Retinoic Acid-Induced Epidermal Transdifferentiation in Skin

Abstract
:
1. Introduction
2. Effects of RA on Morphogenesis of Chick Embryonic Skin
3. Effects of RA on the Expression of Homeobox Genes in Chick Embryonic Skin
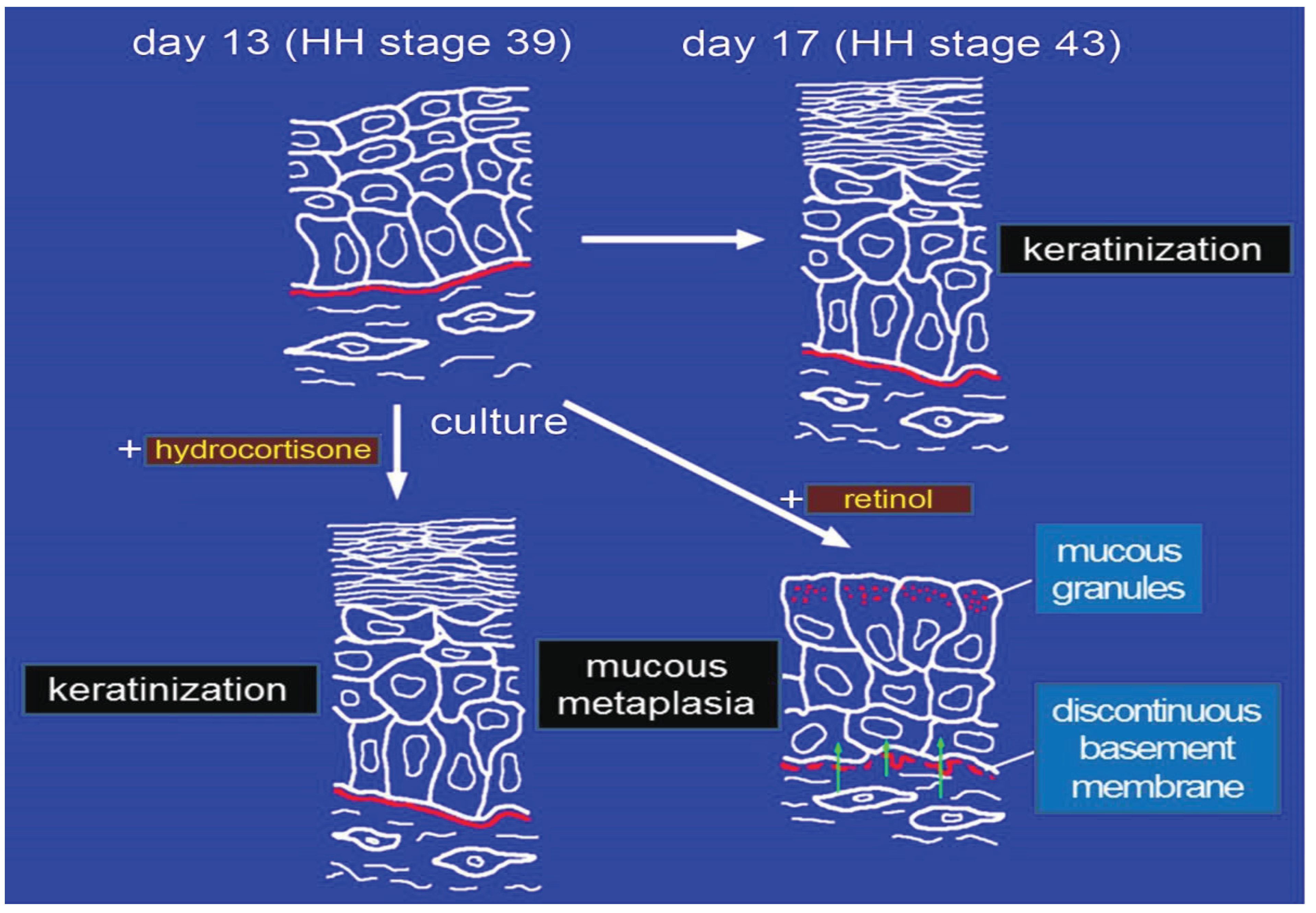
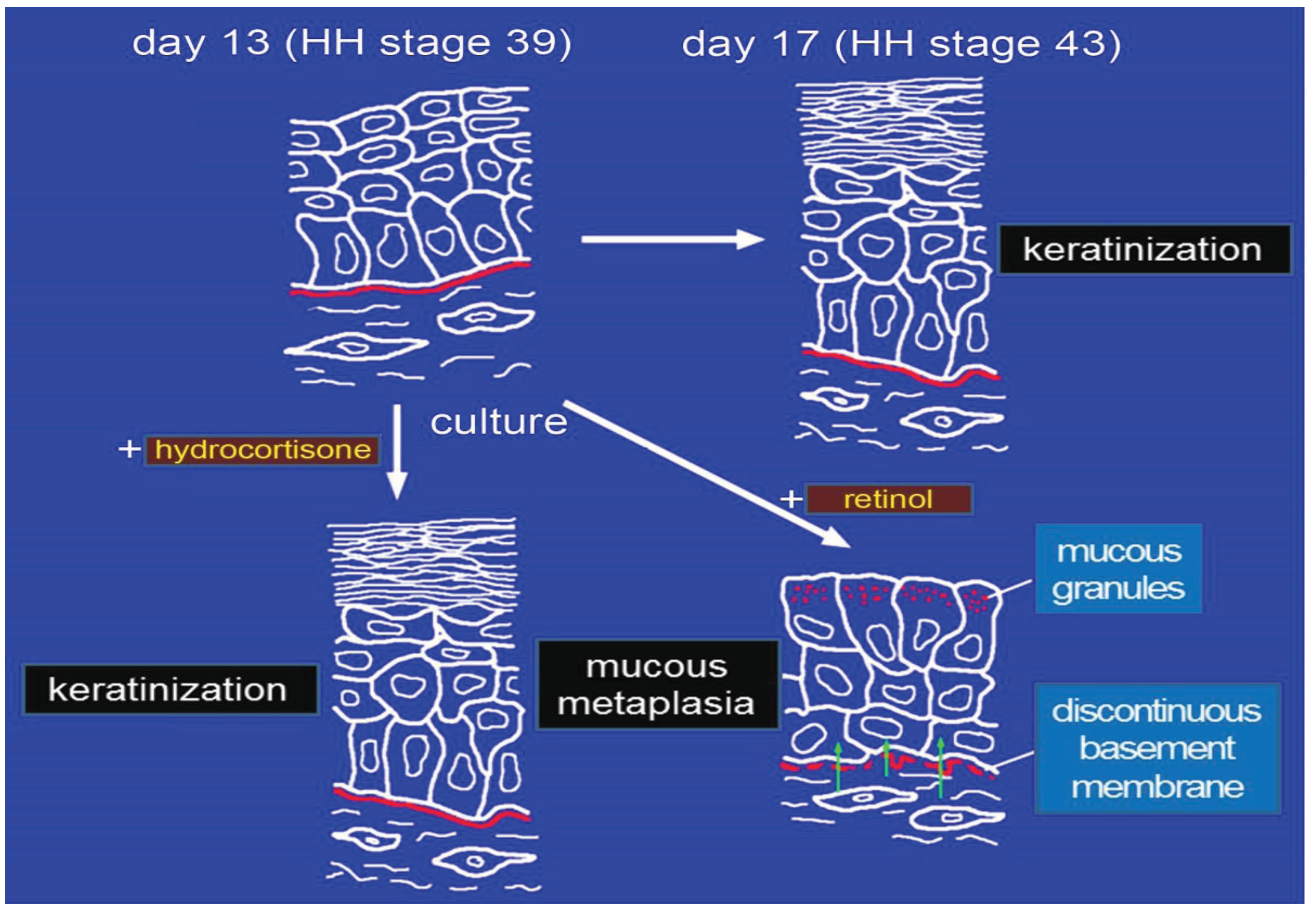
4. Effects of Gbx1 on Chick Skin
5. ATRA Induces Transdifferentiation of Rat Embryonic Epidermis to Mucosal Epithelium with Up-Regulation of Esophageal Markers MUC4 and Keratin 4

{kind=link}
{kind=link}
{kind=link}
| Fold | Gene Symbol | Gene Title | Public ID |
|---|---|---|---|
| 22.07 | Stra6 | stimulated by retinoic acid gene 6 | BI284420 |
| 17.40 | LOC363060 | similar to RIKEN cDNA 1600029D21 | AI599133 |
| 11.37 | Lrat | lecithin-retinol acyltransferase | NM_022280 |
| 9.18 | A2m | alpha-2-macroglobulin | NM_012488 |
| 8.61 | Amy1a | amylase, alpha 1A (salivary) | AB057450 |
| 8.43 | Crisp1 | cysteine-rich secretory protein 1 | NM_022859 |
| 8.04 | Gpr85 | G protein-coupled receptor 85 | AF203907 |
| 7.94 | Cldn7 | claudin 7 | AJ011811 |
| 7.41 | Dhrs3 | dehydrogenase/reductase (SDR family) member 3 | BI276935 |
| 7.12 | Nupl1 | nucleoporin like 1 | AF000901 |
| 7.10 | Dusp14 | dual specificity phosphatase 14 | AI236997 |
| 6.83 | Il6r | interleukin 6 receptor | NM_017020 |
| 6.66 | Ces1d | carboxylesterase 1D | L46791 |
| 6.57 | Tmprss2 | transmembrane protease, serine 2 | AI412136 |
| 6.35 | LOC685158 | similar to CG8138-PA | AI639305 |
| 6.24 | Igfbp5 | insulin-like growth factor binding protein 5 | BE113270 |
| 5.62 | Zfp667 | zinc finger protein 667 | BF402458 |
| 5.51 | Cyp26b1 | cytochrome P450, family 26, subfamily b, polypeptide 1 | BE105541 |
| 5.37 | Tgm2 | transglutaminase 2, C polypeptide | BI275994 |
| 5.27 | Amy2 | amylase 2, pancreatic | NM_031502 |
| 5.17 | Trdn | triadin | AF220558 |
| 5.14 | Sorl1 | sortilin-related receptor, LDLR class A repeats-containing | AI177589 |
| 5.11 | Cyp26a1 | cytochrome P450, family 26, subfamily a, polypeptide 1 | NM_130408 |
| 4.95 | Akap5 | A kinase (PRKA) anchor protein 5 | NM_133515 |
| 4.69 | Mcpt10 | mast cell protease 10 | X68657 |
| 4.59 | Klf2 | Kruppel-like factor 2 (lung) | BF288243 |
| 4.50 | Ush1c | Usher syndrome 1C homolog (human) | BI291932 |
| 4.50 | Cfi | complement factor I | NM_024157 |
| 4.48 | Tpc1808 | tropic 1808 | NM_022625 |
| 4.46 | Car5b | carbonic anhydrase 5b, mitochondrial | AI411132 |
| 4.45 | Scgb1d2 | secretoglobin, family 1D, member 2 | BI285057 |
| 4.37 | Srpx2 | sushi-repeat-containing protein, X-linked 2 | AA818334 |
| 4.37 | Synpr | synaptoporin | BG666364 |
| 4.36 | Slc6a1 | solute carrier family 6 (neurotransmitter transporter, GABA), member 1 | NM_024371 |
| 4.34 | St6gal1 | ST6 beta-galactosamide alpha-2,6-sialyltranferase 1 | M83143 |
| 4.32 | Oprk1 | opioid receptor, kappa 1 | L22536 |
| 4.15 | Mmp11 | matrix metallopeptidase 11 | NM_012980 |
| 4.11 | Dcx | doublecortin | NM_053379 |
| 4.08 | Kalrn | kalirin, RhoGEF kinase | AI639313 |
| 4.05 | Wfdc1 | WAP four-disulfide core domain 1 | BI279661 |
| 4.04 | Scaper | S-phase cyclin A-associated protein in the ER | BF405311 |
