

Principles of Zebrafish Nephron Segment Development



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Essential Functions of the Kidney
1.2. Vertebrate Kidney Forms Are Comprised of Nephrons
2. Using the Zebrafish to Study Nephron Development
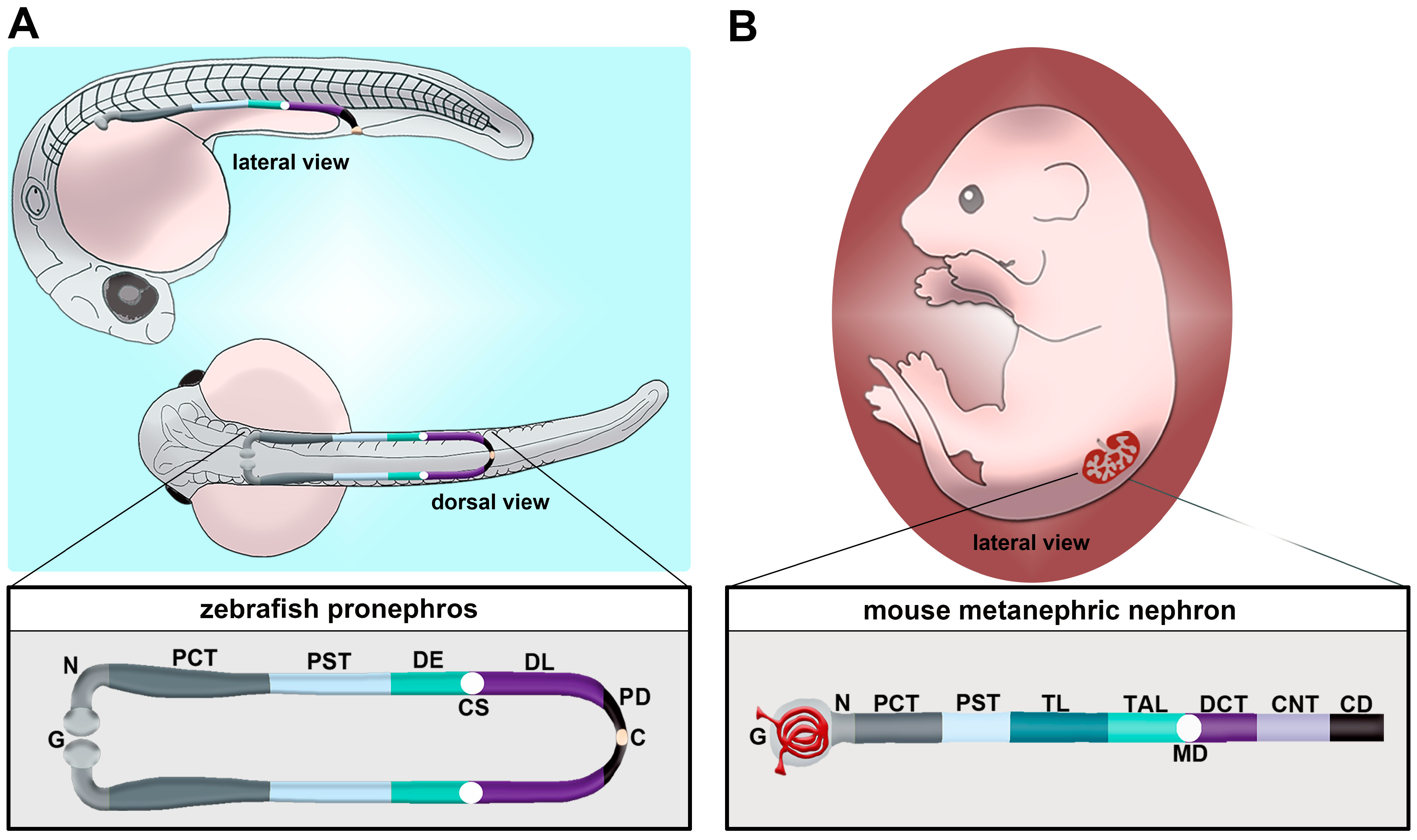
2.1. Zebrafish Pronephros Composition and Function
2.2. Zebrafish Mesonephros Composition and Function: Spotlight on Renal Regeneration Studies
3. Molecular Genetic Toolkit in the Zebrafish Animal Model
3.1. Zebrafish as a Model for Developmental Biology and Biomedical Research
3.2. Forward Genetics: From Random Mutagenesis Screens to Chemical Screens
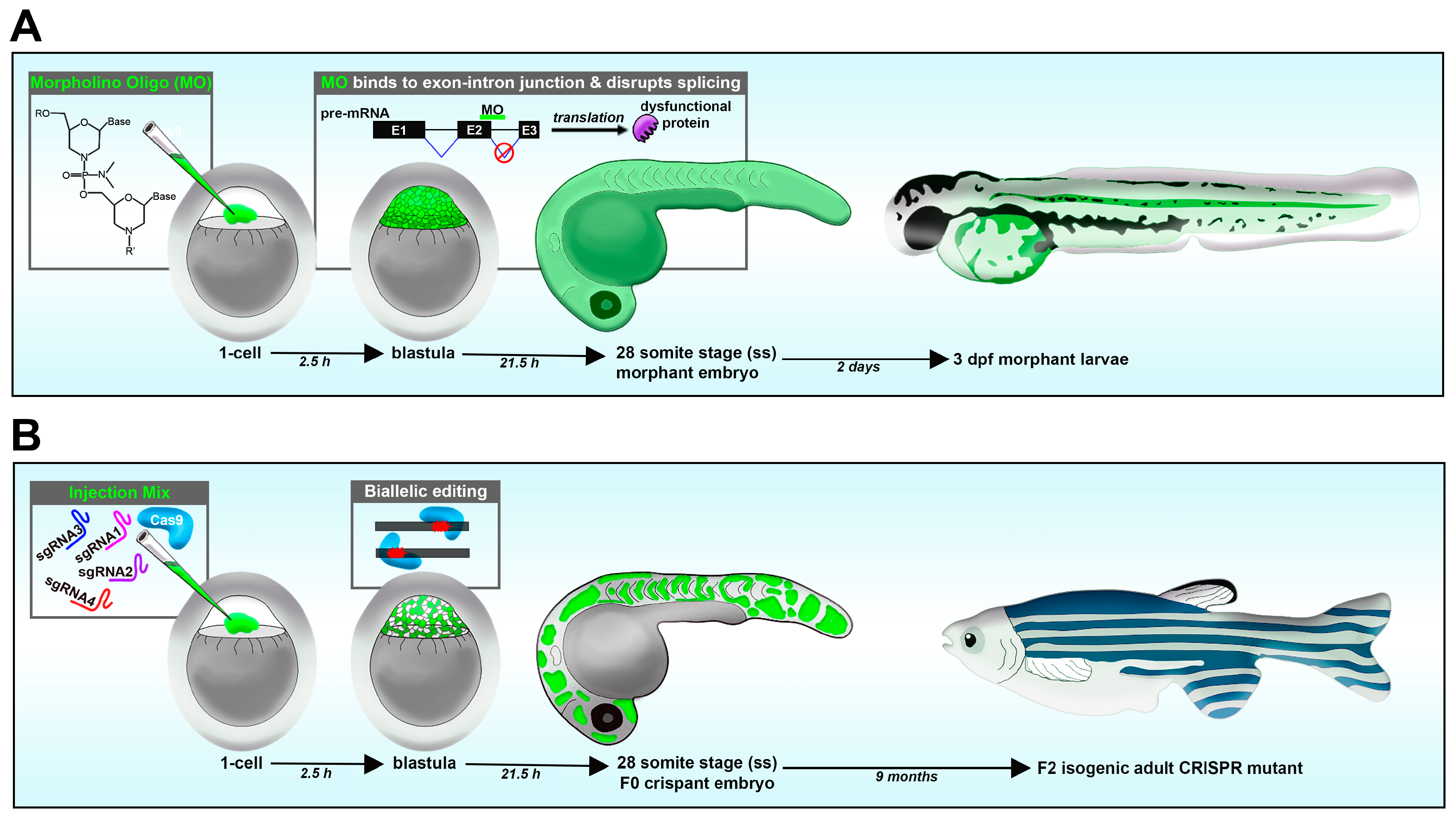
3.3. Reverse Genetics: Loss-of-Function Methods Using Morpholinos and Genome Editing
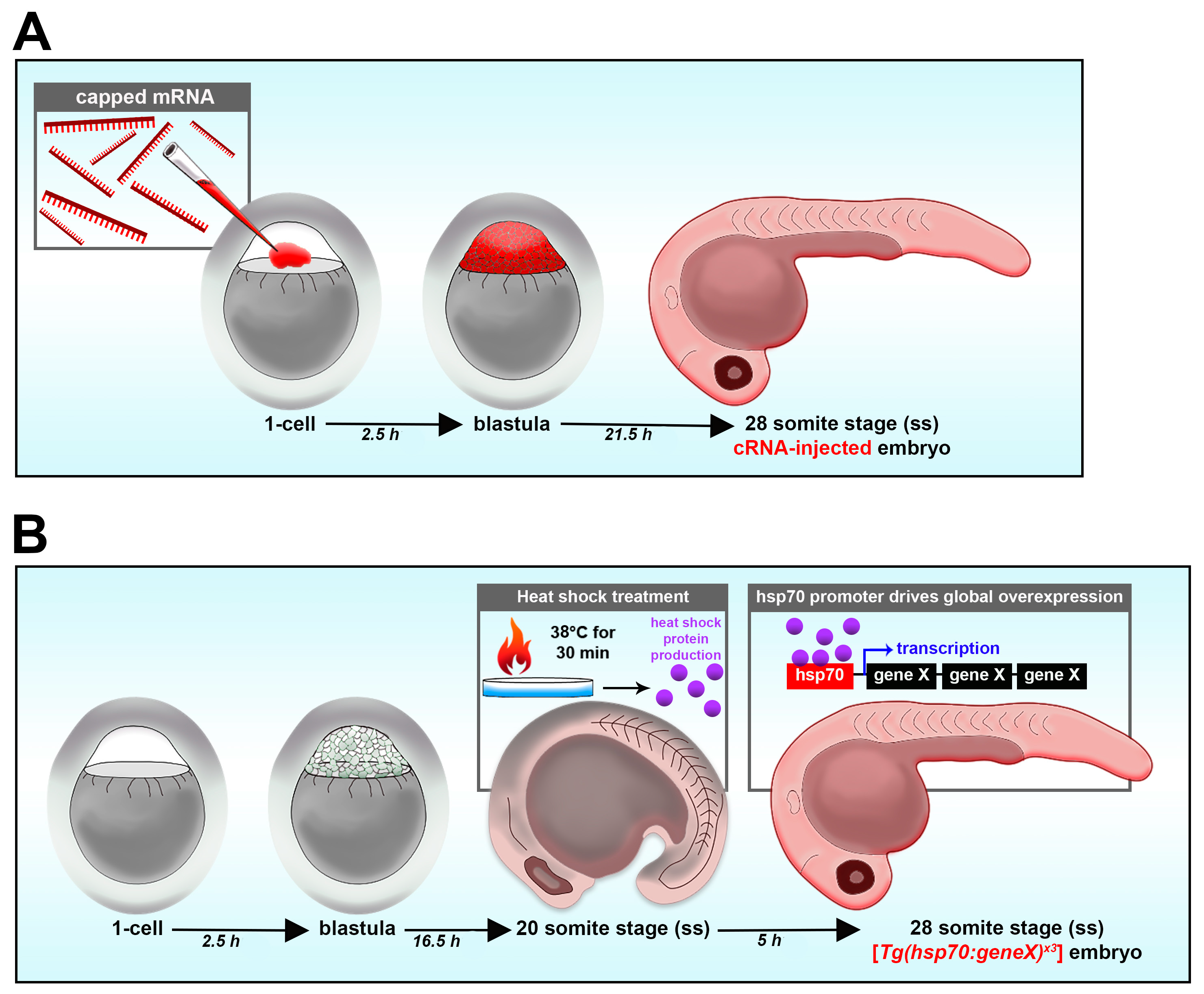
3.4. Reverse Genetics: Gain-of-Function Approaches with mRNA and Transgenic Models
4. Advances in Understanding Distal Segment Development
4.1. The Mecom/Tbx2a/2b/Emx1 Network in DL Pronephros Development
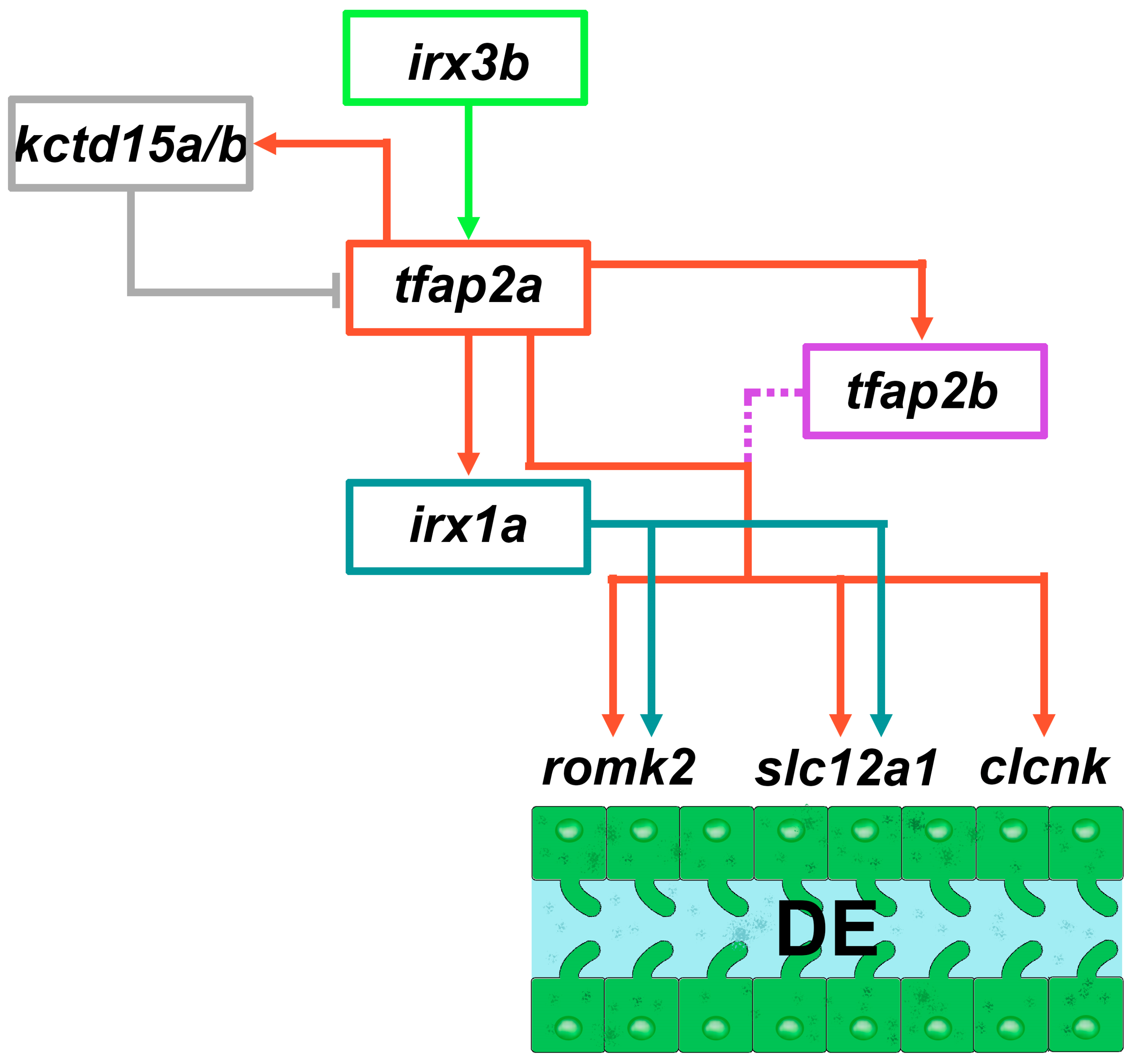
4.2. Role of Irx3b and Irx1a in DE Pronephros Development
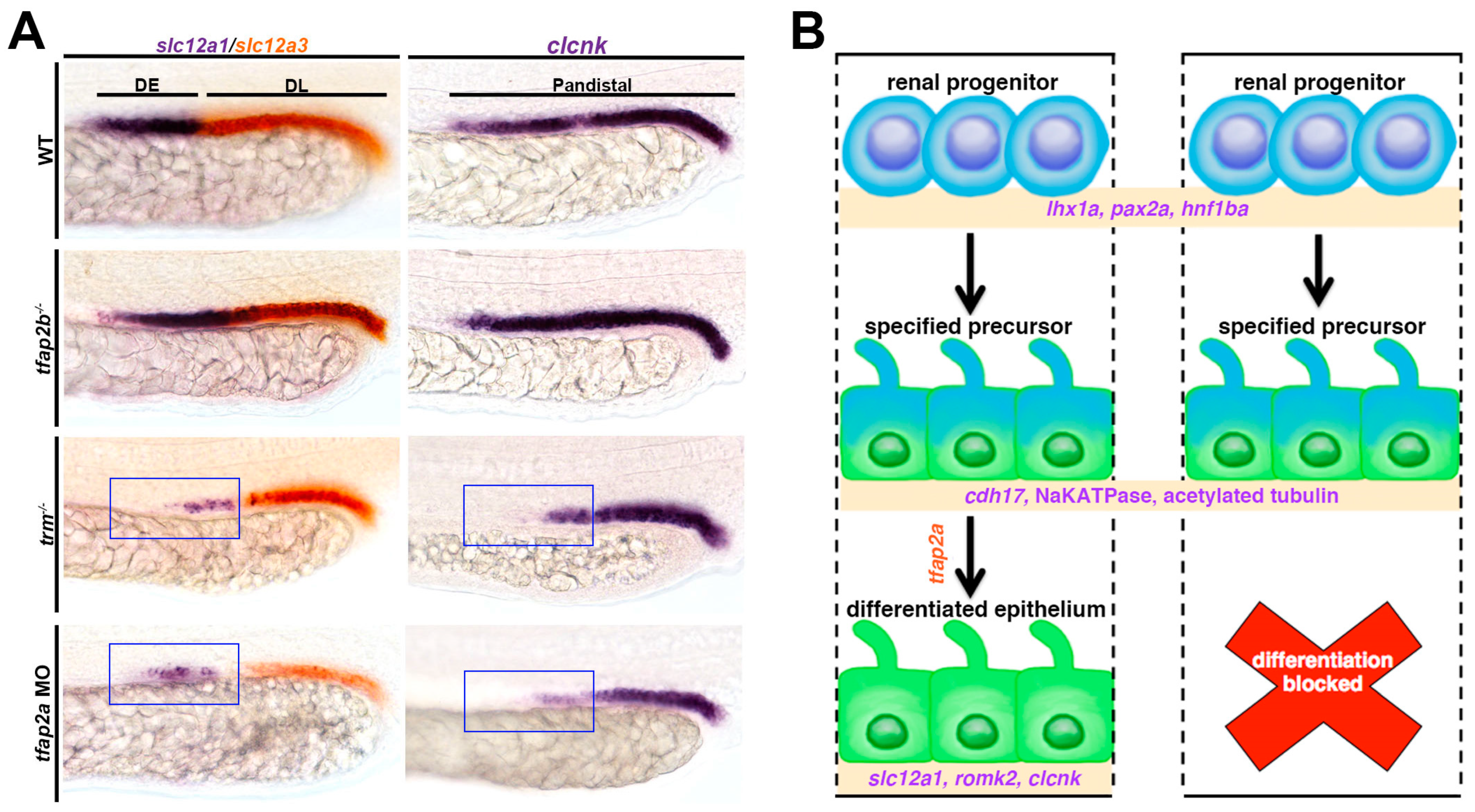
4.3. Tfap2a/b Control DE Differentiation/Maturation
4.4. Tfap2a Autoregulation through the Kctd15a/b Repressors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Preuss, H.G. Basics of renal anatomy and physiology. Clin. Lab. Med. 1993, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hoenig, M.P.; Zeidel, M.L. Homeostasis, the milieu intérieur, and the wisdom of the nephron. Clin. J. Am. Soc. Nephrol. 2014, 9, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Balzer, M.S.; Rohacs, T.; Susztak, K. How many cell types are in the kidney and what do they do? Annu. Rev. Physiol. 2022, 84, 507–531. [Google Scholar] [CrossRef] [PubMed]
- Pollak, M.R.; Quaggin, S.E.; Hoenig, M.P.; Dworkin, L.D. The glomerulus: The sphere of influence. Clin. J. Am. Soc. Nephrol. 2014, 9, 1461–1469. [Google Scholar] [CrossRef]
- Garg, P. A review of podocyte biology. Am. J. Nephrol. 2018, 47, 3–13. [Google Scholar] [CrossRef]
- Zhuo, J.L.; Li, X.C. Proximal nephron. Compr. Physiol. 2013, 3, 1079–1123. [Google Scholar] [PubMed]
- Curthoys, N.P.; Moe, O.W. Proximal tubule function and response to acidosis. Clin. J. Am. Soc. Nephrol. 2014, 9, 1627–1638. [Google Scholar] [CrossRef]
- Dantzler, W.H.; Layton, A.T.; Layton, H.E.; Pannabecker, T.L. Urine-concentrating mechanism in the inner medulla: Function of the thin limbs of the loops of Henle. Clin. J. Am. Soc. Nephrol. 2014, 9, 1781–1789. [Google Scholar] [CrossRef]
- Mount, D.B. Thick ascending limb of the loop of Henle. Clin. J. Am. Soc. Nephrol. 2014, 9, 1974–1986. [Google Scholar] [CrossRef]
- Subramanya, A.R.; Ellison, D.H. Distal convoluted tubule. Clin. J. Am. Soc. Nephrol. 2014, 9, 2147–2163. [Google Scholar] [CrossRef]
- Pearce, D.; Soundararajan, R.; Trimpert, C.; Kashlan, O.B.; Deen, P.M.T.; Kohan, D.E. Collecting duct principal cell transport processes and their regulation. Clin. J. Am. Soc. Nephrol. 2015, 10, 135–146. [Google Scholar] [CrossRef]
- Roy, A.; Al-bataineh, M.M.; Pastor-Soler, N.M. Collecting duct intercalated cell function and regulation. Clin. J. Am. Soc. Nephrol. 2015, 10, 305–324. [Google Scholar] [CrossRef] [PubMed]
- Bertram, J.F.; Douglas-Denton, R.N.; Diouf, B.; Hughson, M.D.; Hoy, W.E. Human nephron number: Implications for health and disease. Pediatr. Nephrol. 2011, 26, 1529–1533. [Google Scholar] [CrossRef]
- Luyckx, V.A.; Shukha, K.; Brenner, B.M. Low nephron number and its clinical consequences. Rambam Maimonides Med. J. 2011, 2, e0061. [Google Scholar] [CrossRef] [PubMed]
- Black, M.J.; Sutherland, M.R.; Gubhaju, L.; Kent, A.L.; Dahlstrom, J.E.; Moore, L. When birth comes early: Effects on nephrogenesis. Nephrology (Carlton) 2013, 18, 180–182. [Google Scholar] [CrossRef]
- McCampbell, K.K.; Wingert, R.A. Renal stem cells: Fact or science fiction? Biochem. J. 2012, 444, 153–168. [Google Scholar] [CrossRef]
