Evolution of Developmental GATA Factors in Nematodes



Abstract

{kind=link}
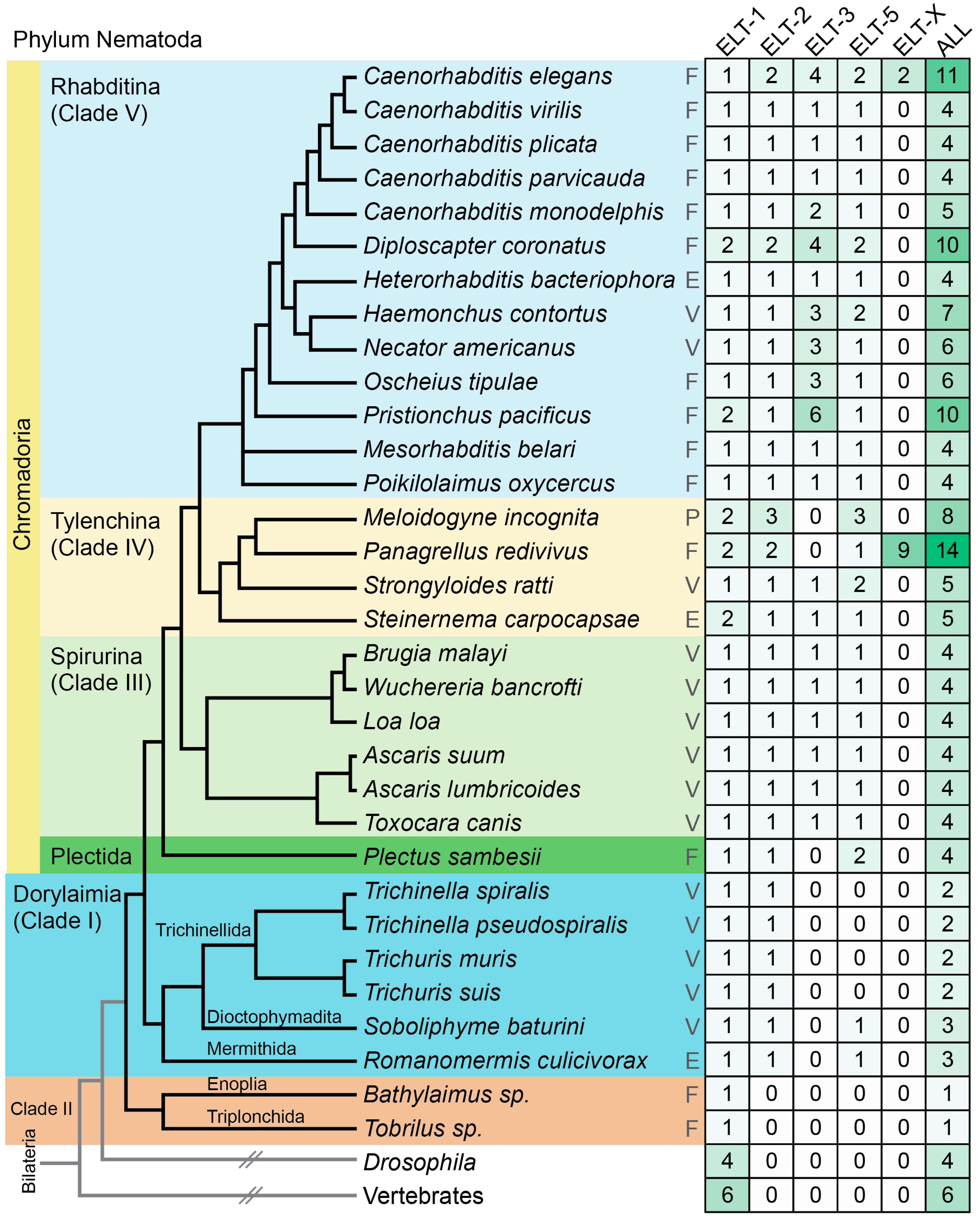{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Identification of GATA Factors in Nematode Species
