Study of Natural Longlife Juvenility and Tissue Regeneration in Caudate Amphibians and Potential Application of Resulting Data in Biomedicine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Immunity, Genetic Features, and Cell Plasticity Associated with Regenerative Abilities in Urodela Amphibians
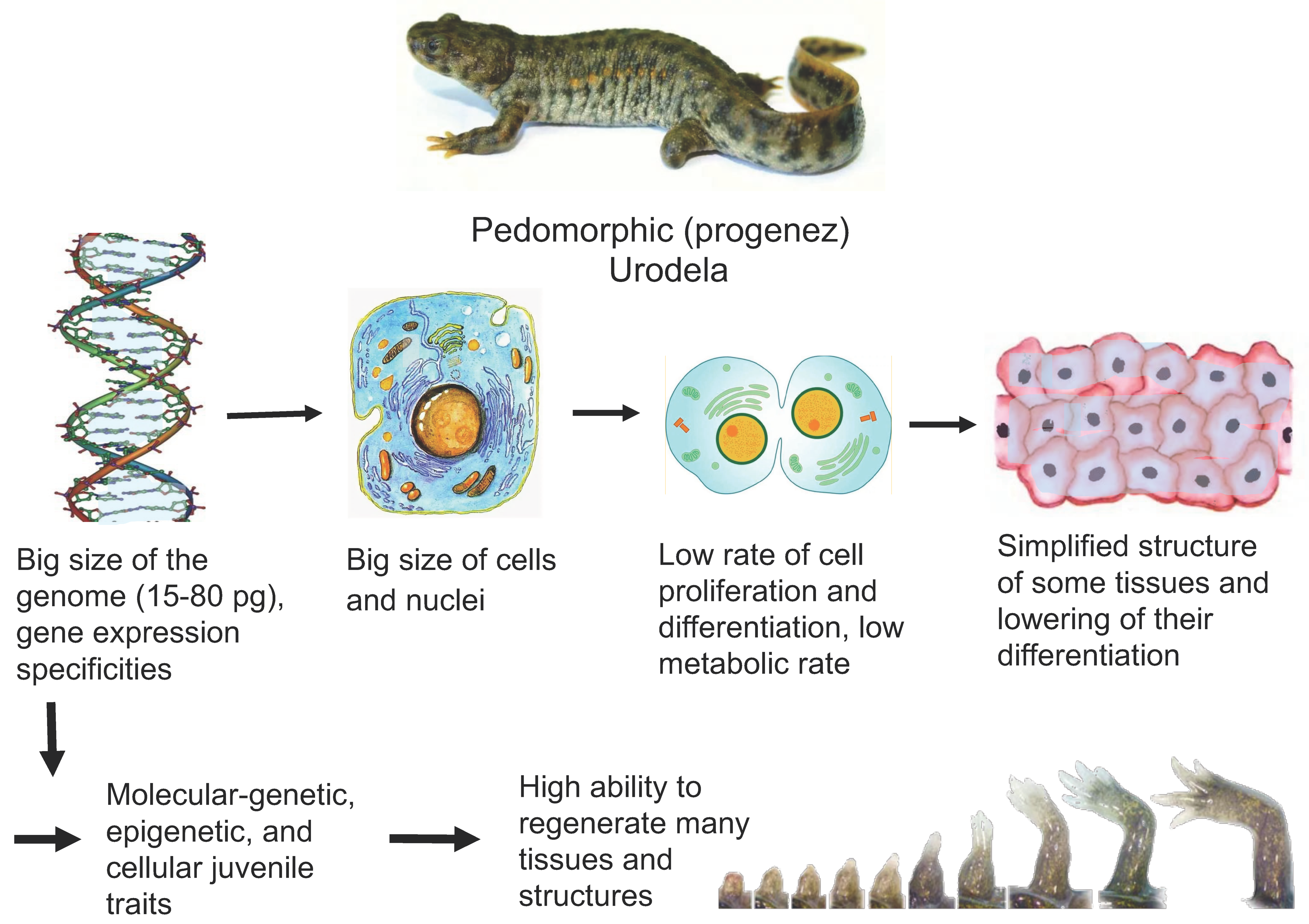
3. Pedomorphosis, Genome, and Cell Sizes in Urodela
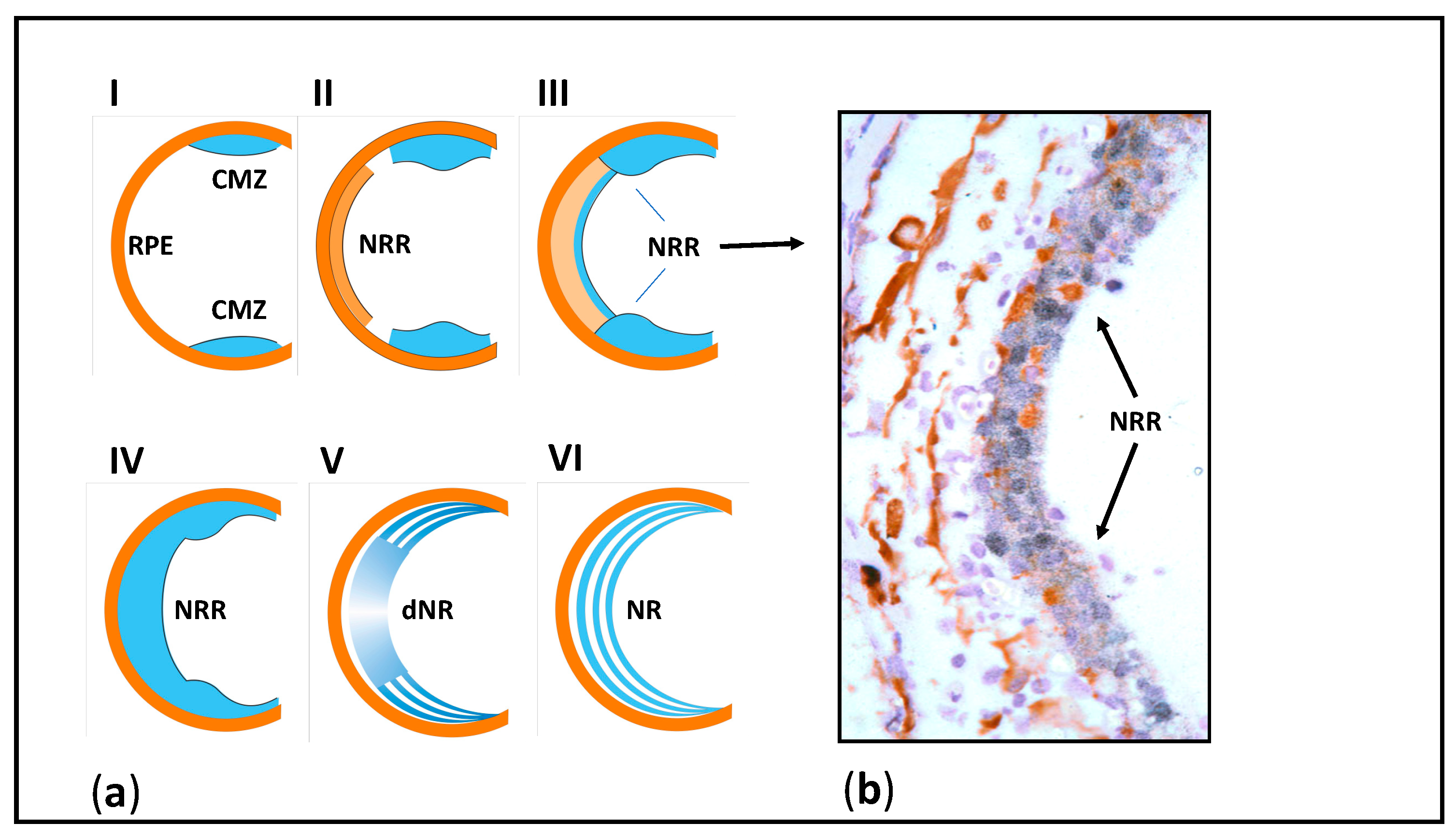
4. Specifics of Eye and Brain Tissues and Their Regeneration in Urodela
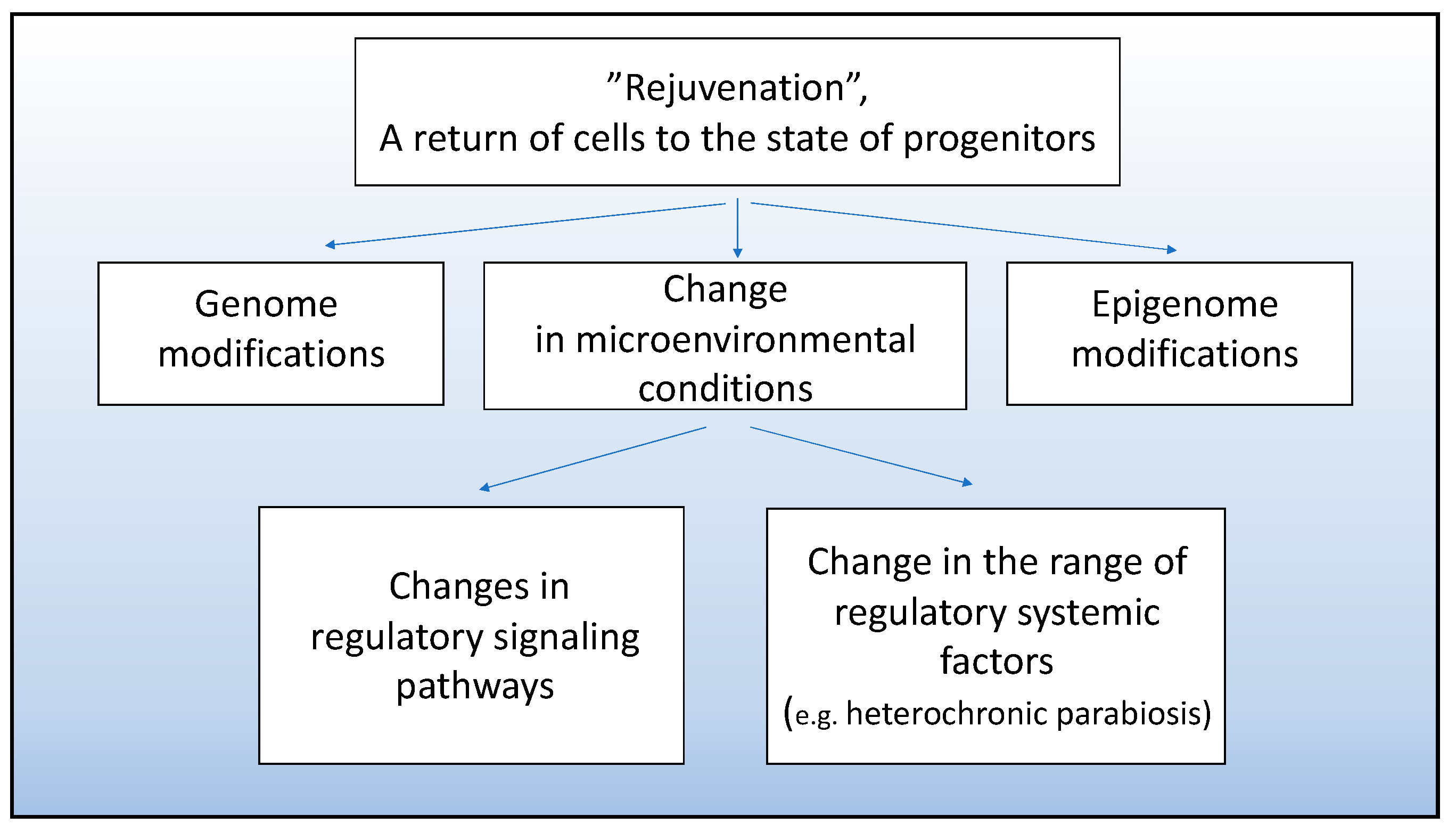
5. Attempts at Experimental Cell “Rejuvenation” to Stimulate Involvement of Mammalian Cells in Tissue Regeneration
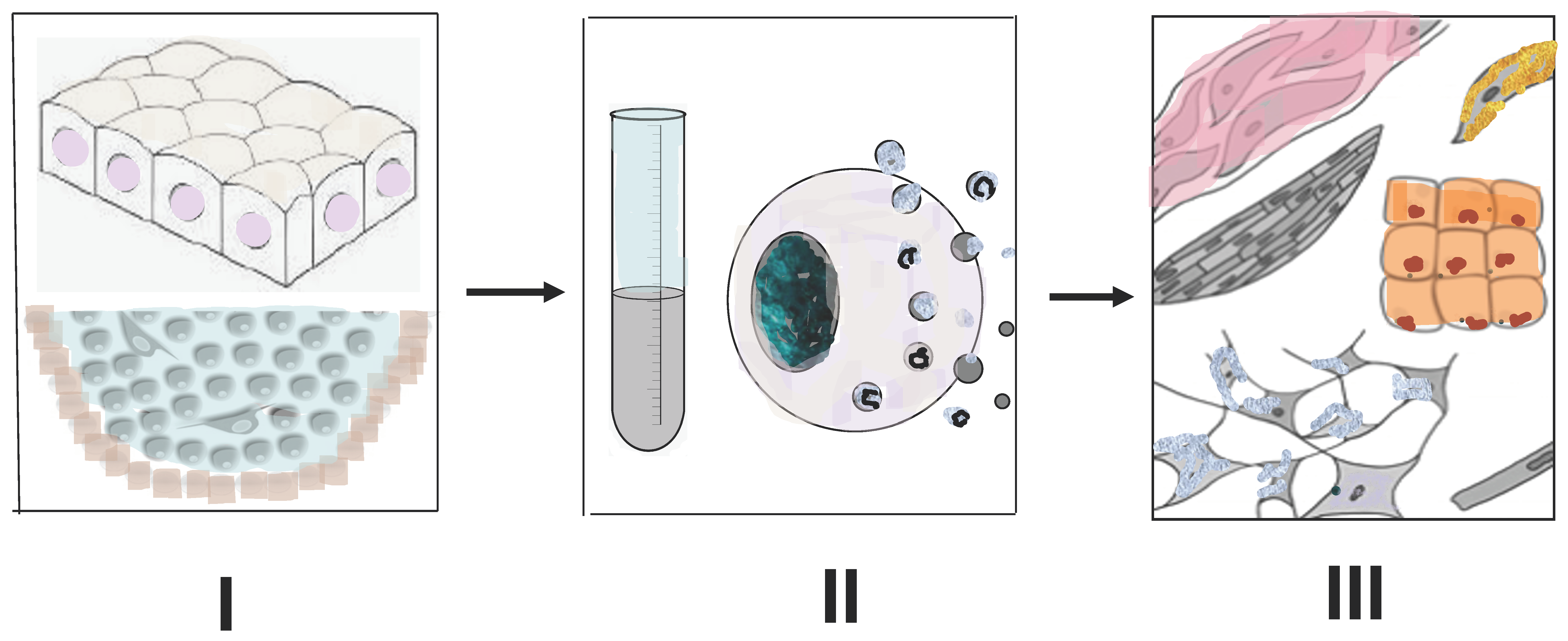
6. Tissues of Animals with High Regenerative Capacities as a Source of Compounds for Stimulating Regeneration in Higher Vertebrates
7. Conclusions
Funding
Conflicts of Interest
References
- Del Rio-Tsonis, K.; Tsonis, P.A. Eye regeneration at the molecular age. Dev. Dyn. 2003, 226, 211–224. [Google Scholar] [CrossRef]
- Sa´nchez Alvarado, A.; Tsonis, P.A. Bridging the regeneration gap: Genetic insights from diverse animal models. Nat. Rev. Genet. 2006, 7, 873–884. [Google Scholar] [CrossRef]
- Mastellos, D.C.; DeAngelis, R.A.; Lambris, J.D. Complement-triggered pathways orchestrate regenerative responses throughout phylogenesis. Semin Immunol. 2013, 25, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Bolanos Castro, L.A.; Walters, H.E.; Vazquez, R.O.G.; Yun, M.H. Immunity in salamander regeneration: Where are we standing and where are we headed? Dev. Dynam. 2020, 1–15. [Google Scholar] [CrossRef]
- Mescher, A.L.; Neff, A.W. Regenerative capacity and the developing immune system. Adv. Biochem. Eng. Biotechnol. 2005, 93, 39–66. [Google Scholar] [PubMed]
- Mescher, A.L.; Neff, A.W.; King, M.W. Inflammation and immunity in organ regeneration. Dev. Comput. Immunol. 2017, 66, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Flajnik, M.F.; Miler, K.; Du Pasquier, L. Evolution of the immune system. In Fundamental Immunology, 5th ed.; Paul, W.E., Ed.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2003; pp. 519–570. [Google Scholar]
- Godwin, J.W.; Rosenthal, N. Scar-free wound healing and regeneration in amphibians: Immunological influences on regenerative success. Differentiation 2014, 87, 66–75. [Google Scholar] [CrossRef]
- Alibardi, L. Organ regeneration evolved in fish and amphibians in relation to metamorphosis: Speculations on a post-embryonic developmental process lost in amniotes after the water to land transition. Annals Anat. Anat. Anzeiger 2019, 222, 114–119. [Google Scholar] [CrossRef]
- Yun, M.H.; Davaapil, H.; Brockes, J.P. Recurrent turnover of senescent cells during regeneration of a complex structure. eLife 2015, 4, e05505. [Google Scholar] [CrossRef]
- Sousa-Victor, P.; García-Prat, L.; Serrano, A.L.; Perdiguero, E.; Munoz-Canoves, P. Muscle stem cell aging: Regulation and rejuvenation. Trends Endocrin. Metabol. 2015, 26, 287–296. [Google Scholar] [CrossRef]
- Tümpel, S.; Rudolph, K.L. Quiescence: Good and bad of stem cell aging. Trends Cell. Biol. 2019, 29, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.H. Changes in Regenerative Capacity through Lifespan. Int. J. Mol. Sci. 2015, 16, 25392–25432. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Microscopic observations show invasion of inflammatory cells in the limb blastema and epidermis in pre-metamorphic frog tadpoles which destroy the Apical Epidermal CAP and impede regeneration. Ann Anat. 2017, 210, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Odelberg, S.J. Cellular plasticity in vertebrate regeneration. Anat. Rec. Part B New Anat. 2005, 287, 25–35. [Google Scholar] [CrossRef]