| 4.03 | Igfbp6 | insulin-like growth factor binding protein 6 | NM_013104 |
| 3.95 | Prelp | proline/arginine-rich end leucine-rich repeat protein | AI011747 |
| 3.90 | Aoc3 | amine oxidase, copper containing 3 (vascular adhesion protein 1) | AI070137 |
| 3.88 | Tmem176a | transmembrane protein 176A | BM388911 |
| 3.83 | Egr4 | early growth response 4 | NM_019137 |
| 3.75 | Naaa | N-acylethanolamine acid amidase | AI412627 |
| 3.74 | RGD1562533 | similar to mKIAA0774 protein | BI299098 |
| 3.74 | Slc19a1 | solute carrier family 19 (folate transporter), member 1 | NM_017299 |
| 3.69 | Rnf207 | ring finger protein 207 | BF408540 |
| 3.68 | Il17d | Interleukin 17D | AI407169 |
| 3.67 | Pkia | Protein kinase (cAMP-dependent, catalytic) inhibitor alpha | AA996685 |
| 3.66 | Klk8 | kallikrein related-peptidase 8 | BI282567 |
| 3.62 | Map7d1 | MAP7 domain containing 1 | AW253217 |
| 3.62 | Mapk11 | mitogen-activated protein kinase 11 | BF414412 |
| 3.61 | Muc4 | mucin 4, cell surface associated | BM391100 |
6. ATRA Increases Expression of Tgm2 and Gbx1 mRNA and Protein in Rat Embryonic Epidermis
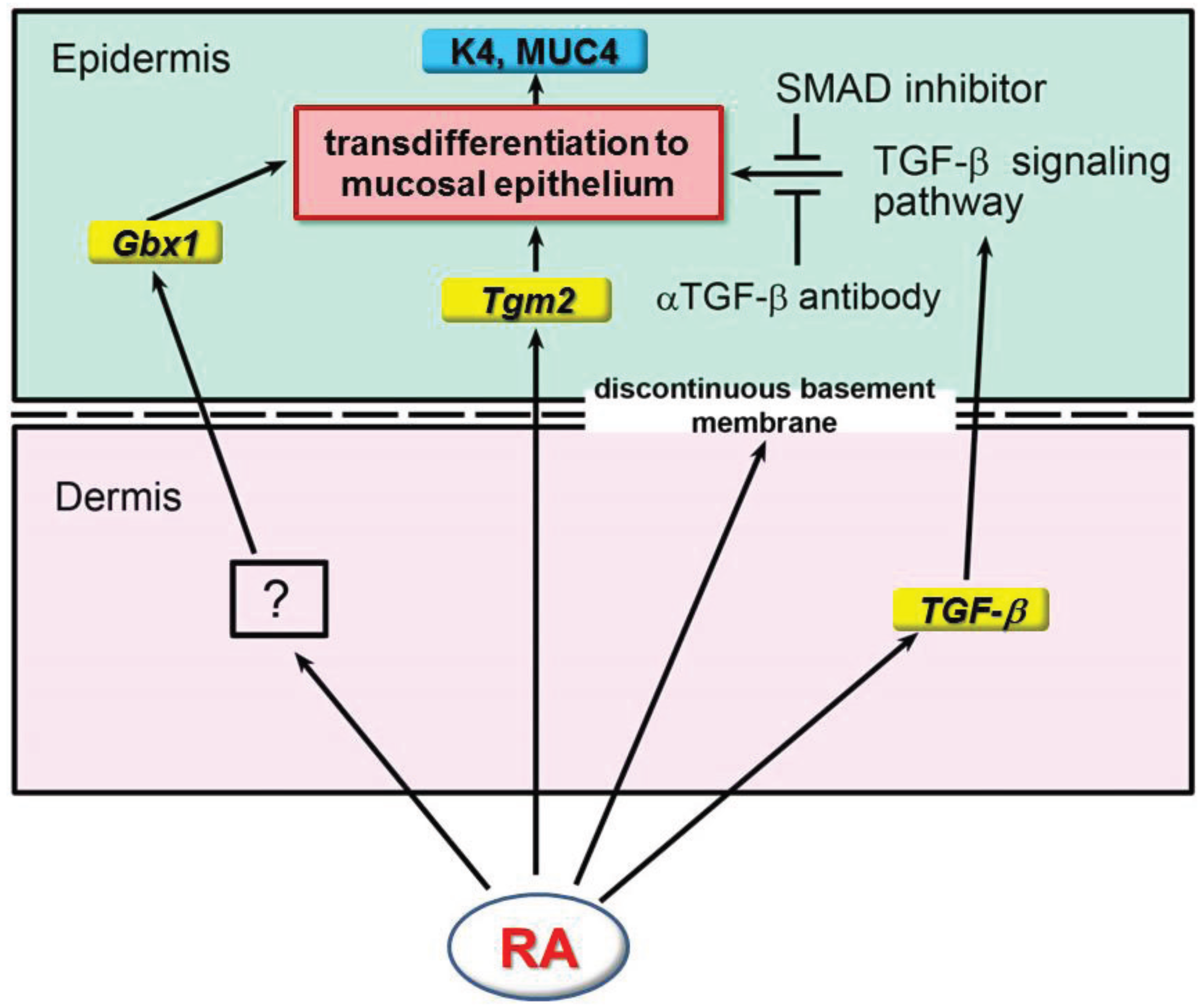
7. Involvement of TGF-β Signaling Pathway in ATRA-Induced Epidermal Transdifferentiation to Mucous Epithelium in Rat Embryonic Skin
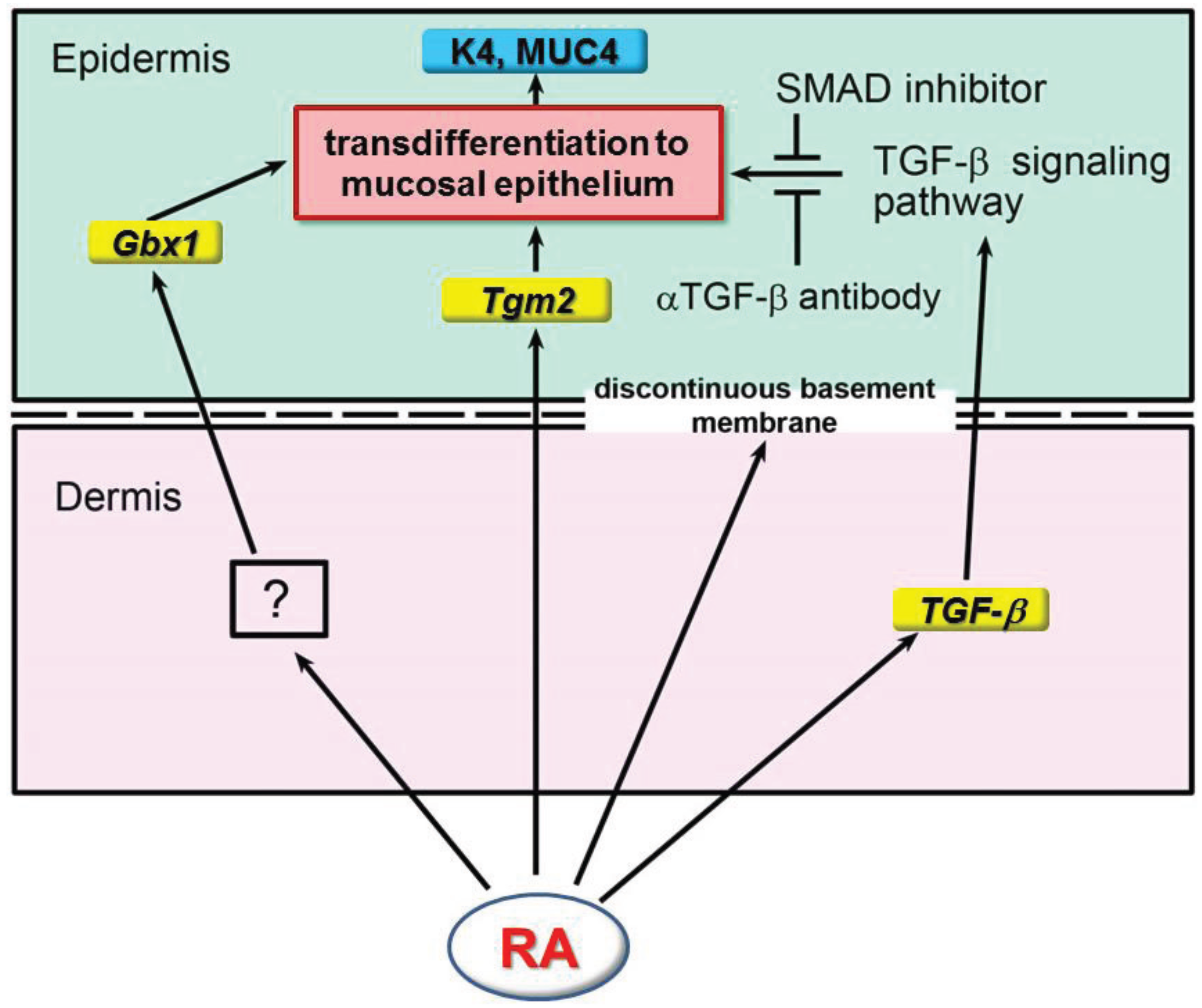
8. Development of Skin in Retinoid Signaling Deficient Mice
9. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Sengel, P. Morphogenesis of Skin; Cambridge University Press: Cambridge, UK, 1976. [Google Scholar]
- Fisher, G.J.; Voorhees, J.J. Molecular mechanisms of retinoid actions in skin. FASEB J. 1996, 10, 1002–1013. [Google Scholar]
- Li, M.; Indra, A.K.; Warot, X.; Brocard, J.; Messaddeq, N.; Kato, S.; Metzger, D.; Chambon, P. Skin abnormalities generated by temporally controlled RXRα mutations in mouse epidermis. Nature 2000, 407, 633–636. [Google Scholar] [CrossRef]
- Chapellier, B.; Mark, M.; Messaddeq, N.; Calléja, C.; Warot, X.; Brocard, J.; Gérard, C.; Li, M.; Metzger, D.; Ghyselinck, N.B.; et al. Physiological and retinoid-induced proliferations of epidermis basal keratinocytes are differently controlled. EMBO J. 2002, 21, 3402–3413. [Google Scholar] [CrossRef]
- Ghyselinck, N.B.; Chapellier, B.; Calléja, C.; Indra, A.K.; Li, M.; Messaddeq, N.; Mark, M.; Metzger, D.; Chambon, P. Genetic dissection of retinoic acid function in epidermis physiology. Ann. Dermatol. Venereol. 2002, 129, 793–799. [Google Scholar]
- García-Fernández, R.A.; Pérez-Martínez, C.; Escudero-Diez, A.; García-Iglesias, M.J. Effects of in utero retinoic acid exposure on mouse pelage hair follicle development. Vet. Dermatol. 2002, 13, 157–163. [Google Scholar] [CrossRef]
- García-Fernández, R.A.; Pérez-Martínez, C.; Alvarez, J.E.; Navarrete, A.J.; García-Iglesias, M.J. Mouse epidermal development: Effects of retinoic acid exposure in utero. Vet. Dermatol. 2006, 17, 36–44. [Google Scholar] [CrossRef]
- Asselineau, D.; Bernard, B.A.; Bailly, C.; Darmon, M. Retinoic acid improves epidermal morphogenesis. Dev. Biol. 1989, 133, 322–335. [Google Scholar] [CrossRef]