2.2. Assignment of Putative GATA Factors to Classes
2.3. Reference Genome Sequences
2.4. Construction of Alignments and Relatedness Trees
2.5. Identification of Putative Regulatory Sequence Motifs
2.6. Additional Computational Resources
3. Results
3.1. GATA Factors Are Found across the Nematode Phylum
3.2. GATA Factor Sequences across the Phylum Tend to Be Related by Clade
3.3. Comparison of Nematode GATA Factor Structures
3.4. Introns Reveal Possible Origins of Nematode GATA Factors
3.5. Extra GATA Factors in D. Coronatus, P. Pacificus and P. Redivivus Are Most Closely Related to ELT-3
3.6. Patterns of Amino Acid Conservation in Nematode GATA Factors
3.7. Possible Conserved Regulatory Regions in Flanking Regions
4. Discussion
4.1. Evolution of the GATA Factors in Nematodes
4.2. What Do the Nematode GATA Factors Do?
4.3. GATA Factor Expansion Favors ELT-3-Like Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parkinson, J.; Mitreva, M.; Whitton, C.; Thomson, M.; Daub, J.; Martin, J.W.L.; Schmid, R.; Hall, N.; Barrell, B.; Waterston, R.H.; et al. A transcriptomic analysis of the phylum Nematoda. Nat. Genet. 2004, 36, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Blaxter, M.; De Ley, P.; Garey, J.R.; Liu, L.X.; Scheldeman, P.; Vierstraete, A.; Vanfleteren, J.R.; Mackey, L.Y.; Dorris, M.; Frisse, L.M.; et al. A molecular evolutionary framework for the phylum Nematoda. Nat. Cell Biol. 1998, 392, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kern, E.; Kim, T.; Sim, M.; Kim, J.; Kim, Y.; Park, C.; Nadler, S.A.; Park, J.-K. Phylogenetic analysis of two Plectus mitochondrial genomes (Nematoda: Plectida) supports a sister group relationship between Plectida and Rhabditida within Chromadorea. Mol. Phylogenetics Evol. 2017, 107, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Aguinaldo, A.M.A.; Turbeville, J.M.; Linford, L.S.; Rivera, M.C.; Garey, J.R.; Raff, R.A.; Lake, J.A. Evidence for a clade of nematodes, arthropods and other moulting animals. Nat. Cell Biol. 1997, 387, 489–493. [Google Scholar] [CrossRef]
- Schulze, J.; Schierenberg, E. Evolution of embryonic development in nematodes. EvoDevo 2011, 2, 18. [Google Scholar] [CrossRef]
- Sulston, J.; Schierenberg, E.; White, J.; Thomson, J. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 1983, 100, 64–119. [Google Scholar] [CrossRef]
- Laugsch, M.; Schierenberg, E. Differences in maternal supply and early development of closely related nematode species. Int. J. Dev. Biol. 2004, 48, 655–662. [Google Scholar] [CrossRef]
- Schierenberg, E. Three sons of fortune: Early embryogenesis, evolution and ecology of nematodes. BioEssays 2001, 23, 841–847. [Google Scholar] [CrossRef]
- Schierenberg, E. Embryological variation during nematode development. WormBook 2006, 1–13. [Google Scholar] [CrossRef]
- Schierenberg, E. Unusual cleavage and gastrulation in a freshwater nematode: Developmental and phylogenetic implications. Dev. Genes Evol. 2004, 215, 103–108. [Google Scholar] [CrossRef]
- Lowry, J.A.; Atchley, W.R. Molecular Evolution of the GATA Family of Transcription Factors: Conservation Within the DNA-Binding Domain. J. Mol. Evol. 2000, 50, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Chudzicka-Ormaniec, P.; Macios, M.; Koper, M.; Weedall, G.D.; Caddick, M.X.; Weglenski, P.; Dzikowska, A. The role of the GATA transcription factor AreB in regulation of nitrogen and carbon metabolism in Aspergillus nidulans. Fems Microbiol. Lett. 2019, 366, 366. [Google Scholar] [CrossRef] [PubMed]