- Schedl, A. Renal abnormalities and their developmental origin. Nat. Rev. Genet. 2007, 8, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Dursun, H.; Bayazit, A.K.; Büyükçelik, M.; Soran, M.; Noyan, A.; Anarat, A. Associated anomalies in children with congenital solitary functioning kidney. Pediatr. Surg. Int. 2005, 21, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Westland, R.; Schreuder, M.F.; Ket, J.C.F.; van Wijk, J.A.E. Unilateral renal agenesis: A systematic review on associated anomalies and renal injury. Nephrol. Dial. Transplant. 2013, 28, 1844–1855. [Google Scholar] [CrossRef]
- Vivante, A.; Kohl, S.; Hwang, D.; Dworschak, G.C.; Hildebrandt, F. Single-gene causes of congenital anomalies of the kidney and urinary tract (CAKUT) in humans. Pediatr. Nephrol. 2014, 29, 695–704. [Google Scholar] [CrossRef]
- Rodriguez, M.M. Congenital anomalies of the kidney and the urinary tract (CAKUT). Fetal Pediatr. Pathol. 2014, 33, 293–320. [Google Scholar] [CrossRef]
- Nicolaou, N.; Renkema, K.Y.; Bongers, E.M.; Giles, R.H.; Knoers, N.V. Genetic, environmental, and epigenetic factors involved in CAKUT. Nat. Rev. Nephrol. 2015, 11, 720–731. [Google Scholar] [CrossRef]
- Romagnani, P.; Lasagni, L.; Remuzzi, G. Renal progenitors: An evolutionary conserved strategy for kidney regeneration. Nat. Rev. Nephrol. 2013, 9, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Peired, A.J.; Melica, M.E.; Molli, A.; Nardi, C.; Romagnani, P.; Lasagni, L. Molecular mechanisms of renal progenitor regulation: How many pieces in the puzzle? Cells 2021, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Naved, B.A.; Bonventre, J.V.; Hubbell, J.A.; Hukriede, N.A.; Humphreys, B.D.; Kesselman, C.; Valerius, M.T.; McMahon, A.P.; Shankland, S.J.; Wertheim, J.A.; et al. Kidney repair and regeneration: Perspectives of the NIDDK (Re)Building a Kidney consortium. Kidney Int. 2022, 101, 845–853. [Google Scholar] [CrossRef]
- Chambers, J.M.; Wingert, R.A. Advances in understanding vertebrate nephrogenesis. Tissue Barriers 2020, 8, e1832844. [Google Scholar] [CrossRef] [PubMed]
- Little, M.H. Returning to kidney development to deliver synthetic kidneys. Dev. Biol. 2021, 474, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Beyenbach, K.W. Kidneys sans glomeruli. Am. J. Physiol. Renal Physiol. 2004, 286, F811–F827. [Google Scholar] [CrossRef]
- Holz, P.H. Anatomy and physiology of the reptile renal system. Vet. Clin. N. Am. Exot. Anim. Pract. 2020, 23, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Orosz, S.E.; Echols, M.S. The urinary and osmoregulatory systems of birds. Vet. Clin. N. Am. Exot. Anim. Pract. 2020, 23, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Keogh, L.; Kilroy, D.; Bhattacharjee, S. The struggle to equilibrate outer and inner milieus: Renal evolution revisited. Ann. Anat. 2021, 233, 151610. [Google Scholar] [CrossRef]
- Schnell, J.; Achieng, M.; Lindström, N.O. Principles of human and mouse nephron development. Nat. Rev. Nephrol. 2022, 18, 628–642. [Google Scholar] [CrossRef]
- Senarat, S.; Kettratad, J.; Pairohakul, S.; Ampawong, S.; Huggins, B.P.; Coleman, M.M.; Kaneko, G. An update on the evolutionary origin of aglomerular kidney with structural and ultrastructural descriptions of the kidney in three fish species. J. Fish Biol. 2022, 100, 1283–1298. [Google Scholar] [CrossRef]
- Evans, R.G. Evolution of the glomerulus in a marine environment and its implications for renal function in terrestrial vertebrates. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2023, 324, R143–R151. [Google Scholar] [CrossRef]
- Wingert, R.A.; Davidson, A.J. The zebrafish pronephros: A model to study nephron segmentation. Kidney Int. 2008, 73, 1120–1127. [Google Scholar] [CrossRef]
- Ebarasi, L.; Oddsson, A.; Hultenby, K.; Betsholtz, C.; Tryggvason, K. Zebrafish: A model system for the study of vertebrate renal development, function, and pathophysiology. Curr. Opin. Nephrol. Hypertens. 2011, 20, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, G.F.; Wingert, R.A. Kidney organogenesis in the zebrafish: Insights into vertebrate nephrogenesis and regeneration. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 559–585. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, P.T.; Wingert, R.A. Using zebrafish to study podocyte genesis during kidney development and regeneration. Genesis 2014, 52, 771–792. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.J. Kidney regeneration in fish. Nephron Exp. Nephrol. 2014, 126, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Desgrange, A.; Cereghini, S. Nephron patterning: Lessons from Xenopus, zebrafish and mouse studies. Cells 2015, 4, 483–499. [Google Scholar] [CrossRef]