- Okada, T.S. Transdifferentiation; Clarendon Press: Oxford, UK, 1991; 238p. [Google Scholar]
- Holliday, R. Epigenetics: A historical overview. Epigenetics 2006, 1, 76–80. [Google Scholar] [CrossRef]
- Katsuyama, T.; Paro, R. Epigenetic reprogramming during tissue regeneration. FEBS Lett. 2011, 585, 1617–1624. [Google Scholar] [CrossRef]
- Maki, N.; Suetsugu-Maki, R.; Sano, S.; Nakamura, K.; Nishimura, O.; Tarui, H.; Del Rio-Tsonis, K.; Ohsumi, K.; Agata, K.; Tsonis, P.A. Oocyte-type linker histone B4 is required for transdifferentiation of somatic cells in vivo. FASEB J. 2010, 24, 3462–3467. [Google Scholar] [CrossRef]
- Maki, N.; Takechi, K.; Sano, S.; Tarui, H.; Sasai, Y.; Agata, K. Rapid accumulation of nucleostemin in nucleolus during newt regeneration. Dev. Dyn. 2007, 263, 941–950. [Google Scholar] [CrossRef]
- Maki, N.; Tsonis, P.A.; Agata, K. Changes in global histone modifications during dedifferentiation in newt lens regeneration. Mol. Vis. 2010, 16, 1893–1897. [Google Scholar]
- Stewart, S.; Tsun, Z.Y.; Izpisua Belmonte, J.C. A histone demethylase is necessary for regeneration in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 19889–19894. [Google Scholar] [CrossRef]
- Akerberg, A.A.; Henner, A.; Stewart, S.; Stankunas, K. Histone demethylases Kdm6ba and Kdm6bb redundantly promote cardiomyocyte proliferation during zebrafish heart ventricle maturation. Dev. Biol. 2017, 426, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Garza-Garcia, A.A.; Driscoll, P.C.; Brockes, J.P. Evidence for the local evolution of mechanisms underlying limb regeneration in salamanders. Integr. Comput. Biol. 2010, 50, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.S.; Tereshina, M.B.; Ermakova, G.V.; Belousov, V.V.; Zaraisky, A.G. Agr genes, missing in amniotes, are involved in the body appendages regeneration in frog tadpoles. Sci Rep. 2013, 3, 1279. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, A.S.; Korotkova, D.D.; Ermakova, G.V.; Belousov, V.V.; Zaraisky, A.G. Ras-dva small GTPases lost during evolution of amniotes regulate regeneration in anamniotes. Sci. Rep. 2018, 8, 13035. [Google Scholar] [CrossRef]
- Kumar, A.; Godwin, J.W.; Gates, P.B.; Garza-Garcia, A.A.; Brockes, J.P. Molecular Basis for the Nerve Dependence of Limb Regeneration in an Adult Vertebrate. Science 2007, 318, 772–777. [Google Scholar] [CrossRef]
- Kumar, A.; Delgado, J.-P.; Gates, P.B.; Neville, G.; Forge, A.; Brockes, J.P. The aneurogenic limb identifies developmental cell interactions underlying vertebrate limb regeneration. Proc. Natl. Acad. Sci. USA 2011, 108, 13588–13593. [Google Scholar] [CrossRef]
- Blassberg, R.A.; Garza-Garcia, A.; Janmohamed, A.; Gates, P.B.; Brockes, J.P. Functional convergence of signaling by GPI-anchored and anchorless forms of a salamander protein implicated in limb regeneration. J. Cell Sci. 2011, 124, 47–56. [Google Scholar] [CrossRef]
- Alibardi, L. Appendage regeneration in anamniotes utilizes genes active during larval-metamorphic stages that have been lost or altered in amniotes: The case for studying lizard tail regeneration. J. Morphol. 2020, 281, 1358–1381. [Google Scholar] [CrossRef]
- Suzuki, N.; Ochi, H. Regeneration enhancers: A clue to reactivation of developmental genes. Dev. Growth Differ. 2020, 62, 343–354. [Google Scholar] [CrossRef]