- Macios, M.; Caddick, M.X.; Weglenski, P.; Scazzocchio, C.; Dzikowska, A. The GATA factors AREA and AREB together with the co-repressor NMRA, negatively regulate arginine catabolism in Aspergillus nidulans in response to nitrogen and carbon source. Fungal Genet. Biol. 2012, 49, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, Y.; Xiao, Z.; Yang, H.; Hao, Q.; Yuan, S.; Chen, H.; Chen, L.; Chen, S.; Zhou, X.; et al. A GATA Transcription Factor from Soybean (Glycine max) Regulates Chlorophyll Biosynthesis and Suppresses Growth in the Transgenic Arabidopsis thaliana. Plants 2020, 9, 1036. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.-H.; Zubo, Y.O.; Tapken, W.; Kim, H.J.; Lavanway, A.M.; Howard, L.; Pilon, M.; Kieber, J.J.; Schaller, G.E. Functional Characterization of the GATA Transcription Factors GNC and CGA1 Reveals Their Key Role in Chloroplast Development, Growth, and Division in Arabidopsis. Plant. Physiol. 2012, 160, 332–348. [Google Scholar] [CrossRef]
- Bi, Y.-M.; Zhang, Y.; Signorelli, T.; Zhao, R.; Zhu, T.; Rothstein, S. Genetic analysis of Arabidopsis GATA transcription factor gene family reveals a nitrate-inducible member important for chlorophyll synthesis and glucose sensitivity. Plant. J. 2005, 44, 680–692. [Google Scholar] [CrossRef]
- Patient, R.K.; McGhee, J.D. The GATA family (vertebrates and invertebrates). Curr. Opin. Genet. Dev. 2002, 12, 416–422. [Google Scholar] [CrossRef]
- Fujiwara, T. GATA Transcription Factors: Basic Principles and Related Human Disorders. Tohoku J. Exp. Med. 2017, 242, 83–91. [Google Scholar] [CrossRef]
- De Madrid, B.H.; Casanova, J. GATA factor genes in the Drosophila midgut embryo. PLoS ONE 2018, 13, e0193612. [Google Scholar] [CrossRef]
- Zarin, A.A.; Daly, A.C.; Hülsmeier, J.; Asadzadeh, J.; Labrador, J.-P. A GATA/homeodomain transcriptional code regulates axon guidance through the Unc-5 receptor. Development 2012, 139, 1798–1805. [Google Scholar] [CrossRef]
- Tokusumi, Y.; Tokusumi, T.; Stoller-Conrad, J.; Schulz, R.A. Serpent, suppressor of hairless and U-shaped are crucial regulators of hedgehog niche expression and prohemocyte maintenance during Drosophila larval hematopoiesis. Development 2010, 137, 3561–3568. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, M.; Sanchez-Ferras, O.; Bouchard, M. GATA transcription factors in development and disease. Development 2018, 145, dev164384. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Zhou, Y.; Han, X.; Shen, C.; Wang, S.; Liu, C.; Yin, W.; Xia, X. The GATA transcription factor GNC plays an important role in photosynthesis and growth in poplar. J. Exp. Bot. 2019, 71, 1969–1984. [Google Scholar] [CrossRef] [PubMed]
- Starich, M.R.; Wikström, M.; Arst, H.N.; Clore, G.; Gronenborn, A.M. The solution structure of a fungal AREA protein-DNA complex: An alternative binding mode for the basic carboxyl tail of GATA factors 1 1Edited by P. E. Wright. J. Mol. Biol. 1998, 277, 605–620. [Google Scholar] [CrossRef]
- Gillis, W.Q.; Bowerman, B.; Schneider, S.Q. The evolution of protostome GATA factors: Molecular phylogenetics, synteny, and intron/exon structure reveal orthologous relationships. BMC Evol. Biol. 2008, 8, 112. [Google Scholar] [CrossRef]
- Gillis, W.Q.; John, J.S.; Bowerman, B.; Schneider, S.Q. Whole genome duplications and expansion of the vertebrate GATA transcription factor gene family. BMC Evol. Biol. 2009, 9, 207. [Google Scholar] [CrossRef]
