
Activation of Marck-like Genes and Proteins During Initial Phases of Regeneration in the Amputated Tail and Limb of the Lizard Podarcis muralis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Tissue Preparation
2.2. Microscopical Methods
3. Results
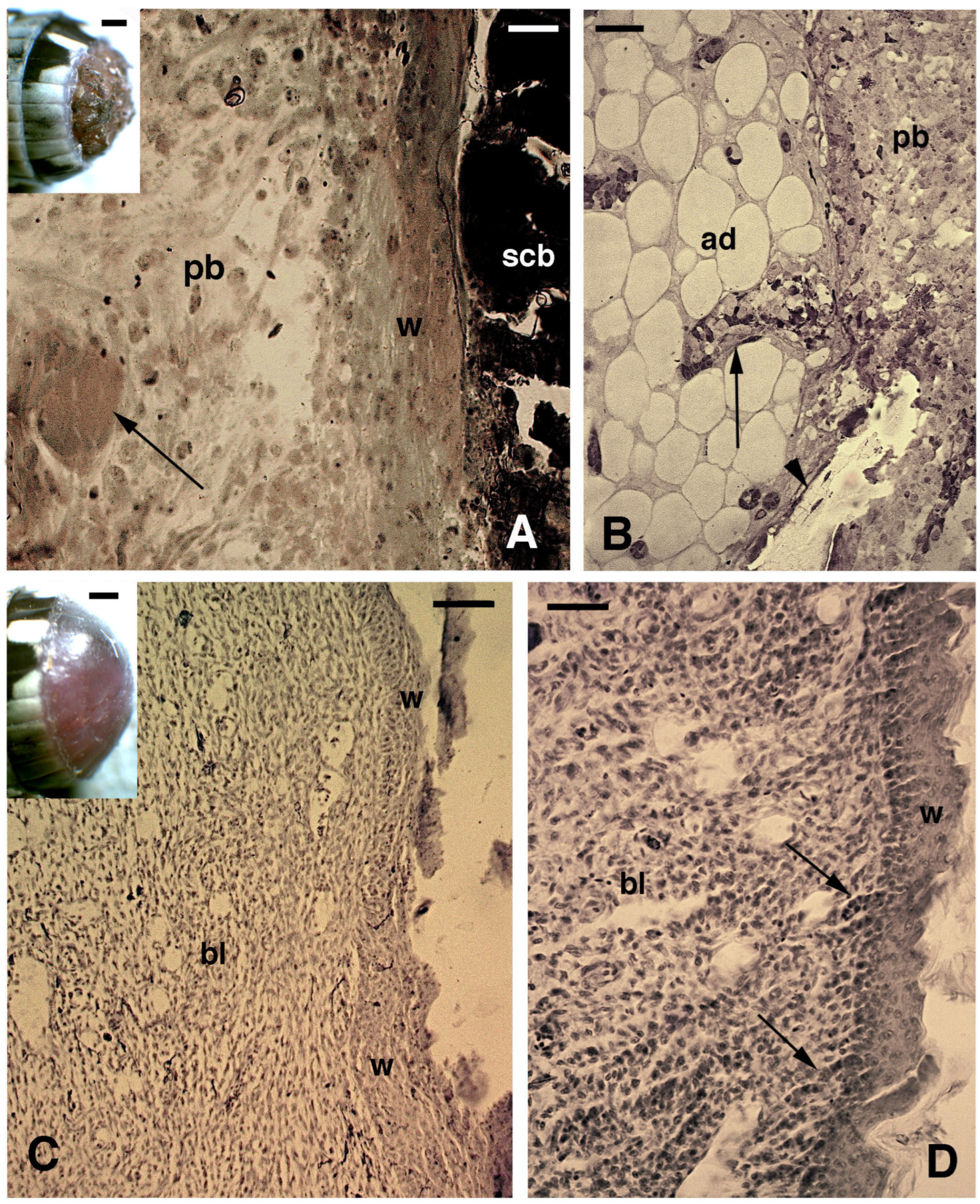
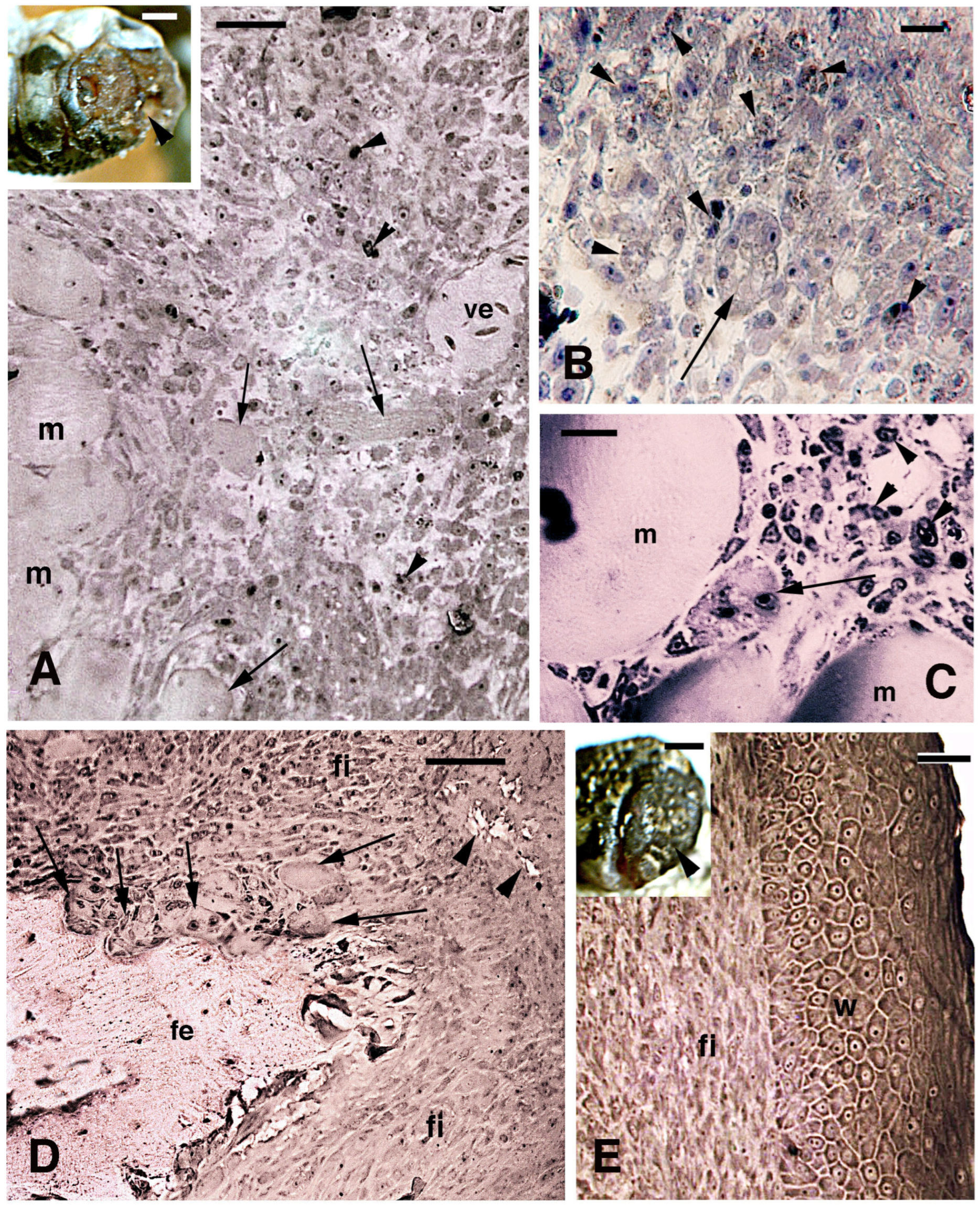
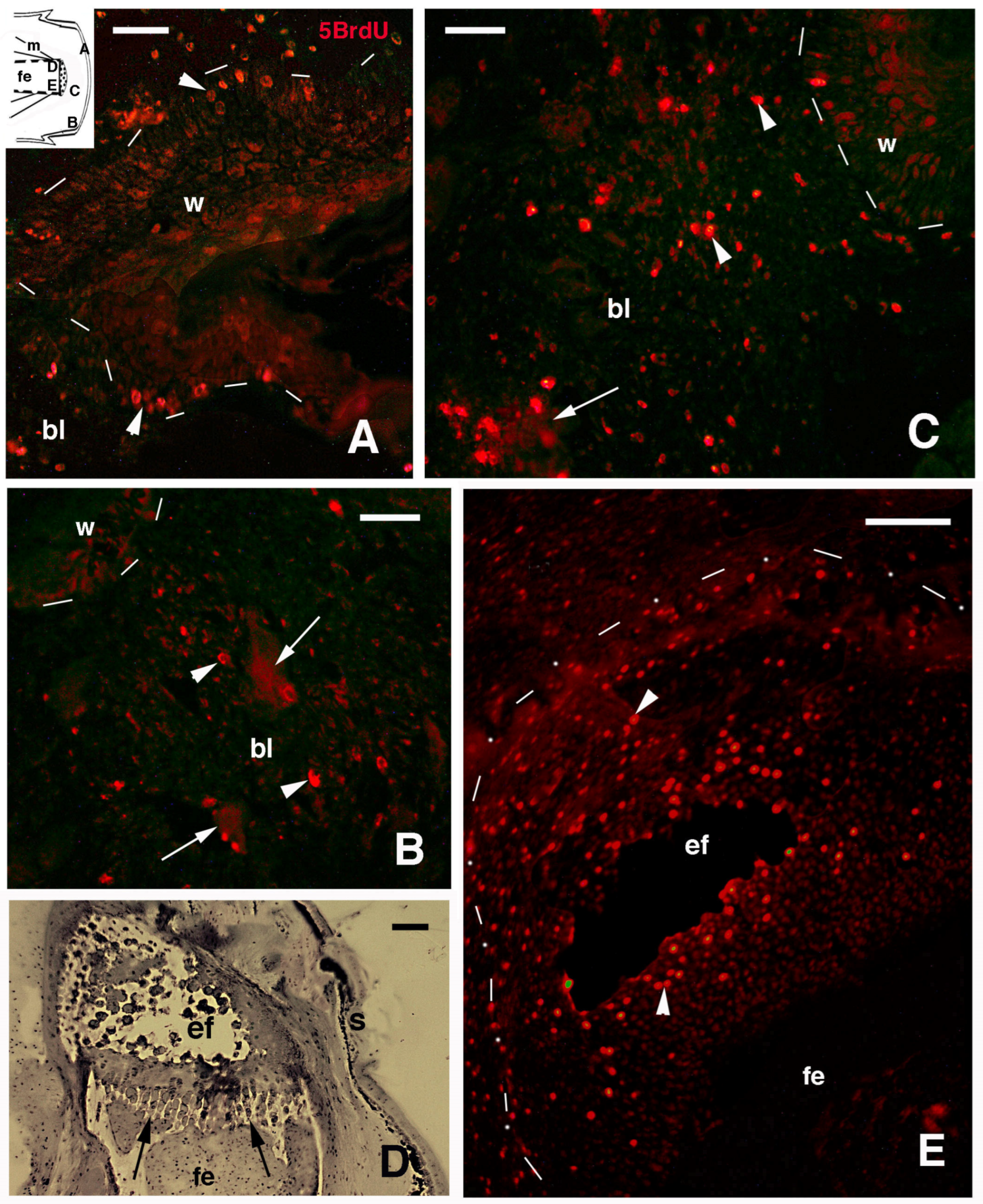
3.1. Histology and 5BrdU-Immunolabeling in the Tail and Limb Initial Blastemas
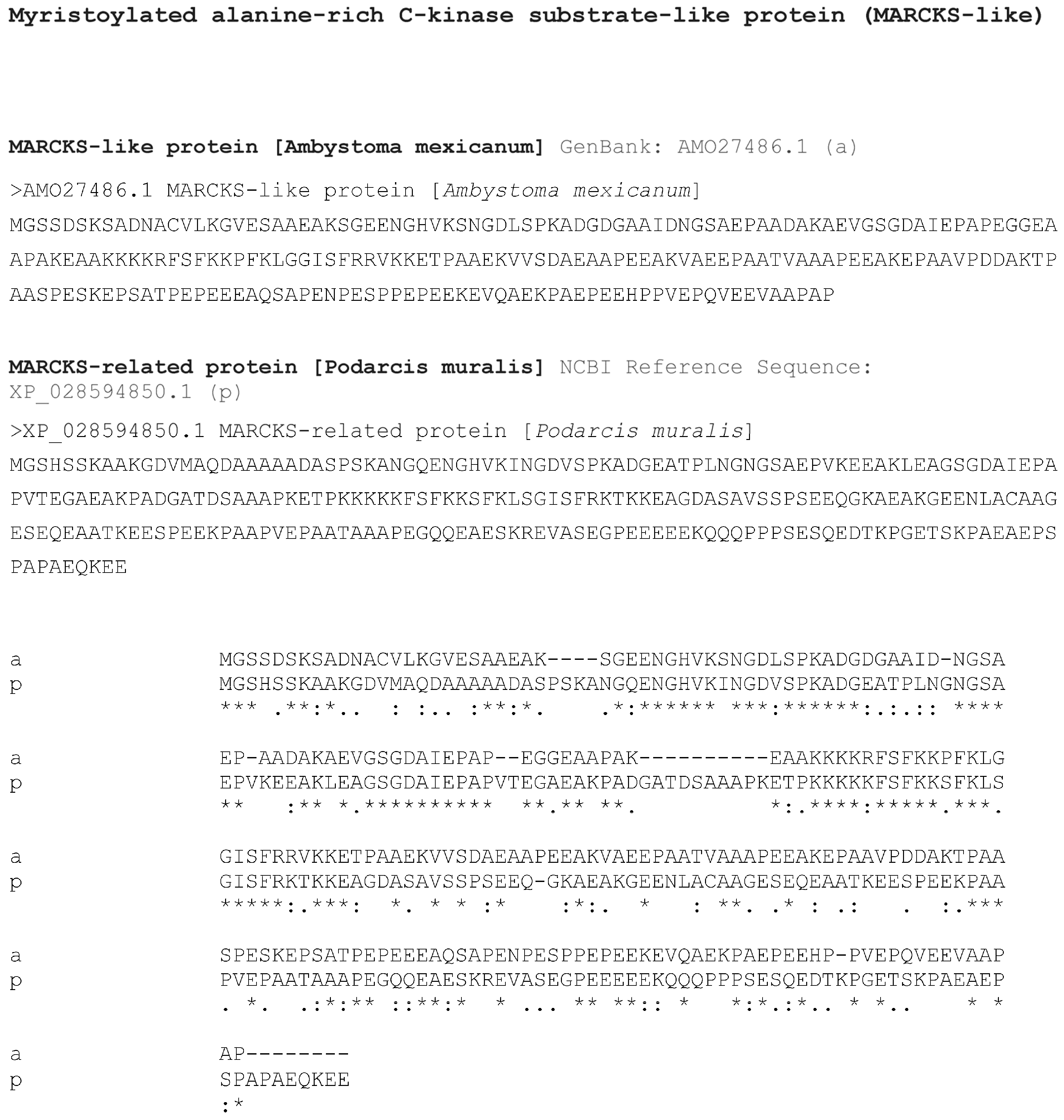
3.2. Bioinformatics Analysis
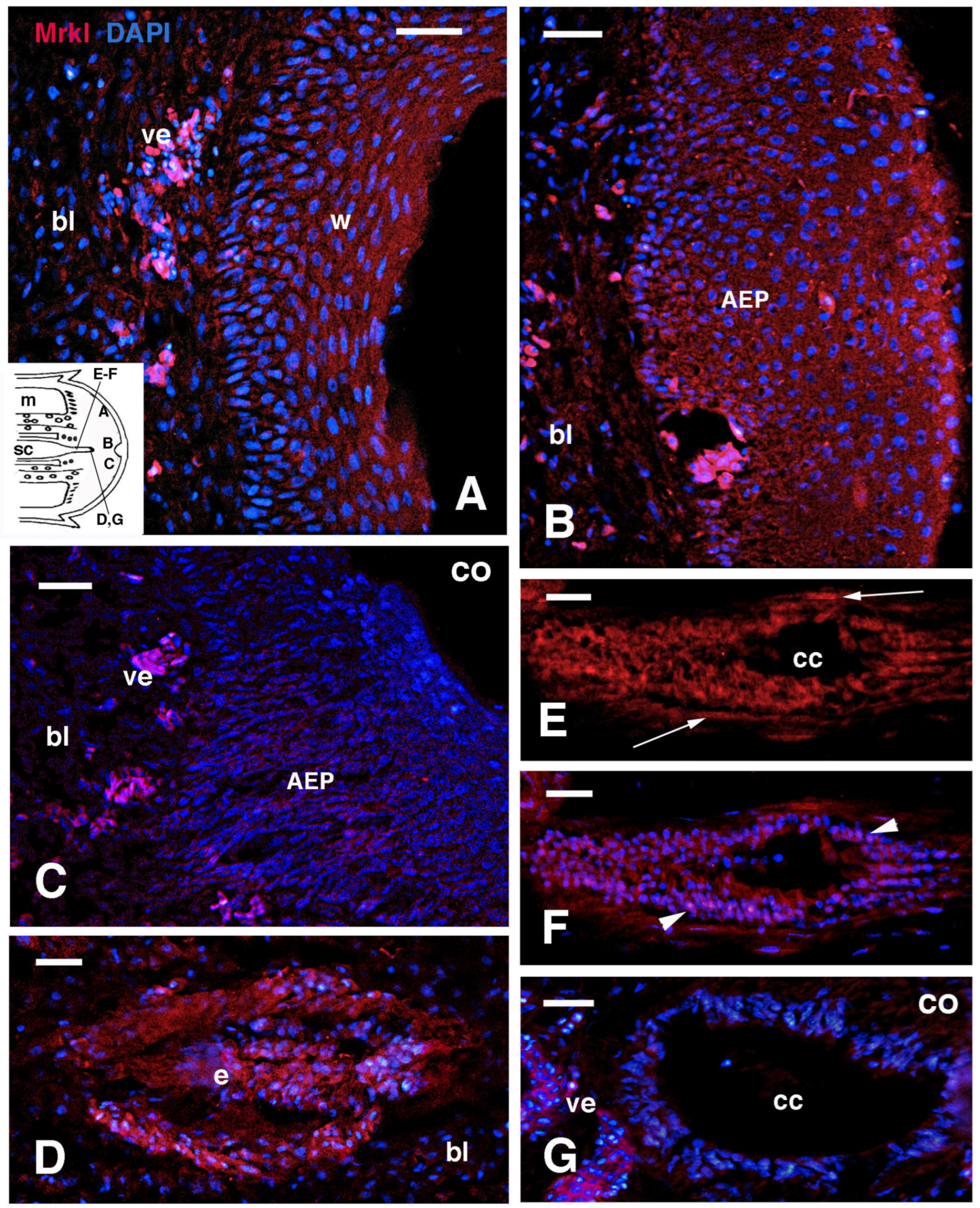
3.3. MARCK-like Immunolocalization
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reichman, O.J. Evolution of regenerative capabilities. Amer. Natur. 1984, 123, 752–763. [Google Scholar] [CrossRef]
- Carlson, B.M. Principles of Regenerative Biology; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Cox, P.G. Some aspects of tail regeneration in the lizard, Anolis carolinensis. I. A description based on histology and autoradiography. J. Exp. Zool. 1969, 171, 127–150. [Google Scholar] [CrossRef]
- Bellairs, D.A.; Bryant, S.V. Autotomy and regeneration in reptiles. In Biology of the Reptilia; Billet, G.C.F., Maderson, P.F.A., Eds.; John Wiley & Sons: New York, NY, USA, 1985; Volume 15B, pp. 302–410. [Google Scholar]