- Mori, S. The changes in the para-ocular glands which follow the administration of diets low in fat-soluble vitamin A; with notes on the effect of the same diets on the salivary glands and the mucosa of the larynx and trachea. Bull. Johns Hopkins Hosp. 1922, 33, 357–359. [Google Scholar]
- Wolbach, S.B.; Howe, P.R. Tissue changes following deprivation of fat-soluble vitamin A. J. Exp. Med. 1925, 42, 753–777. [Google Scholar] [CrossRef]
- Fell, H.B.; Mellanby, E. Metaplasia produced in cultures of chick ectoderm by high vitamin A. J. Physiol. 1953, 119, 470–488. [Google Scholar]
- Viallet, J.P.; Ruberte, E.; du Manoir, S.; Krust, A.; Zelent, A.; Dhouailly, D. Retinoic acid-induced glandular metaplasia in mouse skin is linked to the dermal expression of retinoic acid receptor beta mRNA. Dev. Biol. 1991, 144, 424–428. [Google Scholar] [CrossRef]
- Viallet, J.P.; Dhouailly, D. Retinoic acid and mouse skin morphogenesis. II. Role of epidermal competence in hair glandular metaplasia. Dev. Biol. 1994, 166, 277–288. [Google Scholar] [CrossRef]
- Chuong, C.-M.; Ting, S.A.; Widelitz, R.B.; Lee, Y.-S. Mechanism of skin morphogenesis. II. Retinoic acid modulates axis orientation and phenotypes of skin appendages. Development 1992, 115, 839–852. [Google Scholar]
- Dhouailly, D. Genetic expression and morphogenesis of the skin in vertebrates. Ann. Genet. 1993, 36, 47–55. [Google Scholar]
- Gudas, L.J. Retinoids and vertebrate development. J. Biol. Chem. 1994, 269, 15399–15402. [Google Scholar]
- Cardoso, W.V.; Mitsialis, S.A.; Brody, J.S.; Williams, M.C. Retinoic acid alters the expression of pattern-related genes in the developing rat lung. Dev. Dyn. 1996, 207, 47–59. [Google Scholar] [CrossRef]
- Kanzler, B.; Prin, F.; Thelu, J.; Dhouailly, D. CHOXC-8 and CHOXD-13 expression in embryonic chick skin and cutaneous appendage specification. Dev. Dyn. 1997, 210, 274–287. [Google Scholar] [CrossRef]
- Noveen, A.; Jiang, T.X.; Ting-Berreth, S.A.; Chuong, C.M. Homeobox genes Msx-1 and Msx-2 are associated with induction and growth of skin appendages. J. Invest. Dermatol. 1995, 104, 711–719. [Google Scholar]
- Lin, C.M.; Jiang, T.X.; Widelitz, R.B.; Chuong, C.M. Molecular signaling in feather morphogenesis. Curr. Opin. Cell Biol. 2006, 18, 730–741. [Google Scholar] [CrossRef]
- Obinata, A.; Akimoto, Y.; Omoto, Y.; Hirano, H. Increase in expression of the homeobox gene, Gbx1, in retinol-induced epidermal mucous metaplasia. Biochem. Biophys. Res. Commun. 2001, 280, 1055–1061. [Google Scholar] [CrossRef]
- Obinata, A.; Osakabe, K.; Yamaguchi, M.; Morimoto, R.; Akimoto, Y. Tgm2/Gh, Gbx1 and TGF-β are involved in retinoic acid-induced transdifferentiation from epidermis to mucosal epithelium. Int. J. Dev. Biol. 2011, 55, 933–943. [Google Scholar] [CrossRef]
- Imakado, S.; Bickenbach, J.R.; Bundman, D.S.; Rothnagel, J.A.; Attar, P.S.; Wang, X.J.; Walczak, V.R.; Wisniewski, S.; Pote, J.; Gordon, J.S.; Heyman, R.A.; et al. Targeting expression of a dominant-negative retinoic acid receptor mutant in the epidermis of transgenic mice results in loss of barrier function. Genes Dev. 1995, 9, 317–329. [Google Scholar] [CrossRef]