- Lentjes, M.H.F.M.; Niessen, H.E.C.; Akiyama, Y.; De Bruine, A.P.; Melotte, V.; Van Engeland, M. The emerging role of GATA transcription factors in development and disease. Expert Rev. Mol. Med. 2016, 18, e3. [Google Scholar] [CrossRef]
- Martin, D.I.; Orkin, S.H. Transcriptional activation and DNA binding by the erythroid factor GF-1/NF-E1/Eryf 1. Genes Dev. 1990, 4, 1886–1898. [Google Scholar] [CrossRef]
- Yang, H.Y.; Evans, T. Distinct roles for the two cGATA-1 finger domains. Mol. Cell. Biol. 1992, 12, 4562–4570. [Google Scholar] [CrossRef]
- Waltzer, L.; Bataillé, L.; Peyrefitte, S.; Haenlin, M. Two isoforms of Serpent containing either one or two GATA zinc fingers have different roles in Drosophila haematopoiesis. EMBO J. 2002, 21, 5477–5486. [Google Scholar] [CrossRef]
- Gilleard, J.; McGhee, J.D. Activation of Hypodermal Differentiation in theCaenorhabditis elegans Embryo by GATA Transcription Factors ELT-1 and ELT-3. Mol. Cell. Biol. 2001, 21, 2533–2544. [Google Scholar] [CrossRef]
- Fukushige, T.; Hawkins, M.G.; McGhee, J.D. The GATA-factor elt-2 is essential for formation of the Caenorhabditis elegans intestine. Dev. Biol. 1998, 198, 286–302. [Google Scholar] [CrossRef]
- Page, B.D.; Zhang, W.; Steward, K.; Blumenthal, T.; Priess, J.R. ELT-1, a GATA-like transcription factor, is required for epidermal cell fates in Caenorhabditis elegans embryos. Genes Dev. 1997, 11, 1651–1661. [Google Scholar] [CrossRef]
- Koh, K.; Rothman, J.H. ELT-5 and ELT-6 are required continuously to regulate epidermal seam cell differentiation and cell fusion in C elegans. Development 2001, 128, 2867–2880. [Google Scholar] [CrossRef]
- Maduro, M.F.; Meneghini, M.D.; Bowerman, B.; Broitman-Maduro, G.; Rothman, J.H. Restriction of mesendoderm to a single blastomere by the combined action of SKN-1 and a GSK-3beta homolog is mediated by MED-1 and -2 in C. elegans. Mol. Cell 2001, 7, 475–485. [Google Scholar] [CrossRef]
- Maduro, M.F.; Hill, R.J.; Heid, P.J.; Newman-Smith, E.D.; Zhu, J.; Priess, J.R.; Rothman, J.H. Genetic redundancy in endoderm specification within the genus Caenorhabditis. Dev. Biol. 2005, 284, 509–522. [Google Scholar] [CrossRef]
- Maduro, M.F. Gut Development in C. elegans. Semin Cell Dev. Biol. 2017, 3–11. [Google Scholar] [CrossRef]
- Sommermann, E.M.; Strohmaier, K.R.; Maduro, M.F.; Rothman, J.H. Endoderm development in Caenorhabditis elegans: The synergistic action of ELT-2 and -7 mediates the specification→differentiation transition. Dev. Biol. 2010, 347, 154–166. [Google Scholar] [CrossRef]
- Fukushige, T.; Goszczynski, B.; Tian, H.; McGhee, J.D. The Evolutionary Duplication and Probable Demise of an Endodermal GATA Factor in Caenorhabditis elegans. Genetics 2003, 165, 575–588. [Google Scholar]
- Maduro, M.F. Evolutionary Dynamics of the SKN-1 → MED → END-1,3 Regulatory Gene Cascade in Caenorhabditis Endoderm Specification. G3 (Bethesda) 2019, 10, 333–356. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information Basic Local Alignment Search Tool (NCBI BLAST). Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 19 October 2020).
- Grove, C.; Cain, S.; Chen, W.J.; Davis, P.; Harris, T.; Howe, K.L.; Kishore, R.; Lee, R.; Paulini, M.; Raciti, D.; et al. Using WormBase: A Genome Biology Resource for Caenorhabditis elegans and Related Nematodes. Methods Mol. Biol. 2018, 1757, 399–470. [Google Scholar] [CrossRef]
- Caenorhabditis Genomes Project. Available online: https://caenorhabditis.org (accessed on 19 October 2020).