- Alibardi, L. Ultrastructural features of the process of wound healing after tail and limb amputation in lizard. Acta Zool. 2010, 91, 306–318. [Google Scholar] [CrossRef]
- Fisher, R.E.; Geiger, L.A.; Stroik, L.K.; Hutchins, E.D.; George, R.M.; DeNardo, D.F.; Kosumi, K.; Rawls, J.A.; Wilson-Rawls, J. A histological comparison of the original and regenerated tail in the green anole, Anolis carolinensis. Anat. Rec. 2012, 295, 1609–1619. [Google Scholar] [CrossRef]
- Gilbert, E.A.B.; Payne, S.L.; Vickaryous, M.K. The anatomy and histology of caudal autotomy and regeneration in lizards. Physiol. Bioch. Zool. 2013, 86, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Lozito, T.P.; Tuan, R.S. Lizard tail regeneration as an instructive model of enhanced healing capabilities in an adult amniote. Connect. Tiss. Res. 2016, 58, 145–154. [Google Scholar] [CrossRef]
- Jacyniak, K.; McDonald, R.P.; Vickaryous, M.K. Tail regeneration and other phenomena of wound healing and tissue restoration in lizards. J. Exp. Biol. 2017, 220, 2858–2869. [Google Scholar] [CrossRef]
- Londono, R.; Wenzhong, W.; Wang, B.; Tuan, R.S.; Lozito, T.P. Cartilage and muscle cell fate and origin during lizard tail regeneration. Front. Bioeng. Biotechnol. 2017, 5, 70. [Google Scholar] [CrossRef]
- Hutchins, E.D.; Markov, G.J.; Eckalbar, W.L.; Gorge, R.M.; King, J.M.; Tokuyama, M.A.; Geiger, L.A.; Emmert, N.; Ammar, M.J.; Allen, A.P.; et al. Transcriptomic analysis of tail regeneration in the lizard Anolis carolinensis reveals activation of conserved vertebrate developmental and repair mechanisms. PLoS ONE 2014, 9, e105004. [Google Scholar] [CrossRef]