- Seifert, A.W.; Monaghan, J.R.; Smith, M.D.; Pasch, B.; Stier, A.C.; Michonneau, F.; Maden, M. The influence of fundamental traits on mechanisms controlling appendage regeneration. Biol. Rev. Camb. Philos. Soc. 2012, 87, 330–345. [Google Scholar] [CrossRef]
- Seifert, A.W.; Voss, S.R. Revisiting the relationship between regenerative ability and aging. BMC Biol. 2013, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, K.M.; Chittur, S.V.; Szaro, B.G. Metamorphosis and the regenerative capacity of spinal cord axons in Xenopus laevis. Eur. J. Neurosci. 2011, 33, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Lui, K.O.; Zangi, L.; Chien, K.R. Cardiovascular regenerative therapeutics via synthetic paracrine factor modified mRNA. Stem Cell Res. 2014, 13, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Denoël, M.; Joly, P. Neoteny and progenesis as two heterochronic processes involved in paedomorphosis in Triturus alpestris (Amphibia: Caudata). Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 1481–1485. [Google Scholar] [CrossRef]
- Sessions, S.K. Evolutionary cytogenetics in salamanders. Chromosome Res. 2008, 16, 183–201. [Google Scholar] [CrossRef]
- Olmo, E. Nucleotype and cell size in vertebrates: A review. Bas. Appl. Histochem. 1983, 27, 227–256. [Google Scholar]
- Sclavi, B.; Herrick, J. Genome size variation and species diversity in salamanders. J. Evol. Biol. 2019, 32, 278–286. [Google Scholar] [CrossRef]
- Horner, H.A.; MacGregor, H. C-value and cell volume: Their significance in the evolution and development of amphibians. J. Cell Sci. 1983, 63, 135–146. [Google Scholar]
- Smith, J.J.; Putta, S.; Zhu, W.; Pao, G.M.; Verma, I.M.; Hunter, T.; Bryant, S.V.; Gardiner, D.M.; Harkins, T.T.; Voss, S.R. Genic regions of a large salamander genome contain long introns and novel genes. BMC Genom. 2009, 10, 19. [Google Scholar] [CrossRef]
- Roth, G.; Walk1owiak, W. The Influence of Genome and Cell Size on Brain Morphology in Amphibians. Cold Spring Harb. Perspect. Biol. 2015, 7, a019075. [Google Scholar] [CrossRef]
- Nowoshilow, S.; Schloissnig, S.; Fei, J.F.; Dahl, A.; Pang, A.W.C.; Pippel, M.; Winkler, S.; Hastie, A.R.; Young, G.; Roscito, J.G.; et al. The axolotl genome and the evolution of key tissue formation regulators. Nature 2018, 554, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Herrick, J.; Sclavi, B. A new look at genome size, evolutionary duration and genetic variation in salamanders. C. R. Palevol. 2014, 13, 611–621. [Google Scholar] [CrossRef]
- Prehn, R.T. Regeneration versus neoplastic growth. Carcinogenesis 1997, 18, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Pomerantz, J.H.; Blau, H.M. Tumor suppressors: Enhancers or suppressors of regeneration? Development 2013, 140, 2502–2512. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M. Simultaneous demonstration of lens regeneration from dorsal iris and tumour production from ventral iris in the same newt eye after carcinogen administration. Differentiation 1997, 61, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Anton, H.J.; Schpiller, M.; Grigoryan, E.N. DNA content in cells of cutaneous glands of tailed amphibians. Rep. USSR Acad. Sci. 1993, 332, 251–253. [Google Scholar]
- Sessions, S.K.; Larson, A. Developmental correlates of genome size in plathodontid salamanders and their implications for genome evolution. Evolution 1987, 41, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Stroeva, O.G.; Mitashov, V.I. Retinal pigment epithelium: Proliferation and differentiation during development and regeneration. Int. Rev. Cytol. 1983, 83, 221–293. [Google Scholar]