- Attar, P.S.; Wertz, P.W.; McArthur, M.; Imakado, S.; Bickenbach, J.R.; Roop, D.R. Inhibition of retinoid signaling in transgenic mice alters lipid processing and disrupts epidermal barrier function. Mol. Endocrinol. 1997, 11, 792–800. [Google Scholar] [CrossRef]
- Calléja, C.; Messaddeq, N.; Chapellier, B.; Yang, H.; Krezel, W.; Li, M.; Metzger, D.; Mascrez, B.; Ohta, K.; Kagechika, H.; et al. Genetic and pharmacological evidence that a retinoic acid cannot be the RXR-activating ligand in mouse epidermis keratinocytes. Genes Dev. 2006, 20, 1525–1538. [Google Scholar] [CrossRef]
- Okano, J.; Lichti, U.; Mamiya, S.; Aronova, M.; Zhang, G.; Yuspa, S.H.; Hamada, H.; Skai, Y,; Morasso, M.I. Increased retinoic acid levels through ablation of Cyp26b1 determine the processes of embryonic skin barrier formation and peridermal development. J. Cell Sci. 2012, 125, 1827–1836. [Google Scholar] [CrossRef]
- Okano, J.; Levy, C.; Lichti, U.; Sun, H.W.; Yuspa, S.H.; Sakai, Y.; Morasso, M.I. Cutaneous retinoic acid levels determine hair follicle development and downgrowth. J. Biol. Chem. 2012, 287, 39304–39315. [Google Scholar]
- Duester, G. Retinoic acid synthesis and signaling during early organogenesis. Cell 2008, 134, 921–931. [Google Scholar] [CrossRef]
- Dhouailly, D.; Hardy, M.H.; Sengel, P. Formation of feathers on chick foot scales: A stage-dependent morphogenetic response to retinoic acid. J. Embryol. Exp. Morphol. 1980, 58, 63–78. [Google Scholar]
- Cadi, R.; Dhouailly, D.; Sengel, P. Use of retinoic acid for the analysis of dermal-epidermal interactions in the tarsometatarsal skin of the chick embryo. Dev. Biol. 1983, 100, 489–495. [Google Scholar] [CrossRef]
- Prin, F.; Dhouailly, D. How and when the regional competence of chick epidermis is established: feathers vs. scutate and reticulate scales, a problem en route to a solution. Int. J. Dev. Biol. 2004, 48, 137–148. [Google Scholar] [CrossRef]
- Obinata, A.; Kawada, M.; Endo, H. Induction of epidermal mucous metaplasia by culture of recombinants of undifferentiated epidermis and retinol-treated dermis in a chemically defined medium. Dev. Biol. 1987, 123, 59–62. [Google Scholar] [CrossRef]
- Obinata, A.; Akimoto, Y.; Hirano, H.; Endo, H. Stimulation by Bt2cAMP of epidermal mucous metaplasia in retinol-pretreated chick embryonic cultured skin, and its inhibition by herbimycin A, an inhibitor for protein tyrosine kinase. Exp. Cell Res. 1991, 193, 36–44. [Google Scholar] [CrossRef]
- Obinata, A.; Akimoto, Y.; Hirano, H.; Endo, H. Short-term retinol treatment in vitro induces stable transdifferentiation of chick epidermal cells into mucus-secreting cells. Roux’s Arch. Dev. Biol. 1991, 200, 289–295. [Google Scholar] [CrossRef]
- Obinata, A.; Akimoto, Y.; Kawamata, T.; Hirano, H. Induction of mucous metaplasia in chick embryonic skin by retinol-pretreated embryonic chick or quail dermal fibroblasts through cell-cell interaction: Correlation of a transient increase in retinoic acid receptor β mRNA in retinol-treated dermal fibroblasts with their competence to induce epidermal mucous metaplasia. Dev. Growth Differ. 1994, 36, 579–587. [Google Scholar]