- Vitulo, N.; Dalla Valle, L.; Skobo, T.; Valle, G.; Alibardi, L. Transcriptome analysis of the regenerating tail versus the scarring limb in lizard reveals pathways leading to successful versus unsuccessful organ regeneration in amniotes. Dev. Dyn. 2017, 246, 116–134. [Google Scholar] [CrossRef]
- Vitulo, N.; Dalla Valle, L.; Skobo, T.; Valle, G.; Alibardi, L. Down-regulation of lizard immuno-genes in the regenerating tail and myo-genes in the scarring limb suggests that tail regeneration occurs in an immuno-privileged organ. Protoplasma 2017, 254, 2127–2141. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Original and regenerating lizard tail cartilage contain putative resident stem/progenitor cells. Micron 2015, 78, 10–18. [Google Scholar] [CrossRef]
- Alibardi, L. Immunolocalization indicates that both original and regenerated lizard tail tissues contain populations of long retaining cells, putative stem/progenitor cells. Micr. Res. Tech. 2015, 78, 1032–1045. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Temporal distribution of 5BrdU-labeled cells suggests that most injured tissues contribute proliferating cells for the regeneration of the tail and limb in lizard. Acta Zool. 2019, 100, 303–319. [Google Scholar] [CrossRef]
- Nourhidayat, L.; Benes, V.; Blom, S.; Gomes, I.; Firdausi, N.; de Bakker, M.A.G.; Spaink, H.P.; Richardson, M.K. Tokay gecko tail regeneration involves temporally collinear expression of HOXC genes and early expression of satellite cell markers. BMC Biol. 2025, 23, 6. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Ultrastructural analysis of early regenerating tail suggests that a process of dedifferentiation is involved in the formation of the regenerative blastema. J. Morphol. 2018, 279, 1171–1184. [Google Scholar] [CrossRef]
- Simon, A.; Tanaka, E.M. Limb regeneration. WIREs Dev. Biol. 2013, 2, 291–300. [Google Scholar] [CrossRef]
- Daponte, V.; Tylzanowski, P.; Forlino, A. Appendage regeneration in vertebrates: What makes this possible? Cells 2021, 10, 242. [Google Scholar] [CrossRef]
- Grigoryan, E. Study on natural longlife juvenility and tissue regeneration in caudate amphibians and potential application of resulting data in biomedicine. J. Dev. Biol. 2021, 9, 2. [Google Scholar] [CrossRef]
- Sugiura, T.; Wang, H.; Barsacchi, R.; Simon, A.; Tanaka, E.M. MARCKS-Like proteins is an initiator in axolotl appendage regeneration. Nature 2016, 531, 237–240. [Google Scholar] [CrossRef]
