taylor & Francis: London, UK, 2006. [Google Scholar]
- Zhang, Y.; Li, J.; Villemoes, K.; Pedersen, A.M.; Purup, S.; Vajta, G. An epigenetic modifier results in improved in vitro blastocyst production after somatic cell nuclear transfer. Cloning Stem Cells 2007, 9, 357–363. [Google Scholar] [CrossRef]
- Holker, M.; Petersen, B.; Hassel, P.; Kues, W.A.; Lemme, E.; Lucas-Hahn, A.; Niemann, H. Duration of in vitro maturation of recipient oocytes affects blastocyst development of cloned porcine embryos. Cloning Stem Cells 2005, 7, 35–44. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, G.S.; Kim, J.H.; Kang, S.K.; Lee, B.C.; Hwang, W.S. Expression of leptin ligand and receptor and effect of exogenous leptin supplement on in vitro development of porcine embryos. Theriogenology 2006, 65, 831–844. [Google Scholar] [CrossRef]
- Lagutina, I.; Lazzari, G.; Duchi, R.; Turini, P.; Tessaro, I.; Brunetti, D.; Colleoni, S.; Crotti, G.; Galli, C. Comparative aspects of somatic cell nuclear transfer with conventional and zona-free method in cattle, horse, pig and sheep. Theriogenology 2007, 67, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Kurome, M.; Geistlinger, L.; Kessler, B.; Zakhartchenko, V.; Klymiuk, N.; Wuensch, A.; Richter, A.; Baehr, A.; Kraehe, K.; Burkhardt, K.; et al. Factors influencing the efficiency of generating genetically engineered pigs by nuclear transfer: Multi-factorial analysis of a large data set. BMC Biotechnol. 2013, 13, 43–56. [Google Scholar] [CrossRef]
- Long, C.R.; Westhusin, M.E.; Golding, M.C. Reshaping the transcriptional frontier: Epigenetics and somatic cell nuclear transfer. Mol. Reprod. Dev. 2014, 81, 183–193. [Google Scholar] [CrossRef]
- Simmet, K.; Wolf, E.; Zakhartchenko, V. Manipulating the Epigenome in Nuclear Transfer Cloning: Where, When and How. Int. J. Mol. Sci. 2021, 22, 236. [Google Scholar] [CrossRef] [PubMed]
- Bauersachs, S.; Ulbrich, S.E.; Zakhartchenko, V.; Minten, M.; Reichenbach, M.; Reichenbach, H.D.; Blum, H.; Spencer, T.E.; Wolf, E. The endometrium responds differently to cloned versus fertilized embryos. Proc. Natl. Acad. Sci. USA 2009, 106, 5681–5686. [Google Scholar] [CrossRef]
- Mansouri-Attia, N.; Sandra, O.; Aubert, J.; Degrelle, S.; Everts, R.E.; Giraud-Delville, C.; Heyman, Y.; Galio, L.; Hue, I.; Yang, X.; et al. Endometrium as an early sensor of in vitro embryo manipulation technologies. Proc. Natl. Acad. Sci. USA 2009, 106, 5687–5692. [Google Scholar] [CrossRef] [PubMed]
- Young, L.E.; Sinclair, K.D.; Wilmut, I. Large offspring syndrome in cattle and sheep. Rev. Reprod. 1998, 3, 155–163. [Google Scholar] [CrossRef]
- Archer, G.S.; Dindot, S.; Friend, T.H.; Walker, S.; Zaunbrecher, G.; Lawhorn, B.; Piedrahita, J.A. Hierarchical phenotypic and epigenetic variation in cloned swine. Biol. Reprod. 2003, 69, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Han, J.; Wu, J.; Li, Q.; Liu, S.; Zhang, W.; Pei, Y.; Ruan, X.; Liu, Z.; Wang, X.; et al. Specific gene-regulation networks during the pre-implantation development of the pig embryo as revealed by deep sequencing. BMC Genom. 2014, 15, 4–16. [Google Scholar] [CrossRef]
- Laible, G.; Wei, J.W.; Wagner, S. Improving livestock for agriculture—Technological progress from random transgenesis to precision genome editing heralds a new era. Biotechnol. J. 2015, 10, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Ratner, L.D.; La Motta, G.E.; Briski, O.; Salamone, D.F.; Fernandez-Martin, R. Practical approaches for knock-out gene editing in pigs. Front. Genet. 2020, 11, 1847. [Google Scholar] [CrossRef]