- Obinata, A.; Akimoto, Y. Effects of retinoic acid and Gbx1 on feather-bud formation and epidermal transdifferentiation in chick embryonic cultured dorsal skin. Dev. Dyn. 2012, 241, 1405–1412. [Google Scholar]
- Gehring, W.J.; Affolter, M.; Bürglin, T. Homeodomain proteins. Annu. Rev. Biochem. 1994, 63, 487–526. [Google Scholar] [CrossRef]
- Obinata, A.; Akimoto, Y.; Omoto, Y.; Hirano, H. Expression of Hex homeobox gene during skin development: Increase in epidermal cell proliferation by transfecting the Hex to the dermis. Dev. Growth Differ. 2002, 44, 281–292. [Google Scholar]
- Obinata, A.; Akimoto, Y. Expression of Hex during feather bud development. Int. J. Dev. Biol. 2005, 49, 885–890. [Google Scholar] [CrossRef]
- Obinata, A.; Akimoto, Y. Involvement of Hex in the initiation of feather morphogenesis. Int. J. Dev. Biol. 2005, 49, 953–960. [Google Scholar] [CrossRef]
- Kosaka, Y.; Akimoto, Y.; Omoto, Y.; Obinata, A.; Hirano, H. Expression of the HB9 homeobox gene concomitant with proliferation accompanying epidermal stratification during development of chick embryonic tarsometatarsal skin. Histochem. J. 2000, 32, 275–280. [Google Scholar] [CrossRef]
- Kosaka, Y.; Akimoto, Y.; Obinata, A.; Hirano, H. Localization of HB9 homeobox gene mRNA and protein during the early stages of chick feather development. Biochem. Biophys. Res. Commun. 2000, 276, 1112–1117. [Google Scholar] [CrossRef]
- Niss, K.; Leutz, A. Expression of the homeobox gene GBX2 during chicken development. Mech. Dev. 1998, 76, 151–155. [Google Scholar] [CrossRef]
- Reid, A.I.; Gaunt, S.J. Colinearity and non-colinearity in the expression of Hox genes in developing chick skin. Int. J. Dev. Biol. 2002, 46, 209–215. [Google Scholar]
- Rouzankina, I.; Abate-Shen, C.; Niswander, L. Dlx genes integrate positive and negative signals during feather bud development. Dev. Biol. 2004, 265, 219–233. [Google Scholar]
- Asbreuk, C.H.J.; van Schaick, H.S.A.; Cox, J.J.; Kromkamp, M.; Smidt, M.P.; Burbach, J.P.H. The homeobox genes Lhx7 and Gbx1 are expressed in the basal forebrain cholinergic system. Neuroscience 2002, 109, 287–298. [Google Scholar] [CrossRef]
- Waters, S.T.; Wilson, C.P.; Lewandoski, M. Cloning and embryonic expression analysis of the mouse Gbx1 gene. Gene Expr. Patterns 2003, 3, 313–317. [Google Scholar] [CrossRef]
- Rhinn, M.; Lun, K.; Werner, M.; Simeone, A.; Brand, M. Isolation and expression of the homeobox gene Gbx1 during mouse development. Dev. Dyn. 2004, 229, 334–339. [Google Scholar] [CrossRef]
- Widelitz, R.B.; Jiang, T.X.; Noveen, A.; Chen, C.W.; Chuong, C.M. FGF induces new feather buds from developing avian skin. J. Invest. Dermatol. 1996, 107, 797–803. [Google Scholar]
- Noramly, S.; Morgan, B.A. BMPs mediate lateral inhibition at successive stages in feather tract development. Development 1998, 125, 3775–3787. [Google Scholar]
- Bitgood, M.J.; McMahon, A.P. Hedgehog and Bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev. Biol. 1995, 172, 126–138. [Google Scholar] [CrossRef]