- Mitashov, V.I. Mechanisms of retina regeneration in urodeles. Int. J. Dev. Biol. 1996, 40, 833–844. [Google Scholar]
- Chiba, C.; Mitashov, V.I. Cellular and molecular events in the adult newt retinal regeneration. In Strategies for Retinal Tissue Repair and Regeneration in Vertebrates: From Fish to Human; Chiba, C., Ed.; Research Signpost: Kerala, India, 2007; pp. 15–33. [Google Scholar]
- Grigoryan, E. Shared triggering mechanisms of retinal regeneration in lower vertebrates and retinal rescue in higher ones. In Tissue Regeneration—From Basic Biology to Clinical Application; Davies, J., Ed.; In Tech: Rijeka, Croatia, 2012; pp. 145–164. [Google Scholar]
- Grigoryan, E.N. Potential Endogenous Cell Sources for Retinal Regeneration in Vertebrates and Humans: Progenitor Traits and Specialization. Biomedicines 2020, 8, 208. [Google Scholar] [CrossRef]
- Grigoryan, E.N.; Markitantova, Y.V. Cellular and molecular preconditions for retinal pigment epithelium (RPE) natural reprogramming during retinal regeneration in Urodela. Biomedicines 2016, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Leghissa, S. L’evoluzione del tetto ottico nei bassi vertebrati (I). Arch. Ital Anat. Embriol. 1962, 67, 343–413. [Google Scholar]
- Keefe, J.R. An analysis of urodelean retinal regeneration. J. Exp. Zool. 1973, 184, 185–257. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, E.N.; Ivanova, I.P.; Poplinskaya, V. A Discovery of new internal sources of the neural retina regeneration after its detachment in newts. I. Morphological and quantitative studies. Biol. Bull. 1996, 23, 263–274. [Google Scholar]
- Grigoryan, E.N. Alternative intrinsic cell sources for neural retina regeneration in adult urodelean amphibians. In Strategies for Retinal Tissue Repair and Regeneration in Vertebrates: From Fish to Human; Chiba, C., Ed.; Research Signpost: Kerala, India, 2007; pp. 35–62. [Google Scholar]
- Mitashov, V.I. Retinal regeneration in amphibians. Int. J. Dev. Biol. 1997, 41, 893–905. [Google Scholar]
- Mitashov, V.I.; Panova, I.G.; Koussoulakos, C. Transdifferential potential of ciliary and pigment epithelial cells in lower vertebrates and mammals. Biol. Bull. 2004, 31, 324–331. [Google Scholar] [CrossRef]
- Kaneko, Y.; Hirota, K.; Matsumoto, G.; Hanyu, Y. Expression pattern of a newt Notch homologue in regenerating newt retina. Dev. Brain Res. 2001, 128, 53–62. [Google Scholar] [CrossRef]
- Roth, G.; Blanke, J.; Wakeo, D.B. Cell size predicts morphological complexity in the brains of frogs and salamanders. Proc. Natl. Acad. Sci. USA 1994, 91, 4796–4800. [Google Scholar] [CrossRef]
- Roth, G.; Nishikawa, K.C.; Naujoks-Manteuffel, C.; Schmidt, A.; Wake, D.B. Paedomorphosis and simplification in the nervous system of salamanders. Brain Behav. Evol. 1993, 42, 137–170. [Google Scholar] [CrossRef]
- Roth, G.; Dicke, U.; Nishikawa, K.C. How do ontogeny, morphology and phisiology of sensory systems constrain and direct the evolution of amphibians? Am. Nat. Suppl. 1992, 139, S105–S124. [Google Scholar] [CrossRef]
- Northcutt, R.G. Evolution of the vertebrate central nervous system: Patterns and processes. Am. Zool. 1984, 24, 701–716. [Google Scholar] [CrossRef]
- ten Donkelaar, H.J. The Central Nervous System of Vertebrates; Nieuwenhuys, R., ten Donkelaar, H.J., Nicholson, C., Eds.; Springer: London, UK, 1998; pp. 1045–1150. [Google Scholar]
- Northcutt, R.G. Lungfish neural characters and their bearing on sarcopterygian phyligeny. J. Morphol. Suppl. 1987, 1, 277–297. [Google Scholar]