- Georges, M.; Charlier, C.; Hayes, B. Harnessing genomic information for livestock improvement. Nat. Rev. Genet. 2019, 20, 135–156. [Google Scholar] [CrossRef]
- Tan, W.; Proudfoot, C.; Lillico, S.G.; Whitelaw, C.B. Gene targeting, genome editing: From Dolly to editors. Transgenic Res. 2016, 25, 273–287. [Google Scholar] [CrossRef]
- Vochozkova, P.; Simmet, K.; Jemiller, E.M.; Wunsch, A.; Klymiuk, N. Gene Editing in Primary Cells of Cattle and Pig. Methods Mol. Biol. 2019, 1961, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Simmet, K.; Zakhartchenko, V.; Wolf, E. Comparative aspects of early lineage specification events in mammalian embryos—Insights from reverse genetics studies. Cell Cycle 2018, 17, 1688–1695. [Google Scholar] [CrossRef]
- Perisse, I.V.; Fan, Z.; Singina, G.N.; White, K.L.; Polejaeva, I.A. Improvements in Gene Editing Technology Boost Its Applications in Livestock. Front. Genet. 2020, 11, 614688–614709. [Google Scholar] [CrossRef] [PubMed]
- Simmet, K.; Zakhartchenko, V.; Philippou-Massier, J.; Blum, H.; Klymiuk, N.; Wolf, E. OCT4/POU5F1 is required for NANOG expression in bovine blastocysts. Proc. Natl. Acad. Sci. USA 2018, 115, 2770–2775. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.S.; Hyun, S.H.; Kim, H.S.; Kim, D.Y.; Lee, S.H.; Lim, J.M.; Lee, E.S.; Kang, S.K.; Lee, B.C.; Hwang, W.S. Improvement of a porcine somatic cell nuclear transfer technique by optimizing donor cell and recipient oocyte preparations. Theriogenology 2003, 59, 1949–1957. [Google Scholar] [CrossRef]
- Guo, Y.; Li, H.; Wang, Y.; Yan, X.; Sheng, X.; Chang, D.; Qi, X.; Wang, X.; Liu, Y.; Li, J.; et al. Screening somatic cell nuclear transfer parameters for generation of transgenic cloned cattle with intragenomic integration of additional gene copies that encode bovine adipocyte-type fatty acid-binding protein (A-FABP). Mol. Biol. Rep. 2017, 44, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.A.; Tanihara, F.; Wittayarat, M.; Namula, Z.; Sato, Y.; Lin, Q.; Takebayashi, K.; Hirata, M.; Otoi, T. Comparison of the effects of introducing the CRISPR/Cas9 system by microinjection and electroporation into porcine embryos at different stages. BMC Res. Notes 2021, 14, 1–7. [Google Scholar] [CrossRef]
- Tanihara, F.; Takemoto, T.; Kitagawa, E.; Rao, S.; Do, L.T.; Onishi, A.; Yamashita, Y.; Kosugi, C.; Suzuki, H.; Sembon, S.; et al. Somatic cell reprogramming-free generation of genetically modified pigs. Sci. Adv. 2016, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Camargo, L.S.A.; Owen, J.R.; Van Eenennaam, A.L.; Ross, P.J. Efficient One-Step Knockout by Electroporation of Ribonucleoproteins into Zona-Intact Bovine Embryos. Front. Genet. 2020, 11, 570069. [Google Scholar] [CrossRef]
- Bevacqua, R.J.; Fernandez-Martin, R.; Savy, V.; Canel, N.G.; Gismondi, M.I.; Kues, W.A.; Carlson, D.F.; Fahrenkrug, S.C.; Niemann, H.; Taboga, O.A.; et al. Efficient edition of the bovine PRNP prion gene in somatic cells and IVF embryos using the CRISPR/Cas9 system. Theriogenology 2016, 86, 1886–1896. [Google Scholar] [CrossRef] [PubMed]
- Crispo, M.; Mulet, A.P.; Tesson, L.; Barrera, N.; Cuadro, F.; dos Santos-Neto, P.C.; Nguyen, T.H.; Creneguy, A.; Brusselle, L.; Anegon, I.; et al. Efficient Generation of Myostatin Knock-Out Sheep Using CRISPR/Cas9 Technology and Microinjection into Zygotes. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef]