- Noveen, A.; Jiang, T.X.; Chuong, C.M. cAMP, an activator of protein kinase A, suppresses the expression of sonic hedgehog. Biochem. Biophys. Res. Commun. 1996, 219, 180–185. [Google Scholar] [CrossRef]
- Ting-Berreth, S.A.; Chuong, C.M. Sonic Hedgehog in feather morphogenesis: Induction of mesenchymal condensation and association with cell death. Dev. Dyn. 1996, 207, 157–170. [Google Scholar] [CrossRef]
- Widelitz, R.B.; Jiang, T.X.; Chen, C.W.; Stott, N.S.; Jung, H.S.; Chuong, C.M. Wnt-7a in feather morphogenesis: Involvement of anterior-posterior asymmetry and proximal-distal elongation demonstrated with an in vitro reconstitution model. Development 1999, 126, 2577–2587. [Google Scholar]
- Chen, C.W.; Jung, H.S.; Jiang, T.X.; Chuong, C.M. Asymmetric expression of Notch/Delta/Serrate is associated with the anterior-posterior axis of feather buds. Dev. Biol. 1997, 188, 181–187. [Google Scholar] [CrossRef]
- Noramly, S.; Freeman, A.; Morgan, B.A. β-catenin signaling can initiate feather bud development. Development 1999, 126, 3509–3521. [Google Scholar]
- Takata, K.; Obinata, A.; Endo, H.; Hirano, H. Induction of the α-type keratinization by hydrocortisone in embryonic chick skins grown in a chemically defined medium. An electron microscopic study. Dev. Biol. 1981, 85, 370–379. [Google Scholar] [CrossRef]
- Ness, S.L.; Edelmann, W.; Jenkins, T.D.; Liedtke, W.; Rustgi, A.K.; Kucherlapati, R. Mouse keratin 4 is necessary for internal epithelial integrity. J. Biol. Chem. 1998, 273, 23904–23911. [Google Scholar] [CrossRef]
- Guillem, P.; Billeret, V.; Buisine, M.-P.; Flejou, J.-F.; Lecomte-Houcke, M.; Degand, P.; Aubert, J.-P.; Triboulet, J.-P.; Porchet, N. Mucin gene expression and cell differentiation in human normal, premalignant and malignant esophagus. Int. J. Cancer 2000, 88, 856–861. [Google Scholar] [CrossRef]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2009, 214, 516–559. [Google Scholar] [CrossRef]
- Lichti, U.; Yuspa, S.H. Inhibition of epidermal terminal differentiation and tumour promotion by retinoids. Ciba Found. Symp. 1985, 113, 77–89. [Google Scholar]
- Foitzik, K.; Spexard, T.; Nakamura, M.; Halsner, U.; Paus, R. Towards dissecting the pathogenesis of retinoid-induced hair loss: All-trans retinoic acid induces premature hair follicle regression (catagen) by upregulation of transforming growth factor-β2 in the dermal papilla. J. Invest. Dermatol. 2005, 124, 1119–1126. [Google Scholar]
- Fesus, L.; Piacentini, M. Transglutaminase 2: An enigmatic enzyme with diverse functions. Trends Biochem. Sci. 2002, 27, 534–539. [Google Scholar] [CrossRef]
- Nakaoka, H.; Perez, D.M.; Baek, K.J.; Das, T.; Husain, A.; Misono, K.; Im, M.-J.; Graham, R.M. Gh: A GTP-binding protein with transglutaminase activity and receptor signaling function. Science 1994, 264, 1593–1596. [Google Scholar]
- Mishra, S.; Murphy, L.J. Tissue transglutaminase has intrinsic kinase activity: Identification of transglutaminase 2 as an insulin-like growth factor-binding protein-3 kinase. J. Biol. Chem. 2004, 279, 23863–23868. [Google Scholar] [CrossRef]
- Tucholski, J.; Johnson, G.V.W. Tissue transglutaminase directly regulates adenylyl cyclase resulting in enhanced cAMP-response element-binding protein (CREB) activation. J. Biol. Chem. 2003, 278, 26838–26843. [Google Scholar] [CrossRef]