- McKee, R.A.; Wingert, R.A. Zebrafish renal pathology: Emerging models of acute kidney injury. Curr. Pathobiol. Rep. 2015, 3, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Drummond, B.E.; Wingert, R.A. Insights into kidney stem cell development and regeneration using zebrafish. World J. Stem Cells 2016, 8, 22–31. [Google Scholar] [CrossRef]
- Marra, A.N.; Li, Y.; Wingert, R.A. Antennas of organ morphogenesis: The roles of cilia in vertebrate kidney development. Genesis 2016, 54, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Drummond, I.A.; Davidson, A.J. Zebrafish kidney development. Methods Cell Biol. 2016, 134, 391–429. [Google Scholar] [PubMed]
- Poureetezadi, S.J.; Wingert, R.A. Little fish, big catch: Zebrafish as a model for kidney disease. Kidney Int. 2016, 89, 1204–1210. [Google Scholar] [CrossRef]
- Morales, E.E.; Wingert, R.A. Zebrafish as a model of kidney disease. Results Probl. Cell Differ. 2017, 60, 55–75. [Google Scholar]
- Elmonem, M.A.; Berlingerio, S.P.; van den Heuvel, L.P.; de Witte, P.A.; Lowe, M.; Levtchenko, E.N. Genetic renal diseases: The emerging role of zebrafish models. Cells 2018, 7, 130. [Google Scholar] [CrossRef]
- Chambers, B.E.; Wingert, R.A. Renal progenitors: Roles in kidney disease and regeneration. World J. Stem Cells 2016, 8, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Adhish, M.; Manjubala, I. Effectiveness of zebrafish models in understanding human diseases—A review of models. Heliyon 2023, 9, e14557. [Google Scholar] [CrossRef]
- Fatma, S.; Nayak, U.; Swain, R.K. Methods to generate and evaluate zebrafish models of human kidney diseases. Int. J. Dev. Biol. 2021, 65, 475–485. [Google Scholar] [CrossRef]
- Wesselman, H.M.; Nguyen, T.K.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Advances in understanding the genetic mechanisms of zebrafish renal multiciliated cell development. J. Dev. Biol. 2022, 11, 1. [Google Scholar] [CrossRef]
- Bolten, J.S.; Pratsinis, A.; Alter, C.L.; Fricker, G.; Huwyler, J. Zebrafish (Danio rerio) larva as an in vivo vertebrate model to study renal function. Am. J. Physiol. Renal Physiol. 2022, 322, F280–F294. [Google Scholar] [CrossRef] [PubMed]
- Drummond, I.A.; Majumdar, A.; Hentschel, H.; Elger, M.; Solnica-Krezel, L.; Schier, A.F.; Neuhauss, S.C.; Stemple, D.L.; Zwartkruis, F.; Rangini, Z.; et al. Early development of the zebrafish pronephros and analysis of mutations affecting pronephric function. Development 1998, 125, 4655–4667. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, A.; Drummond, I.A. Podocyte differentiation in the absence of endothelial cells as revealed in the zebrafish avascular mutant, cloche. Dev. Genet. 1999, 24, 220–229. [Google Scholar] [CrossRef]
- Majumdar, A.; Drummond, I.A. The zebrafish floating head mutant demonstrates podocytes play an important role in directing glomerular differentiation. Dev. Biol. 2000, 222, 147–157. [Google Scholar] [CrossRef]
- Serluca, F.C.; Fishman, M.C. Pre-pattern in the pronephric kidney field of zebrafish. Development 2001, 128, 2233–2241. [Google Scholar] [CrossRef]
- Hsu, H.; Lin, G.; Chung, B. Parallel early development of zebrafish interrenal glands and pronephros: Differential control by Wt1 and Ff1b. Development 2003, 130, 2107–2116. [Google Scholar] [CrossRef] [PubMed]
- Drummond, B.E.; Ercanbrack, W.S.; Wingert, R.A. Modeling podocyte ontogeny and podocytopathies with the zebrafish. J. Dev. Biol. 2023, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Zucker, A.G.; Wiessner, S.; Jensen, A.M.; Drummond, I.A. Organization of the pronephric filtration apparatus in zebrafish requires Nephrin, Podocin and the FERM domain protein Mosaic eyes. Dev. Biol. 2005, 285, 316–329. [Google Scholar] [CrossRef]