- Liang, P.; Xu, Y.; Zhang, X.; Ding, C.; Huang, R.; Zhang, Z.; Lv, J.; Xie, X.; Chen, Y.; Li, Y.; et al. CRISPR/Cas9-mediated gene editing in human tripronuclear zygotes. Protein Cell 2015, 6, 363–372. [Google Scholar] [CrossRef]
- Yen, S.T.; Zhang, M.; Deng, J.M.; Usman, S.J.; Smith, C.N.; Parker-Thornburg, J.; Swinton, P.G.; Martin, J.F.; Behringer, R.R. Somatic mosaicism and allele complexity induced by CRISPR/Cas9 RNA injections in mouse zygotes. Dev. Biol. 2014, 393, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.R.; Hennig, S.L.; McNabb, B.R.; Mansour, T.A.; Smith, J.M.; Lin, J.C.; Young, A.E.; Trott, J.F.; Murray, J.D.; Delany, M.E.; et al. One-step generation of a targeted knock-in calf using the CRISPR-Cas9 system in bovine zygotes. BMC Genom. 2021, 22, 118–129. [Google Scholar] [CrossRef]
- Ma, H.; Marti-Gutierrez, N.; Park, S.W.; Wu, J.; Lee, Y.; Suzuki, K.; Koski, A.; Ji, D.; Hayama, T.; Ahmed, R.; et al. Correction of a pathogenic gene mutation in human embryos. Nature 2017, 548, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, J.; Berg, D.K.; Pfeffer, P.L. Morphological and Gene Expression Changes in Cattle Embryos from Hatched Blastocyst to Early Gastrulation Stages after Transfer of In Vitro Produced Embryos. PLoS ONE 2015, 10, 1–26. [Google Scholar] [CrossRef]
- Kurome, M.; Ueda, H.; Tomii, R.; Naruse, K.; Nagashima, H. Production of transgenic-clone pigs by the combination of ICSI-mediated gene transfer with somatic cell nuclear transfer. Transgenic Res. 2006, 15, 229–240. [Google Scholar] [CrossRef]
- Umeyama, K.; Saito, H.; Kurome, M.; Matsunari, H.; Watanabe, M.; Nakauchi, H.; Nagashima, H. Characterization of the ICSI-mediated gene transfer method in the production of transgenic pigs. Mol. Reprod. Dev. 2012, 79, 218–228. [Google Scholar] [CrossRef]
- Watanabe, M.; Kurome, M.; Matsunari, H.; Nakano, K.; Umeyema, K.; Shiota, A.; Nakauchi, H.; Nagashima, H. The creation of transgenic pigs expressing human proteins using BAC-derived, full-length genes and intracytoplasmic sperm injection-mediated gene transfer. Transgenic Res. 2012, 21, 605–618. [Google Scholar] [CrossRef]
- Arias, M.E.; Sanchez-Villalba, E.; Delgado, A.; Felmer, R. Effect of transfection and co-incubation of bovine sperm with exogenous DNA on sperm quality and functional parameters for its use in sperm-mediated gene transfer. Zygote 2017, 25, 85–97. [Google Scholar] [CrossRef]
- Canovas, S.; Gutierrez-Adan, A.; Gadea, J. Effect of exogenous DNA on bovine sperm functionality using the sperm mediated gene transfer (SMGT) technique. Mol. Reprod. Dev. 2010, 77, 687–698. [Google Scholar] [CrossRef]
- Lavitrano, M.; Forni, M.; Bacci, M.L.; Di Stefano, C.; Varzi, V.; Wang, H.; Seren, E. Sperm mediated gene transfer in pig: Selection of donor boars and optimization of DNA uptake. Mol. Reprod. Dev. 2003, 64, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Schellander, K.; Peli, J.; Schmoll, F.; Brem, G. Artificial insemination in cattle with DNA-treated sperm. Anim. Biotechnol. 1995, 6, 41–50. [Google Scholar] [CrossRef]