- Lai, T.S.; Liu, Y.; Tucker, T.; Daniel, K.R.; Sane, D.C.; Toone, E.; Burke, J.R.; Strittmatter, W.J.; Greenberg, C.S. Identification of chemical inhibitors to human tissue transglutaminase by screening existing drug libraries. Chem. Biol. 2008, 15, 969–978. [Google Scholar] [CrossRef]
- Singh, U.S.; Pan, J.; Kao, Y.-L.; Joshi, S.; Young, K.L.; Baker, K.M. Tissue transglutaminase mediates activation of RhoA and MAP kinase pathways during retinoic acid-induced differentiation of SH-SY5Y cells. J. Biol. Chem. 2003, 278, 391–399. [Google Scholar]
- Ou, H.; Haendeler, J.; Aebly, M.R.; Kelly, L.A.; Cholewa, B.C.; Koike, J.; Kwitek-Black, A.; Jacob, H.J.; Berk, B.C.; Miano, J.M. Retinoic acid-induced tissue transglutaminase and apoptosis in vascular smooth muscle cells. Circ. Res. 2000, 87, 881–887. [Google Scholar] [CrossRef]
- Antonyak, M.A.; Singh, U.S.; Lee, D.A.; Boehm, J.E.; Combs, C.; Zgola, M.M.; Page, R.L.; Cerione, R.A. Effects of tissue transglutaminase on retinoic acid-induced cellular differentiation and protection against apoptosis. J. Biol. Chem. 2001, 276, 33582–33587. [Google Scholar]
- Lichti, U.; Ben, T.; Yuspa, S.H. Retinoic acid-induced transglutaminase in mouse epidermal cells is distinct from epidermal transglutaminase. J. Biol. Chem. 1985, 260, 1422–1426. [Google Scholar]
- Nanda, N.; Iismaa, S.E.; Owens, W.A.; Husain, A.; Mackay, F.; Graham, R.M. Targeted inactivation of Gh/tissue transglutaminase II. J. Biol. Chem. 2001, 276, 20673–20678. [Google Scholar]
- Lorand, L.; Graham, R.M. Transglutaminases crosslinking enzymes with pleiotropic functions. Nat. Rev. Mol. Cell Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef]
- Dalby, K.N.; Takedereli, I.; Lopez-Berestein, G.; Ozpolat, B. Targeting the prodeath and prosurvival functions of autophagy as novel therapeutic strategies in cancer. Autophagy 2010, 6, 322–329. [Google Scholar] [CrossRef]
- Telci, D.; Griffin, M. Tissue transglutaminase (TG2)—A wound response enzyme. Front. Biosci. 2006, 11, 867–882. [Google Scholar] [CrossRef]
- Steinert, P.M.; Kim, S.Y.; Chung, S.I.; Marecov, L.N. The transglutaminase 1 enzyme is variably acylated by myristate and palmitate during differentiation in epidermal keratinocytes. J. Biol. Chem. 1996, 271, 26242–26250. [Google Scholar]
- Zhang, J.; Zhi, H.Y.; Ding, F.; Luo, A.P.; Liu, Z.H. Transglutaminase3 expression in C57BL/6J mouse embryo epidermis and the correlation with its differentiation. Cell Res. 2005, 15, 105–110. [Google Scholar] [CrossRef]
- Candi, E.; Oddi, S.; Paradisi, A.; Terrinoni, A.; Ranalli, M.; Teofoli, P.; Citro, G.; Scarpato, S.; Puddu, P.; Melino, G. Expression of transglutaminase 5 in normal and pathologic human epidermis. J. Invest. Dermatol. 2002, 119, 670–677. [Google Scholar] [CrossRef]
- Inman, G.J.; Nicolás, F.J.; Callahan, J.F.; Harling, J.D.; Gaster, L.M.; Reith, A.D.; Laping, N.J.; Hill, C.S. SB-431542 is a potent and specific inhibitor of transforming growth factor-β superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol. Pharmacol. 2002, 62, 65–74. [Google Scholar] [CrossRef]