- Kramer-Zucker, A.G.; Olale, F.; Haycraft, C.J.; Yoder, B.K.; Schier, A.F.; Drummond, I.A. Cilia-driven fluid flow in the zebrafish pronephros, brain and Kupffer’s vesicle is required for normal organogenesis. Development 2005, 132, 1907–1921. [Google Scholar] [CrossRef]
- Bollig, F.; Mehringer, R.; Perner, B.; Hartung, C.; Schäfer, M.; Schartl, M.; Volff, J.; Winkler, C.; Englert, C. Identification and comparative expression analysis of a second Wt1 gene in zebrafish. Dev. Dyn. 2006, 235, 554–561. [Google Scholar] [CrossRef]
- Perner, B.; Englert, C.; Bollig, F. The Wilms tumor genes Wt1a and Wt1b control different steps during formation of the zebrafish pronephros. Dev. Biol. 2007, 309, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Jiang, Y.J. Jagged2a-notch signaling mediates cell fate choice in the zebrafish pronephric duct. PLoS Genet. 2007, 3, e18. [Google Scholar] [CrossRef]
- Liu, Y.; Pathak, N.; Kramer-Zucker, A.; Drummond, I.A. Notch signaling controls the differentiation of transporting epithelia and multiciliated cells in the zebrafish pronephros. Development 2007, 134, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Ikenaga, T.; Urban, J.M.; Gebhart, N.; Hatta, K.; Kawakami, K.; Ono, F. Formation of the spinal network in zebrafish determined by domain-specific pax genes. J. Comp. Neurol. 2011, 519, 1562–1579. [Google Scholar] [CrossRef]
- Tena, J.J.; Neto, A.; de la Calle-Mustienes, E.; Bras-Pereira, C.; Casares, F.; Gómez-Skarmeta, J.L. Odd-Skipped genes encode repressors that control kidney development. Dev. Biol. 2007, 301, 518–531. [Google Scholar] [CrossRef] [PubMed]
- Mudumana, S.P.; Hentschel, D.; Liu, Y.; Vasilyev, A.; Drummond, I.A. odd skipped related1 reveals a novel role for endoderm in regulating kidney versus vascular cell fate. Development 2008, 135, 3355–3367. [Google Scholar] [CrossRef]
- Neto, A.; Mercader, N.; Gómez-Skarmeta, J.L. The Osr1 and Osr2 genes act in the pronephric anlage downstream of retinoic acid signaling and upstream of Wnt2b to maintain pectoral fin development. Development 2012, 139, 301–311. [Google Scholar] [CrossRef]
- Tomar, R.; Mudumana, S.P.; Pathak, N.; Hukriede, N.A.; Drummond, I.A. Osr1 is required for podocyte development downstream of Wt1a. J. Am. Soc. Nephrol. 2014, 25, 2539–2545. [Google Scholar] [CrossRef]
- Perens, E.A.; Garavito-Aguilar, Z.V.; Guio-Vega, G.P.; Peña, K.T.; Schindler, Y.L.; Yelon, D. Hand2 inhibits kidney specification while promoting vein formation within the posterior mesoderm. eLife 2016, 5, e19941. [Google Scholar] [CrossRef]
- Perens, E.A.; Diaz, J.T.; Quesnel, A.; Crump, J.G.; Yelon, D. osr1 couples intermediate mesoderm cell fate with temporal dynamics of vessel progenitor cell differentiation. Development 2021, 148, dev198408. [Google Scholar] [CrossRef] [PubMed]
- Drummond, B.E.; Chambers, B.E.; Wesselman, H.M.; Gibson, S.; Arceri, L.; Ulrich, M.N.; Gerlach, G.F.; Kroeger, P.T.; Leshchiner, I.; Goessling, W.; et al. osr1 maintains renal progenitors and regulates podocyte development by promoting wnt2ba via the antagonism of hand2. Biomedicines 2022, 10, 2868. [Google Scholar] [CrossRef] [PubMed]
- Wingert, R.A.; Selleck, R.; Yu, J.; Song, H.; Chen, Z.; Song, A.; Zhou, Y.; Thisse, B.; Thisse, C.; McMahon, A.P.; et al. The cdx genes and retinoic acid control the positioning and segmentation of the zebrafish pronephros. PLoS Genet. 2007, 3, e189. [Google Scholar] [CrossRef]
- Ebarasi, L.; He, L.; Hultenby, K.; Takemoto, M.; Betsholtz, C.; Tryggvason, K.; Majumdar, A. A reverse genetic screen in the zebrafish identifies Crb2b as a regulator of the glomerular filtration barrier. Dev. Biol. 2009, 334, 1–9. [Google Scholar] [CrossRef]
- Lyons, J.P.; Miller, R.K.; Zhou, X.; Weidinger, G.; Deroo, T.; Denayer, T.; Park, J.; Ji, H.; Hong, J.Y.; Li, A.; et al. Requirement of Wnt/Β-Catenin signaling in pronephric kidney development. Mech. Dev. 2009, 126, 142–159. [Google Scholar] [CrossRef] [PubMed]