- Smith, K.; Spadafora, C. Sperm-mediated gene transfer: Applications and implications. Bioessays 2005, 27, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.; Kessler, B.; Ewerling, S.; Weppert, M.; Vogg, B.; Ludwig, H.; Stojkovic, M.; Boelhauve, M.; Brem, G.; Wolf, E.; et al. Efficient transgenesis in farm animals by lentiviral vectors. EMBO Rep. 2003, 4, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.; Zakhartchenko, V.; Weppert, M.; Sebald, H.; Wenigerkind, H.; Brem, G.; Wolf, E.; Pfeifer, A. Generation of transgenic cattle by lentiviral gene transfer into oocytes. Biol. Reprod. 2004, 71, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.; Kessler, B.; Ewerling, S.; Kabermann, A.; Brem, G.; Wolf, E.; Pfeifer, A. Epigenetic regulation of lentiviral transgene vectors in a large animal model. Mol. Ther. 2006, 13, 59–66. [Google Scholar] [CrossRef]
- Reichenbach, M.; Lim, T.; Reichenbach, H.D.; Guengoer, T.; Habermann, F.A.; Matthiesen, M.; Hofmann, A.; Weber, F.; Zerbe, H.; Grupp, T.; et al. Germ-line transmission of lentiviral PGK-EGFP integrants in transgenic cattle: New perspectives for experimental embryology. Transgenic Res. 2010, 19, 549–556. [Google Scholar] [CrossRef]
- Whitelaw, C.B.; Lillico, S.G.; King, T. Production of transgenic farm animals by viral vector-mediated gene transfer. Reprod. Domest. Anim. 2008, 43 (Suppl. S2), 355–358. [Google Scholar] [CrossRef]
- Messerschmidt, D.M.; Kemler, R. Nanog is required for primitive endoderm formation through a non-cell autonomous mechanism. Dev. Biol. 2010, 344, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, S.; Gerbe, F.; Bessonnard, S.; Belville, C.; Pouchin, P.; Bardot, O.; Chazaud, C. Primitive endoderm differentiates via a three-step mechanism involving Nanog and RTK signaling. Dev. Cell 2011, 21, 1005–1013. [Google Scholar] [CrossRef]
- Cai, K.Q.; Capo-Chichi, C.D.; Rula, M.E.; Yang, D.H.; Xu, X.X. Dynamic GATA6 expression in primitive endoderm formation and maturation in early mouse embryogenesis. Dev. Dyn. 2008, 237, 2820–2829. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.A.; Teo, R.T.; Li, H.; Robson, P.; Glover, D.M.; Zernicka-Goetz, M. Origin and formation of the first two distinct cell types of the inner cell mass in the mouse embryo. Proc. Natl. Acad. Sci. USA 2010, 107, 6364–6369. [Google Scholar] [CrossRef] [PubMed]
- Plusa, B.; Piliszek, A.; Frankenberg, S.; Artus, J.; Hadjantonakis, A.K. Distinct sequential cell behaviours direct primitive endoderm formation in the mouse blastocyst. Development 2008, 135, 3081–3091. [Google Scholar] [CrossRef]
- Mitsui, K.; Tokuzawa, Y.; Itoh, H.; Segawa, K.; Murakami, M.; Takahashi, K.; Maruyama, M.; Maeda, M.; Yamanaka, S. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 2003, 113, 631–642. [Google Scholar] [CrossRef]
- Silva, J.; Nichols, J.; Theunissen, T.W.; Guo, G.; van Oosten, A.L.; Barrandon, O.; Wray, J.; Yamanaka, S.; Chambers, I.; Smith, A. Nanog is the gateway to the pluripotent ground state. Cell 2009, 138, 722–737. [Google Scholar] [CrossRef]
- Piliszek, A.; Madeja, Z.E. Pre-implantation Development of Domestic Animals. Curr. Top. Dev. Biol. 2018, 128, 267–294. [Google Scholar] [CrossRef]
- Nishioka, N.; Inoue, K.; Adachi, K.; Kiyonari, H.; Ota, M.; Ralston, A.; Yabuta, N.; Hirahara, S.; Stephenson, R.O.; Ogonuki, N.; et al. The Hippo signaling pathway components Lats and Yap pattern Tead4 activity to distinguish mouse trophectoderm from inner cell mass. Dev. Cell 2009, 16, 398–410. [Google Scholar] [CrossRef]
- Sasaki, H. Roles and regulations of Hippo signaling during preimplantation mouse development. Dev. Growth Differ. 2017, 59, 12–20. [Google Scholar] [CrossRef]