- Howe, K.L.; Bolt, B.J.; Shafie, M.; Kersey, P.; Berriman, M. WormBase ParaSite—A comprehensive resource for helminth genomics. Mol. Biochem. Parasitol. 2017, 215, 2–10. [Google Scholar] [CrossRef]
- Slos, D.; Sudhaus, W.; Stevens, L.; Bert, W.; Blaxter, M. Caenorhabditis monodelphis sp. n.: Defining the stem morphology and genomics of the genus Caenorhabditis. BMC Zoöl. 2017, 2, 4. [Google Scholar] [CrossRef]
- McLean, F.; Berger, D.; Laetsch, D.R.; Schwartz, H.T.; Blaxter, M. Improving the annotation of the Heterorhabditis bacteriophora genome. GigaScience 2018, 7. [Google Scholar] [CrossRef]
- Grosmaire, M.; Launay, C.; Siegwald, M.; Brugière, T.; Estrada-Virrueta, L.; Berger, D.; Burny, C.; Modolo, L.; Blaxter, M.; Meister, P.; et al. Males as somatic investment in a parthenogenetic nematode. Science 2019, 363, 1210–1213. [Google Scholar] [CrossRef]
- Beltran, T.; Barroso, C.; Birkle, T.Y.; Stevens, L.; Schwartz, H.T.; Sternberg, P.W.; Fradin, H.; Gunsalus, K.; Piano, F.; Sharma, G.; et al. Comparative Epigenomics Reveals that RNA Polymerase II Pausing and Chromatin Domain Organization Control Nematode piRNA Biogenesis. Dev. Cell 2019, 48, 793–810.e6. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 11, 43–48. [Google Scholar] [CrossRef]
- Vaughan, T.G. IcyTree: Rapid browser-based visualization for phylogenetic trees and networks. Bioinformatics 2017, 33, 2392–2394. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. In Proceedings of the International Conference on Intelligent Systems for Molecular Biology, Stanford, CA, USA, 14–17 August 1994; Volume 2, pp. 28–36. [Google Scholar]
- Weirauch, M.T.; Yang, A.; Albu, M.; Cote, A.G.; Montenegro-Montero, A.; Drewe, P.; Najafabadi, H.S.; Lambert, S.A.; Mann, I.; Cook, K.; et al. Determination and inference of eukaryotic transcription factor sequence specificity. Cell 2014, 158, 1431–1443. [Google Scholar] [CrossRef] [PubMed]
- Stevens, L.; Felix, M.A.; Beltran, T.; Braendle, C.; Caurcel, C.; Fausett, S.; Fitch, D.; Frezal, L.; Gosse, C.; Kaur, T.; et al. Comparative genomics of 10 new Caenorhabditis species. Evol. Let. 2019, 3, 217–236. [Google Scholar] [CrossRef] [PubMed]
- Smythe, A.B.; Holovachov, O.; Kocot, K.M. Improved phylogenomic sampling of free-living nematodes enhances resolution of higher-level nematode phylogeny. BMC Evol. Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, N.; Kiontke, K.; Tanaka, R.; Hirooka, Y.; Schwarz, A.; Muller-Reichert, T.; Chaudhuri, J.; Pires-daSilva, A. Description of two three-gendered nematode species in the new genus Auanema (Rhabditina) that are models for reproductive mode evolution. Sci. Rep. 2017, 7, 11135. [Google Scholar] [CrossRef]
- Gabriel, W.N.; McNuff, R.; Patel, S.K.; Gregory, T.R.; Jeck, W.R.; Jones, C.D.; Goldstein, B. The tardigrade Hypsibius dujardini, a new model for studying the evolution of development. Dev. Biol. 2007, 312, 545–559. [Google Scholar] [CrossRef]
- Hunt, V.L.; Tsai, I.J.; Coghlan, A.; Reid, A.J.; Holroyd, N.; Foth, B.J.; Tracey, A.; Cotton, J.A.; Stanley, E.J.; Beasley, H.; et al. The genomic basis of parasitism in the Strongyloides clade of nematodes. Nat. Genet. 2016, 48, 299–307. [Google Scholar] [CrossRef]