- Joven, A.; Morona, R.; Gonzalez, A.; Moreno, N. Expression Patterns of Pax6 and Pax7 in the Adult Brain of a Urodele Amphibian, Pleurodeles waltl. J. Comput. Neurol. Res. Syst. Neurosci. 2013, 521, 2088–2124. [Google Scholar] [CrossRef] [PubMed]
- Joven, A.; Morona, R.; González, A.; Moreno, N. Spatiotemporal patterns of Pax3, Pax6, and Pax7 expression in the developing brain of a urodele amphibian Pleurodeles waltl. J. Comput. Neurol. 2013, 521, 3913–3953. [Google Scholar]
- Avdonin, P.P.; Markitantova, Y.V.; Zinov’eva, R.D.; Mitashov, V.I. Expression of regulatory genes Pax6, Otx2, Six3, and FGF2 during newt retina regeneration. Biol. Bull. 2008, 35, 355–361. [Google Scholar] [CrossRef]
- Avdonin, P.P.; Grigoryan, E.N.; Markitantova, Y.V. Transcriptional factor Pitx2: Localization during triton retina regeneration. Biol. Bull. 2010, 37, 231–235. [Google Scholar] [CrossRef]
- Markitantova, Y.V.; Makar’ev, E.O.; Smirnova, Y.A.; Zinov’eva, R.D.; Mitashov, V.I. Analysis of the expression pattern of regulatory genes Pax6, Prox1, and Six3 during regeneration of eye structures in the newt. Biol. Bull. 2004, 31, 428–436. [Google Scholar] [CrossRef]
- Markitantova, Y.V.; Avdonin, P.P.; Grigoryan, E.N. Nucleostemin expression in the course of in situ reprogramming of eye pigment epithelium cells during retina regeneration in adult newt. Cell Tissue Biol. 2014, 56, 671–672. [Google Scholar]
- Markitantova, Y.V.; Avdonin, P.P.; Grigoryan, E.N. Identification of the gene encoding nucleostemin in the eye tissues of Pleurodeles waltl. Biol. Bull. 2015, 42, 379–386. [Google Scholar] [CrossRef]
- Eguchi, G.; Eguchi, Y.; Nakamura, K. Regenerative capacity in newts is not altered by repeated regeneration and ageing. Nat. Commun. 2011, 2, 1–5. [Google Scholar] [CrossRef]
- Sousounis, K.; Qi, F.; Yadav, M.C.; Millán, J.L.; Toyama, F.; Chiba, C.; Eguchi, Y.; Eguchi, G.; Tsonis, P.A. A robust transcriptional program in newts undergoing multiple events of lens regeneration throughout their lifespan. eLife 2015, 4, e09594. [Google Scholar] [CrossRef] [PubMed]
- Sousounis, K.; Baddour, J.A.; Tsonis, P.A. Aging and regeneration in vertebrates. Curr. Top. Dev. Biol. 2014, 108, 217–246. [Google Scholar]
- Grigoryan, E.N. Molecular factors of the maintenance and activation of the juvenile phenotype of cellular sources for eye tissue regeneration. Biochemistry 2018, 83, 1627–1642. [Google Scholar] [CrossRef] [PubMed]
- Raff, R.A.; Wray, G.A. Heterochrony: Developmental mechanisms and evolutionary results. J. Evol. Biol. 1989, 2, 409–434. [Google Scholar] [CrossRef]
- Zamora, A.J.; Mutin, M. Vimentin and glial fibrillary acidic protein filaments in radial glia of the adult Urodele spinal cord. Neuroscience 1988, 27, 279–288. [Google Scholar] [CrossRef]
- Schnapp, E.; Kragl, M.; Rubin, L.; Tanaka, E.M. Hedgehog signaling controls dorsoventral patterning, blastema cell proliferation and cartilage induction during axolotl tail regeneration. Development 2005, 132, 3243–3253. [Google Scholar] [CrossRef]
- McHedlishvili, L.; Epperlein, H.H.; Telzerow, A.; Tanaka, E.M. A clonal analysis of neural progenitors during axolotl spinal cord regeneration reveals evidence for both spatially restricted and multipotent progenitors. Development 2007, 134, 2083–2093. [Google Scholar] [CrossRef] [PubMed]
- Lust, K.; Tanaka, E.M. A Comparative Perspective on Brain Regeneration in Amphibians and Teleost Fish. Dev. Neurobiol. 2019, 79, 424–436. [Google Scholar] [CrossRef]
- Kaslin, J.; Kroehne, V.; Ganz, J.; Hans, S.; Brand, M. Distinct roles of neuroepithelial-like and radial glia-like progenitor cells in cerebellar regeneration. Development 2017, 144, 1462–1471. [Google Scholar] [CrossRef]