- Chazaud, C.; Yamanaka, Y. Lineage specification in the mouse preimplantation embryo. Development 2016, 143, 1063–1074. [Google Scholar] [CrossRef]
- Niakan, K.K.; Eggan, K. Analysis of human embryos from zygote to blastocyst reveals distinct gene expression patterns relative to the mouse. Dev. Biol. 2013, 375, 54–64. [Google Scholar] [CrossRef]
- Blakeley, P.; Fogarty, N.M.; del Valle, I.; Wamaitha, S.E.; Hu, T.X.; Elder, K.; Snell, P.; Christie, L.; Robson, P.; Niakan, K.K. Defining the three cell lineages of the human blastocyst by single-cell RNA-seq. Development 2015, 142, 3151–3165. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.; Edsgard, D.; Reinius, B.; Deng, Q.; Panula, S.P.; Codeluppi, S.; Plaza Reyes, A.; Linnarsson, S.; Sandberg, R.; Lanner, F. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell 2016, 165, 1012–1026. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, N.M.E.; McCarthy, A.; Snijders, K.E.; Powell, B.E.; Kubikova, N.; Blakeley, P.; Lea, R.; Elder, K.; Wamaitha, S.E.; Kim, D.; et al. Genome editing reveals a role for OCT4 in human embryogenesis. Nature 2017, 550, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.K.; Smith, C.S.; Pearton, D.J.; Wells, D.N.; Broadhurst, R.; Donnison, M.; Pfeffer, P.L. Trophectoderm lineage determination in cattle. Dev. Cell 2011, 20, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Gerri, C.; McCarthy, A.; Alanis-Lobato, G.; Demtschenko, A.; Bruneau, A.; Loubersac, S.; Fogarty, N.M.E.; Hampshire, D.; Elder, K.; Snell, P.; et al. Initiation of a conserved trophectoderm program in human, cow and mouse embryos. Nature 2020, 587, 443–447. [Google Scholar] [CrossRef]
- Wicklow, E.; Blij, S.; Frum, T.; Hirate, Y.; Lang, R.A.; Sasaki, H.; Ralston, A. HIPPO pathway members restrict SOX2 to the inner cell mass where it promotes ICM fates in the mouse blastocyst. PLoS Genet. 2014, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Sasaki, H. Epiblast Formation by TEAD-YAP-Dependent Expression of Pluripotency Factors and Competitive Elimination of Unspecified Cells. Dev. Cell 2019, 50, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Chazaud, C.; Yamanaka, Y.; Pawson, T.; Rossant, J. Early lineage segregation between epiblast and primitive endoderm in mouse blastocysts through the Grb2-MAPK pathway. Dev. Cell 2006, 10, 615–624. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Lanner, F.; Rossant, J. FGF signal-dependent segregation of primitive endoderm and epiblast in the mouse blastocyst. Development 2010, 137, 715–724. [Google Scholar] [CrossRef]
- Kuijk, E.W.; van Tol, L.T.; Van de Velde, H.; Wubbolts, R.; Welling, M.; Geijsen, N.; Roelen, B.A. The roles of FGF and MAP kinase signaling in the segregation of the epiblast and hypoblast cell lineages in bovine and human embryos. Development 2012, 139, 871–882. [Google Scholar] [CrossRef]
- Canizo, J.R.; Ynsaurralde Rivolta, A.E.; Vazquez Echegaray, C.; Suva, M.; Alberio, V.; Aller, J.F.; Guberman, A.S.; Salamone, D.F.; Alberio, R.H.; Alberio, R. A dose-dependent response to MEK inhibition determines hypoblast fate in bovine embryos. BMC Dev. Biol. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Warzych, E.; Pawlak, P.; Lechniak, D.; Madeja, Z.E. WNT signalling supported by MEK/ERK inhibition is essential to maintain pluripotency in bovine preimplantation embryo. Dev. Biol. 2020, 463, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Roode, M.; Blair, K.; Snell, P.; Elder, K.; Marchant, S.; Smith, A.; Nichols, J. Human hypoblast formation is not dependent on FGF signalling. Dev. Biol. 2012, 361, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Ibeas, P.; Sang, F.; Zhu, Q.; Tang, W.W.C.; Withey, S.; Klisch, D.; Wood, L.; Loose, M.; Surani, M.A.; Alberio, R. Pluripotency and X chromosome dynamics revealed in pig pre-gastrulating embryos by single cell analysis. Nat. Commun. 2019, 10, 500–517. [Google Scholar] [CrossRef]
- Rodriguez, A.; Allegrucci, C.; Alberio, R. Modulation of pluripotency in the porcine embryo and iPS cells. PLoS ONE 2012, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Piliszek, A.; Madeja, Z.E.; Plusa, B. Suppression of ERK signalling abolishes primitive endoderm formation but does not promote pluripotency in rabbit embryo. Development 2017, 144, 3719–3730. [Google Scholar] [CrossRef]
- Soszynska, A.; Klimczewska, K.; Suwinska, A. FGF/ERK signaling pathway: How it operates in mammalian preimplantation embryos and embryo-derived stem cells. Int. J. Dev. Biol. 2019, 63, 171–186. [Google Scholar] [CrossRef]
- Nichols, J.; Silva, J.; Roode, M.; Smith, A. Suppression of Erk signalling promotes ground state pluripotency in the mouse embryo. Development 2009, 136, 3215–3222. [Google Scholar] [CrossRef]
- McLean, Z.; Meng, F.; Henderson, H.; Turner, P.; Oback, B. Increased MAP kinase inhibition enhances epiblast-specific gene expression in bovine blastocysts. Biol. Reprod. 2014, 91, 49–59. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Deathridge, J.; Antolovic, V.; Parsons, M.; Chubb, J.R. Live imaging of ERK signalling dynamics in differentiating mouse embryonic stem cells. Development 2019, 146, 1–12. [Google Scholar] [CrossRef]
- Pokrass, M.J.; Ryan, K.A.; Xin, T.; Pielstick, B.; Timp, W.; Greco, V.; Regot, S. Cell-Cycle-Dependent ERK Signaling Dynamics Direct Fate Specification in the Mammalian Preimplantation Embryo. Dev. Cell 2020, 55, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.S.; Rahman, S.; Raina, D.; Schroter, C.; Hadjantonakis, A.K. Live Visualization of ERK Activity in the Mouse Blastocyst Reveals Lineage-Specific Signaling Dynamics. Dev. Cell 2020, 55, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Forrester-Gauntlett, B.; Turner, P.; Henderson, H.; Oback, B. Signal inhibition reveals JAK/STAT3 pathway as critical for bovine inner cell mass development. Biol. Reprod. 2015, 93, 132. [Google Scholar] [CrossRef]
- Alberio, R. Regulation of Cell Fate Decisions in Early Mammalian Embryos. Annu. Rev. Anim. Biosci. 2020, 8, 377–393. [Google Scholar] [CrossRef]
- Frum, T.; Halbisen, M.A.; Wang, C.; Amiri, H.; Robson, P.; Ralston, A. Oct4 cell-autonomously promotes primitive endoderm development in the mouse blastocyst. Dev. Cell 2013, 25, 610–622. [Google Scholar] [CrossRef]
- Le Bin, G.C.; Munoz-Descalzo, S.; Kurowski, A.; Leitch, H.; Lou, X.; Mansfield, W.; Etienne-Dumeau, C.; Grabole, N.; Mulas, C.; Niwa, H.; et al. Oct4 is required for lineage priming in the developing inner cell mass of the mouse blastocyst. Development 2014, 141, 1001–1010. [Google Scholar] [CrossRef]
- Stamatiadis, P.; Boel, A.; Cosemans, G.; Popovic, M.; Bekaert, B.; Guggilla, R.; Tang, M.; De Sutter, P.; Van Nieuwerburgh, F.; Menten, B.; et al. Comparative analysis of mouse and human preimplantation development following POU5F1 CRISPR/Cas9 targeting reveals interspecies differences. Hum. Reprod. 2021, 1–11. [Google Scholar] [CrossRef]