- Omichinski, J.G.; Clore, G.M.; Schaad, O.; Felsenfeld, G.; Trainor, C.; Appella, E.; Stahl, S.J.; Gronenborn, A.M. NMR structure of a specific DNA complex of Zn-containing DNA binding domain of GATA-1. Science 1993, 261, 438–446. [Google Scholar] [CrossRef]
- Contrino, S.; Smith, R.N.; Butano, D.; Carr, A.; Hu, F.; Lyne, R.; Rutherford, K.; Kalderimis, A.; Sullivan, J.; Carbon, S.; et al. modMine: Flexible access to modENCODE data. Nucleic Acids Res. 2011, 40, D1082–D1088. [Google Scholar] [CrossRef]
- McGhee, J.D.; Fukushige, T.; Krause, M.W.; Minnema, S.E.; Goszczynski, B.; Gaudet, J.; Kohara, Y.; Bossinger, O.; Zhao, Y.; Khattra, J.; et al. ELT-2 is the predominant transcription factor controlling differentiation and function of the C. elegans intestine, from embryo to adult. Dev. Biol. 2009, 327, 551–565. [Google Scholar] [CrossRef]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, K.; Lambert, S.A.; Yang, A.W.H.; Riddell, J.; Mnaimneh, S.; Zheng, H.; Albu, M.; Najafabadi, H.S.; Reece-Hoyes, J.S.; Bass, J.I.F.; et al. Mapping and analysis of Caenorhabditis elegans transcription factor sequence specificities. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Morgunova, E.; Jolma, A.; Kaasinen, E.; Sahu, B.; Khund-Sayeed, S.; Das, P.K.; Kivioja, T.; Dave, K.; Zhong, F.; et al. Impact of cytosine methylation on DNA binding specificities of human transcription factors. Science 2017, 356, eaaj2239. [Google Scholar] [CrossRef]
- Nitta, K.R.; Jolma, A.; Yin, Y.; Morgunova, E.; Kivioja, T.; Akhtar, J.; Hens, K.; Toivonen, J.; Deplancke, B.; Furlong, E.E.M.; et al. Conservation of transcription factor binding specificities across 600 million years of bilateria evolution. eLife 2015, 4, e04837. [Google Scholar] [CrossRef]
- Gerstein, M.B.; Kundaje, A.; Hariharan, M.; Landt, S.G.; Yan, K.-K.; Cheng, C.; Mu, X.J.; Khurana, E.; Rozowsky, J.; Alexander, R.P.; et al. Architecture of the human regulatory network derived from ENCODE data. Nat. Cell Biol. 2012, 489, 91–100. [Google Scholar] [CrossRef]
- Coroian, C.; Broitman-Maduro, G.; Maduro, M.F. Med-type GATA factors and the evolution of mesendoderm specification in nematodes. Dev. Biol. 2006, 289, 444–455. [Google Scholar] [CrossRef]
- Wiesenfahrt, T.; Berg, J.Y.; Nishimura, E.O.; Robinson, A.G.; Goszczynski, B.; Lieb, J.D.; McGhee, J.D. The Function and Regulation of the GATA Factor ELT-2 in the C. elegans Endoderm. Development 2016, 483–491. [Google Scholar] [CrossRef]
- Del Castillo-Olivares, A.; Kulkarni, M.; Smith, H.E. Regulation of sperm gene expression by the GATA factor ELT-1. Dev. Biol. 2009, 333, 397–408. [Google Scholar] [CrossRef]
- Yin, J.; Madaan, U.; Park, A.; Aftab, N.; Savage-Dunn, C. Multiple cis elements and GATA factors regulate a cuticle collagen gene in Caenorhabditis elegans. Genes 2015, 53, 278–284. [Google Scholar] [CrossRef]
- Couthier, A.; Smith, J.; McGarr, P.; Craig, B.; Gilleard, J. Ectopic expression of a Haemonchus contortus GATA transcription factor in Caenorhabditis elegans reveals conserved function in spite of extensive sequence divergence. Mol. Biochem. Parasitol. 2004, 133, 241–253. [Google Scholar] [CrossRef]