- Alibardi, L. Immunolocalization of 5BrdU long retaining labeled cells and macrophage infiltration in the scarring limb of lizard after limb amputation. Tiss. Cell 2016, 48, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Ouimet, C.C.; Wang, J.K.T.; Walass, S.I.; Albert, K.A.; Greengard, P. Localization of the MARCKS (87 kDa) protein, a major specific substrate for protein kinase C, in rat brain. J. Neurosci. 1990, 10, 1683–1698. [Google Scholar] [CrossRef] [PubMed]
- El Amri, M.; Fitzgerald, U.; Schlosser, G. MARCKS and MARCKS-like proteins in development and regeneration. J. Biomed. Sci. 2018, 25, 43. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, W.; Selmi, C.; Ridgway, W.M.; Leung, P.S.C.; Zhang, F.; Gershwin, M.E. The myristoylated alanine-rich C-kinase substrates (MARCKs): A membrane-anchored mediator of the cell function. Autoimmun. Rev. 2021, 20, 102942. [Google Scholar] [CrossRef]
- McNamara, R.K.; Jiang, Y.; Streit, W.J.; Lenox, R.H. Facial motor neuron regeneration induces a unique spatial and temporal pattern of myristoylated alanine-rich C kinase substrate expression. Neuroscience 2000, 97, 581–589. [Google Scholar] [CrossRef]
- Weimer Weimer, J.M.; Yokota, Y.; Stanco, A.; Stumpo, D.J.; Blackshear, P.J.; Anton, E.S. MARCKS modulates radial progenitor placement, proliferation and organization in the developing cerebral cortex. Development 2009, 136, 2965–2975. [Google Scholar] [CrossRef]