- de Groh, E.D.; Swanhart, L.M.; Cosentino, C.C.; Jackson, R.L.; Dai, W.; Kitchens, C.A.; Day, B.W.; Smithgall, T.E.; Hukriede, N.A. Inhibition of histone deacetylase expands the renal progenitor cell population. J. Am. Soc. Nephrol. 2010, 21, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Wingert, R.A.; Davidson, A.J. Zebrafish nephrogenesis involves dynamic spatiotemporal expression changes in renal progenitors and essential signals from retinoic acid and irx3b. Dev. Dyn. 2011, 240, 2011–2027. [Google Scholar] [CrossRef]
- O’Brien, L.L.; Grimaldi, M.; Kostun, Z.; Wingert, R.A.; Selleck, R.; Davidson, A.J. Wt1a, Foxc1a, and the Notch Mediator Rbpj physically interact and regulate the formation of podocytes in zebrafish. Dev. Biol. 2011, 358, 318–330. [Google Scholar] [CrossRef]
- Wang, H.; Lehtonen, S.; Chen, Y.C.; Heikkila, E.; Panula, P.; Holthofer, H. Neph3 associates with regulation of glomerular and neural development in zebrafish. Differentiation 2012, 83, 38–46. [Google Scholar] [CrossRef]
- Ichimura, K.; Powell, R.; Nakamura, T.; Kurihara, H.; Sakai, T.; Obara, T. Podocalyxin regulates pronephric glomerular development in zebrafish. Physiol. Rep. 2013, 1, e00074. [Google Scholar] [CrossRef]
- Naylor, R.W.; Przepiorski, A.; Ren, Q. HNF1B is essential for nephron segmentation during nephrogenesis. J. Am. Soc. Nephrol. 2013, 24, 77–87. [Google Scholar] [CrossRef]
- Gerlach, G.F.; Wingert, R.A. Zebrafish pronephros tubulogenesis and epithelial identity maintenance are reliant on the polarity proteins Prkc iota and zeta. Dev. Biol. 2014, 396, 183–200. [Google Scholar] [CrossRef] [PubMed]
- McKee, R.; Gerlach, G.F.; Jou, J.; Cheng, C.N.; Wingert, R.A. Temporal and spatial expression of tight junction genes during zebrafish pronephros development. Gene Expr. Patterns 2014, 16, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Fukuyo, Y.; Nakamura, T.; Bubenshchikova, E.; Powell, R.; Tsuji, T.; Janknecht, R.; Obara, T. Nephrin and Podocin functions are highly conserved between the zebrafish pronephros and mammalian metanephros. Mol. Med. Rep. 2014, 9, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.N.; Li, Y.; Marra, A.N.; Verdun, V.; Wingert, R.A. Flat mount preparation for observation and analysis of zebrafish embryo specimens stained by whole mount in situ hybridization. J. Vis. Exp. 2014, 89, 51604. [Google Scholar]
- Cheng, C.N.; Wingert, R.A. Nephron proximal tubule patterning and corpuscles of Stannius formation are regulated by the sim1a transcription factor and retinoic acid in zebrafish. Dev. Biol. 2015, 399, 100–116. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Pietsch, S.; Tan, Z.; Perner, B.; Sierig, R.; Kruspe, D.; Groth, M.; Witzgall, R.; Gröne, H.; Platzer, M.; et al. Integration of cistromic and transcriptomic analyses identifies Nphs2, Mafb, and Magi2 as Wilms’ Tumor 1 target genes in podocyte differentiation and maintenance. J. Am. Soc. Nephrol. 2015, 26, 2118–2128. [Google Scholar] [CrossRef]
- Marra, A.N.; Wingert, R.A. Epithelial cell fate in the nephron tubule is mediated by the ETS transcription factors etv5a and etv4 during zebrafish kidney development. Dev. Biol. 2016, 411, 231–245. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, Z.; Zeng, C.; Wang, L.; Xu, F.; Hou, Q.; Liu, Z. Ultrastructural characterization of the pronephric glomerulus development in zebrafish. J. Morphol. 2016, 277, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, P.T.; Drummond, B.E.; Miceli, R.; McKernan, M.; Gerlach, G.F.; Marra, A.N.; Fox, A.; McCampbell, K.K.; Leshchiner, I.; Rodriguez-Mari, A.; et al. The zebrafish kidney mutant zeppelin reveals that brca2/fancd1 is essential for pronephros development. Dev. Biol. 2017, 428, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Boucher, R.C.; Bollig, F.; Englert, C.; Hildebrandt, F. Characterization of mesonephric development and regeneration using transgenic zebrafish. Am. J. Physiol. Renal Physiol. 2010, 299, F1040–F1047. [Google Scholar] [CrossRef]
- Diep, C.Q.; Peng, Z.; Ukah, T.K.; Kelly, P.M.; Daigle, R.V.; Davidson, A.J. Development of the zebrafish mesonephros. Genesis 2015, 53, 257–269. [Google Scholar] [CrossRef]