- Lee, M.; Oh, J.-N.; Kim, S.-H.; Choi, K.-H.; Lee, D.-K.; Choe, G.C.; Jeong, J.; Lee, C.-K. Identification of the Role of SOX2 During Early Embryogenesis in Pigs. Res. Square 2020. [Google Scholar] [CrossRef]
- Avilion, A.A.; Nicolis, S.K.; Pevny, L.H.; Perez, L.; Vivian, N.; Lovell-Badge, R. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev. 2003, 17, 126–140. [Google Scholar] [CrossRef]
- Pan, H.; Schultz, R.M. Sox2 modulates reprogramming of gene expression in two-cell mouse embryos. Biol. Reprod. 2011, 85, 409–416. [Google Scholar] [CrossRef]
- Adachi, K.; Suemori, H.; Yasuda, S.Y.; Nakatsuji, N.; Kawase, E. Role of SOX2 in maintaining pluripotency of human embryonic stem cells. Genes Cells 2010, 15, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Lavagi, I.; Krebs, S.; Simmet, K.; Beck, A.; Zakhartchenko, V.; Wolf, E.; Blum, H. Single-cell RNA sequencing reveals developmental heterogeneity of blastomeres during major genome activation in bovine embryos. Sci. Rep. 2018, 8, 4071–4083. [Google Scholar] [CrossRef] [PubMed]
- Boroviak, T.; Stirparo, G.G.; Dietmann, S.; Hernando-Herraez, I.; Mohammed, H.; Reik, W.; Smith, A.; Sasaki, E.; Nichols, J.; Bertone, P. Single cell transcriptome analysis of human, marmoset and mouse embryos reveals common and divergent features of preimplantation development. Development 2018, 145, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Stirparo, G.G.; Boroviak, T.; Guo, G.; Nichols, J.; Smith, A.; Bertone, P. Integrated analysis of single-cell embryo data yields a unified transcriptome signature for the human pre-implantation epiblast. Development 2018, 145, 1–14. [Google Scholar] [CrossRef]
- Yao, T.; Suzuki, R.; Furuta, N.; Suzuki, Y.; Kabe, K.; Tokoro, M.; Sugawara, A.; Yajima, A.; Nagasawa, T.; Matoba, S.; et al. Live-cell imaging of nuclear-chromosomal dynamics in bovine in vitro fertilised embryos. Sci. Rep. 2018, 8, 7460–7469. [Google Scholar] [CrossRef]
- Brooks, K.E.; Daughtry, B.L.; Davis, B.; Yan, M.Y.; Fei, S.S.; Carbone, L.; Chavez, S.L. Molecular Contribution to Embryonic Aneuploidy and Genotypic Complexity During Initial Cleavage Divisions of Mammalian Development. bioRxiv 2020. [Google Scholar] [CrossRef]
- Plachta, N.; Bollenbach, T.; Pease, S.; Fraser, S.E.; Pantazis, P. Oct4 kinetics predict cell lineage patterning in the early mammalian embryo. Nat. Cell Biol. 2011, 13, 117–123. [Google Scholar] [CrossRef] [PubMed]
- White, M.D.; Angiolini, J.F.; Alvarez, Y.D.; Kaur, G.; Zhao, Z.W.; Mocskos, E.; Bruno, L.; Bissiere, S.; Levi, V.; Plachta, N. Long-Lived Binding of Sox2 to DNA Predicts Cell Fate in the Four-Cell Mouse Embryo. Cell 2016, 165, 75–87. [Google Scholar] [CrossRef]
- White, M.D.; Bissiere, S.; Alvarez, Y.D.; Plachta, N. Mouse Embryo Compaction. Curr. Top. Dev. Biol. 2016, 120, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Bissiere, S.; Gasnier, M.; Alvarez, Y.D.; Plachta, N. Cell Fate Decisions During Preimplantation Mammalian Development. Curr. Top. Dev. Biol. 2018, 128, 37–58. [Google Scholar] [CrossRef]
- Pergament, E.; Fiddler, M.; Cho, N.; Johnson, D.; Holmgren, W. Fertilization and early embryology: Sexual differentiation and preimplantation cell growth. Hum. Reprod. 1994, 9, 1730–1732. [Google Scholar] [CrossRef]