- Tanaka, E.M.; Ferretti, P. Considering the evolution of regeneration in the central nervous system. Nat. Rev. Neurosci. 2009, 10, 713–723. [Google Scholar] [CrossRef]
- Becker, C.G.; Becker, T. Neuronal regeneration from ependymo-radial glial cells: Cook, little pot, cook! Dev. Cell 2015, 32, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Alunni, A.; Bally-Cuif, L. A comparative view of regenerative neurogenesis in vertebrates. Development 2016, 143, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, M.J.; Mysiak, K.S.; Becker, T.; Becker, C.G. Reduce, reuse, recycle—Developmental signals in spinal cord regeneration. Dev. Biol. 2017, 432, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Mitashov, V.I.; Grigoryan, E.N.; Moskovkin, G.N. Autoradiographic investigation of the regeneration of retina and tectum in adult newts. In Ontogenesis of the Brain; V.3.; Trojan, S., Stastny, F., Eds.; Univerzita Karlova: Praha, Chehoslovakia, 1980; pp. 105–108. [Google Scholar]
- Minelli, G.; Franceschini, V.; Del Grande, P.; Ciani, F. Newly-formed neurons in the regenerating optic tectum of Triturus cristatus carnifex. Basic Appl. Histochem. 1987, 31, 43–52. [Google Scholar]
- Okamoto, M.; Ohsawa, H.; Hayashi, T.; Owaribe, K.; Tsonis, P.A. Regeneration of retinotectal projections after optic tectum removal in adult newts. Mol. Vis. 2007, 13, 2112–2118. [Google Scholar]
- Amamoto, R.; Huerta, V.G.L.; Takahashi, E.; Dai, G.; Grant, A.K.; Fu, Z.; Arlotta, P. Adult axolotls can regenerate original neuronal diversity in response to brain injury. eLife 2016, 5, e13998. [Google Scholar] [CrossRef]
- Joven, A.; Simon, A. Homeostatic and regenerative neurogenesis in salamanders. Prog. Neurobiol. 2018, 170, 81–98. [Google Scholar] [CrossRef]
- Tazaki, A.; Tanaka, E.M.; Fei, J.-F. Salamander spinal cord regeneration: The ultimate positive control in vertebrate spinal cord regeneration. Dev. Biol. 2017, 432, 3–71. [Google Scholar] [CrossRef]
- Hass, R. Retrodifferentiation—A mechanism for cellular regeneration? Biol. Chem. 2009, 390, 409–416. [Google Scholar] [CrossRef]
- Du, G.Q.; Shao, Z.B.; Wu, J. Targeted myocardial delivery of GDF11 gene rejuvenates the aged mouse heart and enhances myocardial regeneration after ischemia-reperfusion injury. Basic Res. Cardiol. 2017, 112, 7. [Google Scholar] [CrossRef]
- Pajcini, K.V.; Corbel, C.Y.; Sage, J.; Sage, J.; Pomerantz, J.H.; Blau, H.M. Transient inactivation of Rb and ARF yields regenerative cells from postmitotic mammalian muscle. Cell Stem Cell 2010, 7, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Merrell, A.J.; Stanger, B.Z. Adult cell plasticity in vivo: De-differentiation and transdifferentiation are back in style. Nat. Rev. Mol. Cell Biol. 2016, 17, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Pei, M. Environmental preconditioning rejuvenates adult stem cells’ proliferation and chondrogenic potential. Biomaterials 2017, 117, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Conboy, I.M.; Yousef, H.; Conboy, M.J. Embryonic anti-aging niche. Aging 2011, 3, 555–563. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Favreau-Lessard, A.J.; Ryzhov, S.; Sawyer, D.B. Novel Biological Therapies Targeting Heart Failure: Myocardial Rejuvenation. Heart Fail. Clin. 2016, 12, 461–471. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rochette, L.; Vergely, C. “Pro-youthful” factors in the “labyrinth” of cardiac rejuvenation. Exp. Gerontol. 2016, 83, 1–5. [Google Scholar] [CrossRef]