- El Amri, M.; Pandit, A.; Schlosser, G. Marcks and Marcks like-1 proteins promote spinal cord development and regeneration in Xenopus. eLife 2024, 13, e98277. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.F.; Costa, C.M.; Lorena, J.; Moreira, R.N.; Frota-Lima, G.N.; Furtado, C.; Robinson, M.; Amemiya, C.T.; Darnet, S.; Schneider, I. Tetrapod limb and sarcopterygian fin regeneration share a core genetic program. Nat. Commun. 2016, 7, 13364. [Google Scholar] [CrossRef]
- Kim, S.S.; Kim, J.H.; Kim, H.S.; Park, D.E.; Chung, C.H. Involvement of the O-type protein kinase C in translocation of myristoylated alanine-rich C kinase substrate (MARCKS) during myogenesis of chick embryonic myoblasts. Bioch. J. 2000, 347, 139–146. [Google Scholar] [CrossRef]
- Kim, S.S.; Kim, J.H.; Lee, S.H.; Chung, S.S.; Bang, O.S.; Park, D.; Chung, C.H. Involvement of protein phosphatase-1-mediated MARCKS translocation in myogenic differentiation of embryonic muscle cells. J. Cell Sci. 2002, 115, 2465–2473. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, S.; Song, H.; Zhang, X.; Wang, W.; Du, W.; Song, T.; Liang, H.L.; Chen, X.; Wang, Y. Macrophage migration inhibitory factor derived from spinal cord is involved in activation of macrophages following gecko tail amputation. FASEB J. 2019, 33, 14798–14810. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Song, H.; Wang, Y. Self-control of inflammation during tail regeneration of lizards. J. Dev. Biol. 2021, 9, 48. [Google Scholar] [CrossRef]