- Rosa, B.A.; Jasmer, D.P.; Mitreva, M. Genome-Wide Tissue-Specific Gene Expression, Co-expression and Regulation of Co-expressed Genes in Adult Nematode Ascaris suum. PLoS Negl. Trop. Dis. 2014, 8, e2678. [Google Scholar] [CrossRef] [PubMed]
- Block, D.H.S.; Twumasi-Boateng, K.; Kang, H.S.; Carlisle, J.A.; Hanganu, A.; Lai, T.Y.-J.; Shapira, M. The Developmental Intestinal Regulator ELT-2 Controls p38-Dependent Immune Responses in Adult C. elegans. PLoS Genet. 2015, 11, e1005265. [Google Scholar] [CrossRef] [PubMed]
- Mann, F.G.; Van Nostrand, E.L.; Friedland, A.E.; Liu, X.; Kim, S.K. Deactivation of the GATA Transcription Factor ELT-2 Is a Major Driver of Normal Aging in C. elegans. PLoS Genet. 2016, 12, e1005956. [Google Scholar] [CrossRef]
- Kormish, J.D.; McGhee, J.D. The C. elegans lethal gut-obstructed gob-1 gene is trehalose-6-phosphate phosphatase. Dev. Biol. 2005, 287, 35–47. [Google Scholar] [CrossRef]
- Keith, S.A.; Maddux, S.K.; Zhong, Y.; Chinchankar, M.N.; Ferguson, A.A.; Ghazi, A.; Fisher, A.L. Graded Proteasome Dysfunction in Caenorhabditis elegans Activates an Adaptive Response Involving the Conserved SKN-1 and ELT-2 Transcription Factors and the Autophagy-Lysosome Pathway. PLoS Genet. 2016, 12, e1005823. [Google Scholar] [CrossRef]
- Fukushige, T.; Goszczynski, B.; Yan, J.; McGhee, J.D. Transcriptional control and patterning of the pho-1 gene, an essential acid phosphatase expressed in the C. elegans intestine. Dev. Biol. 2005, 279, 446–461. [Google Scholar] [CrossRef]
- Asan, A.; Raiders, S.A.; Priess, J.R. Morphogenesis of the C. elegans Intestine Involves Axon Guidance Genes. PLoS Genet. 2016, 12, e1005950. [Google Scholar] [CrossRef]
- Altun, Z.F.; Hall, D.H. WormAtlas Hermaphrodite Handbook – Alimentary System – Intestine. WormAtlas 2003. [Google Scholar] [CrossRef]
- Lucchetta, E.M.; Ohlstein, B. Amitosis of Polyploid Cells Regenerates Functional Stem Cells in the Drosophila Intestine. Cell Stem Cell 2017, 20, 609–620.e6. [Google Scholar] [CrossRef]
- Koh, K.; Peyrot, S.M.; Wood, C.G.; Wagmaister, J.A.; Maduro, M.F.; Eisenmann, D.M.; Rothman, J.H. Cell fates and fusion in the C. elegans vulval primordium are regulated by the EGL-18 and ELT-6 GATA factors—Apparent direct targets of the LIN-39 Hox protein. Development 2002, 129, 5171–5180. [Google Scholar]
- Gorrepati, L.; Thompson, K.W.; Eisenmann, D.M. C. elegans GATA factors EGL-18 and ELT-6 function downstream of Wnt signaling to maintain the progenitor fate during larval asymmetric divisions of the seam cells. Development 2013, 140, 2093–2102. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eurmsirilerd, E.; Maduro, M.F. Evolution of Developmental GATA Factors in Nematodes. J. Dev. Biol. 2020, 8, 27. https://doi.org/10.3390/jdb8040027
Eurmsirilerd E, Maduro MF. Evolution of Developmental GATA Factors in Nematodes. Journal of Developmental Biology. 2020; 8(4):27. https://doi.org/10.3390/jdb8040027
Chicago/Turabian StyleEurmsirilerd, Ethan, and Morris F. Maduro. 2020. "Evolution of Developmental GATA Factors in Nematodes" Journal of Developmental Biology 8, no. 4: 27. https://doi.org/10.3390/jdb8040027
APA StyleEurmsirilerd, E., & Maduro, M. F. (2020). Evolution of Developmental GATA Factors in Nematodes. Journal of Developmental Biology, 8(4), 27. https://doi.org/10.3390/jdb8040027