- Reimschuessel, R. A fish model of renal regeneration and development. ILAR J. 2001, 42, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Diep, C.Q.; Ma, D.; Deo, R.C.; Holm, T.M.; Naylor, R.W.; Arora, N.; Wingert, R.A.; Bollig, F.; Djordjevic, G.; Lichman, B.; et al. Identification of adult nephron progenitors capable of kidney regeneration in zebrafish. Nature 2011, 470, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.J. Uncharted waters: Nephrogenesis and renal regeneration in fish and mammals. Pediatr. Nephrol. 2011, 26, 1435–1443. [Google Scholar] [CrossRef]
- McCampbell, K.K.; Wingert, R.A. New tides: Using zebrafish to study renal regeneration. Transl. Res. 2012, 163, 109–122. [Google Scholar] [CrossRef]
- McCampbell, K.K.; Springer, K.N.; Wingert, R.A. Analysis of nephron composition and function in the adult zebrafish kidney. J. Vis. Exp. 2014, 90, e51644. [Google Scholar]
- McCampbell, K.K.; Springer, K.N.; Wingert, R.A. Atlas of cellular dynamics during zebrafish adult kidney regeneration. Stem Cells Int. 2015, 2015, 547636. [Google Scholar] [CrossRef]
- Kamei, C.N.; Gallegos, T.F.; Liu, Y.; Hukriede, N.; Drummond, I.A. Wnt signaling mediates new nephron formation during zebrafish kidney regeneration. Development 2019, 146, dev168294. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yu, T.; He, X.; Fu, Y.; Dai, L.; Wang, B.; Wu, Y.; He, J.; Li, Y.; Zhang, F.; et al. Dual roles of hydrogen peroxide in promoting zebrafish renal repair and regeneration. Biochem. Biophys. Res. Commun. 2019, 516, 680–685. [Google Scholar] [CrossRef]
- Gallegos, T.F.; Kamei, C.N.; Rohly, M.; Drummond, I.A. Fibroblast growth factor signaling mediates progenitor cell aggregation and nephron regeneration in the adult zebrafish kidney. Dev. Biol. 2019, 454, 44–51. [Google Scholar] [CrossRef]
- Liu, X.; Yu, T.; Tan, X.; Jin, D.; Yang, W.; Zhang, J.; Dai, L.; He, Z.; Li, D.; Zhang, Y.; et al. Renal interstitial cells promote nephron regeneration by secreting prostaglandin E2. eLife 2023, 12, e81438. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.B.; Westerfield, M. Zebrafish models in translational research: Tipping the scales toward advancements in human health. Dis. Model Mech. 2014, 7, 739–743. [Google Scholar] [CrossRef]
- Irion, U.; Nüsslein-Volhard, C. Developmental genetics with model organisms. Proc. Natl. Acad. Sci. USA 2022, 119, e2122148119. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.; Torroja, C.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Lawson, N.D.; Wolfe, S.A. Forward and reverse genetic approaches for the analysis of vertebrate development in the zebrafish. Dev. Cell 2011, 21, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Molinari, E.; Sayer, J.A. Disease modeling to understand the pathomechanisms of human genetic kidney disorders. Clin. J. Am. Soc. Nephrol. 2020, 15, 855–872. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Amsterdam, A.; Pazour, G.J.; Cole, D.G.; Miller, M.S.; Hopkins, N. A genetic screen in zebrafish identifies cilia genes as a principal cause of cystic kidney. Development 2004, 131, 4085–4093. [Google Scholar] [CrossRef]
- Kroeger, P.T.; Poureetezadi, S.J.; McKee, R.; Jou, J.; Miceli, R.; Wingert, R.A. Production of haploid zebrafish embryos by in vitro fertilization. J. Vis. Exp. 2014, 89, 51708. [Google Scholar]
- Ebarasi, L.; Ashraf, S.; Bierzynska, A.; Gee, H.Y.; McCarthy, H.J.; Lovric, S.; Sadowski, C.E.; Pabst, W.; Vega-Warner, V.; Fang, H.; et al. Defects of CRB2 cause steroid-resistant nephrotic syndrome. Am. J. Hum. Genet. 2015, 96, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Gehrig, J.; Pandey, G.; Westhoff, J.H. Zebrafish as a model for drug screening in genetic kidney diseases. Front. Pediatr. 2018, 6, 183. [Google Scholar] [CrossRef]
- Skromne, I.; Prince, V.E. Current perspectives in zebrafish reverse genetics: Moving forward. Dev. Dyn. 2008, 237, 861–882. [Google Scholar] [CrossRef] [PubMed]
- Nasevicius, A.; Ekker, S.C. Effective targeted gene ‘knockdown’ in zebrafish. Nat. Genet. 2000, 26, 216–220. [Google Scholar] [CrossRef]