- Park, J.; Fang, S.; Crews, A.L.; Lin, K.W.; Adler, K.B. MARCKS regulation of mucin secretion by airway epithelium in Vitro. Am. J. Respir. Cell Mol. Biol. 2008, 39, 68–76. [Google Scholar] [CrossRef]
- Rombouts, K.; Mello, T.; Liotta, F.; Galli, A.; Caligiuri, A.; Annunziato, F.; Pinzani, M. MARCKS actin binding capacity mediates actin filament assembly during mitosis in human hepatic stellate cells. Amer. J. Physiol. 2012, 303, C357–C367. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, T.D.; Shah, N.; Jackson, W.P.; Feeney, E.V.; Scanlon, S.; Gish, R.; Khodadadi, R.; Hyde, S.O.; Hicks, P.H.; Anderson, J.C.; et al. The effector domain of MARCKS is a nuclear localization signal that regulates cellular PIP2 levels and nuclear PIP2 localization. PLoS ONE 2015, 10, e0140870. [Google Scholar] [CrossRef] [PubMed]
- McLean, C.E.; Vickaryous, M.K. A novel amniote model of epimorphic regeneration: The leopard gecko, Eublepharis macularius. BMC Dev. Biol. 2011, 11, 50–74. [Google Scholar] [CrossRef]
- Simpson, S.B.; Duffy, M.T. The lizard spinal cord: A model system for the study of spinal cord injury and repair. Progr. Brain Res. 1994, 103, 229–241. [Google Scholar]
- Tokuyama, M.A.; Xu, C.; Fisher, R.E.; Wilson-Rawls, J.; Kusumi, K.; Newbern, J.M. Developmental and adult-specific processes contribute to de novo neuromuscular regeneration in the lizard tail. Dev. Biol. 2018, 43, 287–296. [Google Scholar] [CrossRef]