- Everts, H.B. Endogenous retinoids in the hair follicle and sebaceous gland. Biochim. Biophys. Acta 2012, 1821, 222–229. [Google Scholar]
- Everts, H.B.; Silva, K.A.; Montgomery, S.; Suo, L.; Menser, M.; Valet, A.S.; King, L.E.; Ong, D.E.; Sundberg, J.P. Retinoid metabolism is altered in human and mouse cicatricial alopecia. J. Invest. Dermatol. 2013, 133, 325–333. [Google Scholar] [CrossRef]
- Duncan, F.J.; Silva, K.A.; Johnson, C.J.; King, B.L.; Szatkiewicz, J.P.; Kamdar, S.P.; Ong, D.E.; Napoli, J.L.; Wang, J.; King, L.E., Jr.; et al. Endogenous retinoids in the pathogenesis of alopecia areata. J. Invest. Dermatol. 2013, 133, 334–343. [Google Scholar] [CrossRef]
- Saitou, M.; Sugai, S.; Tanaka, T.; Shimouchi, K.; Fuchs, E.; Narumiya, S.; Kakizuka, A. Inhibition of skin development by targeted expression of a dominant-negative retinoic acid receptor. Nature 1995, 374, 159–162. [Google Scholar] [CrossRef]
- Reichrath, J.; Mittmann, M.; Kamradt, J.; Muller, S.M. Expression of retinoid-X receptors (-α, -β, -γ) and retinoic acid receptors (-α, -β, -γ) in normal human skin: An immunohistological evaluation. Histochem. J. 1997, 29, 127–133. [Google Scholar] [CrossRef]
- Li, M.; Chiba, H.; Warot, X.; Messaddeq, N.; Gérard, C.; Chambon, P.; Metzger, D. RXRα ablation in skin keratinocytes results in alopecia and epidermal alterations. Development 2001, 128, 675–688. [Google Scholar]
- Chen, C.F.; Lohnes, D. Dominant-negative retinoic acid receptors elicit epidermal defects through a non-canonical pathway. J. Biol. Chem. 2005, 280, 3012–3021. [Google Scholar] [CrossRef]
- Blanchet, S.; Favier, B.; Chevalier, G.; Kastner, P.; Michaille, J.J.; Chambon, P.; Dhouailly, D. Both retinoic acid receptors α (RARα) and γ (RARγ) are able to initiate mouse upper-lip skin glandular metaplasia. J. Invest Dermatol. 1998, 111, 206–212. [Google Scholar] [CrossRef]
- Shih, M.Y.; Kane, M.A.; Zhou, P.; Yen, C.L.; Streeper, R.S.; Napoli, J.L.; Farese, R.V., Jr. Retinol esterification by DGAT1 is essential for retinoid homeostasis in murine skin. J. Biol. Chem. 2009, 284, 4292–4299. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Akimoto, Y.; Miyaji, M.; Morimoto-Kamata, R.; Kosaka, Y.; Obinata, A. Retinoic Acid-Induced Epidermal Transdifferentiation in Skin. J. Dev. Biol. 2014, 2, 158-173. https://doi.org/10.3390/jdb2030158
Akimoto Y, Miyaji M, Morimoto-Kamata R, Kosaka Y, Obinata A. Retinoic Acid-Induced Epidermal Transdifferentiation in Skin. Journal of Developmental Biology. 2014; 2(3):158-173. https://doi.org/10.3390/jdb2030158
Chicago/Turabian StyleAkimoto, Yoshihiro, Mary Miyaji, Riyo Morimoto-Kamata, Yasuhiro Kosaka, and Akiko Obinata. 2014. "Retinoic Acid-Induced Epidermal Transdifferentiation in Skin" Journal of Developmental Biology 2, no. 3: 158-173. https://doi.org/10.3390/jdb2030158
APA StyleAkimoto, Y., Miyaji, M., Morimoto-Kamata, R., Kosaka, Y., & Obinata, A. (2014). Retinoic Acid-Induced Epidermal Transdifferentiation in Skin. Journal of Developmental Biology, 2(3), 158-173. https://doi.org/10.3390/jdb2030158