- Ruckh, M.J.; Zhao, J.W.; Shadrach, J.L.; Nageswara Rao, T.; Wagers, A.J.; Franklin, R.J.M. Rejuvenation of Regeneration in the Aging Central Nervous System. Cell 2012, 10, 96–103. [Google Scholar] [CrossRef]
- Smith, L.K.; White, C.W., III; Villeda, S.A. The systemic environment: At the interface of aging and adult neurogenesis. Cell Tissue Res. 2018, 371, 105–113. [Google Scholar] [CrossRef]
- Conover, J.C. The neural stem cell niche. Cell Tissue Res. 2008, 331, 211–224. [Google Scholar] [CrossRef]
- Katsimpardi, L.; Litterman, N.K.; Schein, P.A.; Miller, C.M.; Loffredo, F.S.; Wojtkiewicz, J.R.; Chen, J.W.; Lee, R.T.; Wagers, A.J.; Rubin, L.L. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science 2014, 344, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Baulier, E.; Ke, Y.; Young, A.; Ahmedli, N.B.; Schwartz, S.D.; Farber, D.B. Human embryonic stem cells extracellular vesicles and their effects on immortalized human retinal Muller cells. PLoS ONE 2018, 13, e0194004. [Google Scholar] [CrossRef] [PubMed]
- Katsman, D.; Stackpole, E.J.; Domin, D.R.; Farber, D.B. Embryonic stem cell-derived microvesicles induce gene expression changes in Muller cells of the retina. PLoS ONE 2012, 7, e50417. [Google Scholar] [CrossRef] [PubMed]
- Quesenberry, P.J.; Aliotta, J.M. Cellular phenotype switching and microvesicles. Adv. Drug Deliv. Rev. 2010, 62, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Burrola, S.; Wu, J.; Ding, W.-Q. Extracellular Vesicles in the Development of Cancer Therapeutics. Int. J. Mol. Sci. 2020, 21, 6097. [Google Scholar] [CrossRef]
- Basu, J.; Ludlow, J.W. Exosomes for repair, regeneration and rejuvenation. Expert Opin. Biol. Ther. 2016, 16, 489–506. [Google Scholar] [CrossRef]
- Arenas Gómez, C.M.; Molina, A.G.; Zapata, J.D.; Delgado, J.P. Limb regeneration in a direct-developing terrestrial salamander, Bolitoglossa ramosi (Caudata: Plethodontidae). Limb regeneration in plethodontid salamanders. Regeneration 2017, 4, 227–235. [Google Scholar] [CrossRef]
- Naranjo, J.D.; Scarritt, M.E.; Huleihel, L.; Ravindra, A.; Torres, C.M.; Badylak, S.F. Regenerative Medicine: Lessons from Mother Nature. Regen. Med. 2016, 11, 767–775. [Google Scholar] [CrossRef]
- Stocum, D.L. Regenerative Biology and Medicine; Academic Press: Burlington, UK, 2006. [Google Scholar]
- McGann, C.J.; Odelberg, S.J.; Keating, M.T. Mammalian myotube dedifferentiation induced by newt regeneration extract. Proc. Natl. Acad. Sci. USA 2001, 98, 13699–13704. [Google Scholar] [CrossRef]
- Tanaka, E.M.; Drechsel, D.N.; Brockes, J.P. Thrombin regulates S-phase re-entry by cultured newt myotubes. Curr. Biol. 1999, 12, 792–799. [Google Scholar] [CrossRef]
- Kim, H.-J.; Archer, E.; Escobedo, N.; Tapscott, S.J.; Unguez, G.A. Inhibition of Mammalian Muscle Differentiation by Regeneration Blastema Extract of Sternopygus macrurus. Dev. Dyn. 2008, 237, 2830–2843. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kawesa, S.; Vanstone, J.; Tsilfidis, C. Differential response to newt regeneration extract by C2C12 and primary mammalian muscle cells. Skeletal Muscle 2015, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Unguez, G.A.; Kim, H.-J.; Tapscott, S. Inhibition of mammalian muscle differentiation by blastema extract of S. macrurus. Dev. Biol. 2006, 295, 354–355. [Google Scholar] [CrossRef][Green Version]
- Chen, W.C.; Wang, Z.; Missinato, M.A. Decellularized zebrafish cardiac extracellular matrix induces mammalian heart regeneration. Sci. Adv. 2016, 2, e1600844. [Google Scholar] [CrossRef]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef]