- Alibardi, L.; Boschetti, F. Immunolabeling for RhoV and Actin in early regenerating tail of the lizard Podarcis muralis suggests involvement in epithelial and mesenchymal cell motility. Acta Zool. 2021, 102, 51–62. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alibardi, L. Activation of Marck-like Genes and Proteins During Initial Phases of Regeneration in the Amputated Tail and Limb of the Lizard Podarcis muralis. J. Dev. Biol. 2025, 13, 12. https://doi.org/10.3390/jdb13020012
Alibardi L. Activation of Marck-like Genes and Proteins During Initial Phases of Regeneration in the Amputated Tail and Limb of the Lizard Podarcis muralis. Journal of Developmental Biology. 2025; 13(2):12. https://doi.org/10.3390/jdb13020012
Chicago/Turabian StyleAlibardi, Lorenzo. 2025. "Activation of Marck-like Genes and Proteins During Initial Phases of Regeneration in the Amputated Tail and Limb of the Lizard Podarcis muralis" Journal of Developmental Biology 13, no. 2: 12. https://doi.org/10.3390/jdb13020012
APA StyleAlibardi, L. (2025). Activation of Marck-like Genes and Proteins During Initial Phases of Regeneration in the Amputated Tail and Limb of the Lizard Podarcis muralis. Journal of Developmental Biology, 13(2), 12. https://doi.org/10.3390/jdb13020012