- Draper, B.W.; Morcos, P.A.; Kimmel, C.B. Inhibition of zebrafish fgf8 pre-mRNA splicing with morpholino oligos: A quantifiable method for gene knockdown. Genesis 2001, 30, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Doyon, Y.; McCammon, J.M.; Miller, J.C.; Faraji, F.; Ngo, C.; Katibah, G.E.; Amora, R.; Hocking, T.D.; Zhang, L.; Rebar, E.J.; et al. Heritable targeted gene disruption in zebrafish using designed zinc-finger nucleases. Nat. Biotechnol. 2008, 26, 702–708. [Google Scholar] [CrossRef]
- Meng, X.; Noyes, M.B.; Zhu, L.J.; Lawson, N.D.; Wolfe, S.A. Targeted gene inactivation in zebrafish using engineered zinc-finger nucleases. Nat. Biotechnol. 2008, 26, 695–701. [Google Scholar] [CrossRef]
- Gupta, A.; Meng, X.; Zhu, L.J.; Lawson, N.D.; Wolfe, S.A. Zinc finger protein-dependent and -independent contributions to the in vivo off-target activity of zinc finger nucleases. Nucleic Acids Res. 2011, 39, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Smith, T.; McNulty, J.; Rayla, A.L.; Lakshmanan, A.; Siekmann, A.F.; Buffardi, M.; Meng, X.; Shin, J.; Padmanabhan, A.; et al. Evaluation and application of modularly assembled zinc-finger nucleases in zebrafish. Development 2011, 138, 4555–4564. [Google Scholar] [CrossRef]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Joung, J.K.; Sander, J.D. TALENs: A widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Biol. 2013, 14, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.M.; Musunuru, K. Expanding the genetic editing tool kit: ZFNs, TALENs, and CRISPR-Cas9. J. Clin. Investig. 2014, 124, 4154–4161. [Google Scholar] [CrossRef]
- Barrangou, R.; Marraffini, L.A. CRISPR-Cas systems: Prokaryotes upgrade to adaptive immunity. Mol. Cell 2014, 54, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Jao, L.-E.; Wente, S.R.; Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc. Natl. Acad. Sci. USA 2013, 110, 13904–13909. [Google Scholar] [CrossRef]
- Auer, T.O.; Duroure, K.; De Cian, A.; Concordet, J.-P.; Del Bene, F. Highly efficient CRISPR/Cas9-mediated knock-in in zebrafish by homology-independent DNA repair. Genome Res. 2014, 24, 142–153. [Google Scholar] [CrossRef]
- Montague, T.G.; Cruz, J.M.; Gagnon, J.A.; Church, G.M.; Valen, E. CHOPCHOP: A CRISPR/Cas9 and TALEN web tool for genome editing. Nucleic Acids Res. 2014, 42, W401–W407. [Google Scholar] [CrossRef]
- Tsai, S.Q.; Zheng, Z.; Nguyen, N.T.; Liebers, M.; Topkar, V.V.; Thapar, V.; Wyvekens, N.; Khayter, C.; Iafrate, A.J.; Le, L.P.; et al. GUIDE-Seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nat. Biotechnol. 2015, 33, 187–197. [Google Scholar] [CrossRef]
- Hoffmann, S.; Mullins, L.; Rider, S.; Brown, C.; Buckley, C.B.; Assmus, A.; Li, Z.; Sierra Beltran, M.; Henderson, N.; Del Pozo, J.; et al. Comparative studies of renin-null zebrafish and mice provide new functional insights. Hypertension 2022, 79, e56–e66. [Google Scholar] [CrossRef]
- Prykhozhij, S.V.; Cordeiro-Santanach, A.; Caceres, L.; Berman, J.N. Genome editing in zebrafish using high-fidelity Cas9 nucleases: Choosing the right nuclease for the task. Methods Mol. Biol. 2020, 2115, 385–405. [Google Scholar]
- Kok, F.O.; Shin, M.; Ni, C.W.; Gupta, A.; Grosse, A.S.; van Impel, A.; Kirchmaier, B.C.; Peterson-Maduro, J.; Kourkoulis, G.; Male, I.; et al. Reverse genetic screening reveals poor correlation between morpholino-induced and mutant phenotypes in zebrafish. Dev. Cell 2015, 32, 97–108. [Google Scholar] [CrossRef]
- Rossi, A.; Kontarakis, Z.; Gerri, C.; Nolte, H.; Holper, S.; Kruger, M.; Stainier, D.Y.R. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature 2015, 524, 230–233. [Google Scholar] [CrossRef]
- Outtandy, P.; Russell, C.; Kleta, R.; Bockenhauer, D. Zebrafish as a model for kidney function and disease. Pediatr. Nephrol. 2019, 34, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Scheer, N.; Campos-Ortega, J.A. Use of the Gal4-UAS technique for targeted gene expression in the zebrafish. Mech. Dev. 1999, 80, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Thisse, C.; Thisse, B. Fast Release Clones: A High Throughput Expression Analysis. ZFIN Direct Data Submission. 2004. Available online: http://zfin.org/ZDB-PUB-040907-1 (accessed on 1 January 2023).