- Porrello, E.R.; Olson, E.N. A neonatal blueprint for cardiac regeneration. Stem Cell Res. 2014, 13, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Huynh, N.; VonMoss, L.; Smith, D.; Rahman, I.; Felemban, M.F.; Zuo, J.; Rody, W.J.; McHugh, K.P.; Holliday, L.S. Characterization of Regulatory Extracellular Vesicles from Osteoclasts. J. Dent. Res. 2016, 95, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Ohgo, S.H.; Sakamoto, T.; Nakajima, W.; Matsunaga, S.; Wada, N. Visualization of extracellular vesicles in the regenerating caudal fin blastema of zebrafish using in vivo electroporation. Biochem. Biophys Res. Commun. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Middleton, R.C.; Rogers, R.G.; De Couto, G.; Tseliou, E.; Luther, K.; Holewinski, R.; Soetkamp, D.; Van Eyk, J.E.; Antes, T.J.; Marbán, E. Newt cells secrete extracellular vesicles with therapeutic bioactivity in mammalian cardiomyocytes. J. Extracell Vesicles 2018, 7, 1456888. [Google Scholar] [CrossRef] [PubMed]
- Gancharova, O.S. The Modeling of a Damage and Restoration of Eye Tissues and Lacrimal Gland. Ph.D. Thesis, Lomonosov Moscow State University, Moscow, Russia, 2017. (In Russian). [Google Scholar]
- Markitantova, Y.V.; Avdonin, P.P.; Grigoryan, E.N. FGF2 signaling pathway components in tissues of the posterior eye sector in the adult newt Pleurodeles waltl. Biol. Bull. 2014, 41, 297–305. [Google Scholar] [CrossRef]
- Novikova, Y.P.; Grigoryan, E.N. Early appearance of aging signs in retinal pigment epithelium in young albino rats. Russ. J. Dev. Biol. 2020, 51. in press. [Google Scholar] [CrossRef]
- Shafei, E.V.; Rzhanova, L.A.; Novikova, Y.P.; Kurinov, A.M.; Grigoryan, E.N.; Aleksandrova, M.A.; Kuznetsova, A.V. Response of human pigment epithelial cells to the effect of the media conditioned of newt retinal regenerates. Cell Tissue Biol. 2021, 15. in press. [Google Scholar]
- Flament, S.; Dumond, H.; Chardard, D.; Chesnel, A. Lifelong testicular differentiation in Pleurodeles waltl (Amphibia, Caudata). Reprod. Biol. Endocrinol. 2009, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Witman, N.; Murtuza, B.; Davis, B.; Arner, A.; Morrison, J.I. Recapitulation of developmental cardiogenesis governs the morphological and functional regeneration of adult newt hearts following injury. Dev. Biol. 2011, 354, 67–76. [Google Scholar] [CrossRef]
- Lin, B.; Srikanth, P.; Castle, A.C.; Nigwekar, S.; Malhotra, R.; Galloway, J.L.; Sykes, D.B.; Rajagopal, J. Modulating cell fate as a therapeutic strategy. Cell Stem Cell 2018, 23, 329–341. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigoryan, E.N. Study of Natural Longlife Juvenility and Tissue Regeneration in Caudate Amphibians and Potential Application of Resulting Data in Biomedicine. J. Dev. Biol. 2021, 9, 2. https://doi.org/10.3390/jdb9010002
Grigoryan EN. Study of Natural Longlife Juvenility and Tissue Regeneration in Caudate Amphibians and Potential Application of Resulting Data in Biomedicine. Journal of Developmental Biology. 2021; 9(1):2. https://doi.org/10.3390/jdb9010002
Chicago/Turabian StyleGrigoryan, Eleonora N. 2021. "Study of Natural Longlife Juvenility and Tissue Regeneration in Caudate Amphibians and Potential Application of Resulting Data in Biomedicine" Journal of Developmental Biology 9, no. 1: 2. https://doi.org/10.3390/jdb9010002
APA StyleGrigoryan, E. N. (2021). Study of Natural Longlife Juvenility and Tissue Regeneration in Caudate Amphibians and Potential Application of Resulting Data in Biomedicine. Journal of Developmental Biology, 9(1), 2. https://doi.org/10.3390/jdb9010002