- Valdivia, R.P.; Kunieda, T.; Azuma, S.; Toyoda, Y. PCR sexing and developmental rate differences in preimplantation mouse embryos fertilized and cultured in vitro. Mol. Reprod. Dev. 1993, 35, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Adán, A.; Behboodi, E.; Andersen, G.; Medrano, J.; Murray, J. Relationship between stage of development and sex of bovine IVM-IVF embryos cultured in vitro versus in the sheep oviduct. Theriogenology 1996, 46, 515–525. [Google Scholar] [CrossRef]
- Avery, B.; Jorgensen, C.B.; Madison, V.; Greve, T. Morphological development and sex of bovine in vitro-fertilized embryos. Mol. Reprod. Dev. 1992, 32, 265–270. [Google Scholar] [CrossRef]
- Cassar, G.; King, W.; King, G. Influence of sex on early growth of pig conceptuses. Reproduction 1994, 101, 317–320. [Google Scholar] [CrossRef]
- Bernardi, M.L.; Delouis, C. Sex-related differences in the developmental rate of in-vitro matured/in-vitro fertilized ovine embryos. Hum. Reprod. 1996, 11, 621–626. [Google Scholar] [CrossRef]
- Bermejo-Alvarez, P.; Rizos, D.; Rath, D.; Lonergan, P.; Gutierrez-Adan, A. Epigenetic differences between male and female bovine blastocysts produced in vitro. Physiol. Genom. 2008, 32, 264–272. [Google Scholar] [CrossRef]
- Kobayashi, S.; Isotani, A.; Mise, N.; Yamamoto, M.; Fujihara, Y.; Kaseda, K.; Nakanishi, T.; Ikawa, M.; Hamada, H.; Abe, K. Comparison of gene expression in male and female mouse blastocysts revealed imprinting of the X-linked gene, Rhox5/Pem, at preimplantation stages. Curr. Biol. 2006, 16, 166–172. [Google Scholar] [CrossRef]
- Okamoto, I.; Patrat, C.; Thépot, D.; Peynot, N.; Fauque, P.; Daniel, N.; Diabangouaya, P.; Wolf, J.-P.; Renard, J.-P.; Duranthon, V. Eutherian mammals use diverse strategies to initiate X-chromosome inactivation during development. Nature 2011, 472, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; van Tol, H.T.A.; Stout, T.A.E.; Roelen, B.A.J. Initiation of X Chromosome Inactivation during Bovine Embryo Development. Cells 2020, 9, 1016. [Google Scholar] [CrossRef] [PubMed]
- Frum, T.; Ralston, A. Culture conditions antagonize lineage-promoting signaling in the mouse blastocyst. Reproduction 2020, 160, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, J.P.; Schroder, J.; Aberkane, A.; Ouyang, J.F.; Mohenska, M.; Lim, S.M.; Sun, Y.B.Y.; Chen, J.; Sun, G.; et al. Modelling human blastocysts by reprogramming fibroblasts into iBlastoids. Nature 2021, 591, 627–632. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Springer, C.; Wolf, E.; Simmet, K. A New Toolbox in Experimental Embryology—Alternative Model Organisms for Studying Preimplantation Development. J. Dev. Biol. 2021, 9, 15. https://doi.org/10.3390/jdb9020015
Springer C, Wolf E, Simmet K. A New Toolbox in Experimental Embryology—Alternative Model Organisms for Studying Preimplantation Development. Journal of Developmental Biology. 2021; 9(2):15. https://doi.org/10.3390/jdb9020015
Chicago/Turabian StyleSpringer, Claudia, Eckhard Wolf, and Kilian Simmet. 2021. "A New Toolbox in Experimental Embryology—Alternative Model Organisms for Studying Preimplantation Development" Journal of Developmental Biology 9, no. 2: 15. https://doi.org/10.3390/jdb9020015
APA StyleSpringer, C., Wolf, E., & Simmet, K. (2021). A New Toolbox in Experimental Embryology—Alternative Model Organisms for Studying Preimplantation Development. Journal of Developmental Biology, 9(2), 15. https://doi.org/10.3390/jdb9020015