- Thisse, C.; Thisse, B. Expression from: Unexpected Novel Relational Links Uncovered by Extensive Developmental Profiling of Nuclear Receptor Expression. ZFIN Direct Data Submission. 2008. Available online: http://zfin.org/ZDB-PUB-080220-1 (accessed on 1 January 2023).
- Naylor, R.W.; Qubisi, S.S.; Davidson, A.J. Zebrafish pronephros development. Results Probl. Cell Differ. 2017, 60, 27–53. [Google Scholar] [PubMed]
- Li, Y.; Cheng, C.N.; Verdun, V.A.; Wingert, R.A. Zebrafish nephrogenesis is regulated by interactions between retinoic acid, mecom, and Notch signaling. Dev. Biol. 2014, 386, 111–122. [Google Scholar] [CrossRef]
- Drummond, B.E.; Li, Y.; Marra, A.N.; Cheng, C.N.; Wingert, R.A. The tbx2a/b transcription factors direct pronephros segmentation and corpuscle of Stannius formation in zebrafish. Dev. Biol. 2017, 421, 52–66. [Google Scholar] [CrossRef]
- Snelson, C.D.; Santhakumar, K.; Halpern, M.E.; Gamse, J.T. tbx2b is required for the development of the parapineal organ. Development 2008, 135, 1693–1702. [Google Scholar] [CrossRef][Green Version]
- Morales, E.E.; Handa, N.; Drummond, B.E.; Chambers, J.M.; Marra, A.N.; Addiego, A.; Wingert, R.A. Homeogene emx1 is required for nephron distal segment development in zebrafish. Sci. Rep. 2018, 8, 18038. [Google Scholar] [CrossRef]
- Chambers, J.M.; Poureetezadi, S.J.; Addiego, A.; Lahne, M.; Wingert, R.A. ppargc1a controls nephron segmentation during zebrafish embryonic kidney ontogeny. eLife 2018, 7, e40266. [Google Scholar] [CrossRef] [PubMed]
- Reggiani, L.; Raciti, D.; Airik, R.; Kispert, A.; Brandli, A.W. The prepattern transcription factor Irx3 directs nephron segment identity. Genes. Dev. 2007, 21, 2358–2370. [Google Scholar] [CrossRef]
- Naylor, R.W.; Chang, H.G.; Qubisi, S.; Davidson, A.J. A novel mechanism of gland formation in zebrafish involving transdifferentiation of renal epithelial cells and live cell extrusion. eLife 2018, 7, e38911. [Google Scholar] [CrossRef]
- Poureetezadi, S.J.; Cheng, C.N.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Prostaglandin signaling regulates nephron segment patterning of renal progenitors during zebrafish kidney development. eLife 2016, 5, e17551. [Google Scholar] [CrossRef]
- Marra, A.N.; Adeeb, B.D.; Chambers, B.E.; Drummond, B.E.; Ulrich, M.; Addiego, A.; Springer, M.; Poureetezadi, S.J.; Chambers, J.M.; Ronshaugen, M.; et al. Prostaglandin signaling regulates renal multiciliated cell specification and maturation. Proc. Natl. Acad. Sci. USA 2019, 116, 8409–8418. [Google Scholar] [CrossRef]
- Chambers, B.E.; Gerlach, G.F.; Clark, E.G.; Chen, K.H.; Levesque, A.E.; Leshchiner, I.; Goessling, W.; Wingert, R.A. Tfap2a is a novel gatekeeper of nephron differentiation during kidney development. Development 2019, 146, dev172387. [Google Scholar] [CrossRef]
- Chambers, B.E.; Clark, E.G.; Gatz, A.E.; Wingert, R.A. Kctd15 regulates nephron segment development by repressing Tfap2a activity. Development 2020, 147, dev191973. [Google Scholar] [CrossRef]
- Zarelli, V.E.; Dawid, I.B. Inhibition of neural crest formation by Kctd15 involves regulation of transcription factor AP-2. Proc. Natl. Acad. Sci. USA 2013, 110, 2870–2875. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.C.; Rebbert, M.; Wang, C.; Chen, X.; Heffer, A.; Zarelli, V.E.; Dawid, I.B.; Zhao, H. Genes regulated by potassium channel tetramerization domain containing 15 (Kctd15) in the developing neural crest. Int. J. Dev. Biol. 2016, 60, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, C.; Suzuki, T.; Nishida, E.; Kusakabe, M. Identification and characterization of Xenopus kctd15, an ectodermal gene repressed by the FGF pathway. Int. J. Dev. Biol. 2012, 56, 393–402. [Google Scholar] [CrossRef]
- Dutta, S.; Dawid, I.B. Kctd15 inhibits neural crest formation by attenuating Wnt/β-catenin signaling output. Development 2010, 137, 3013–3018. [Google Scholar] [CrossRef]
- Chambers, J.M.; Addiego, A.; Flores-Mireles, A.L.; Wingert, R.A. Ppargc1a controls ciliated cell development by regulating prostaglandin biosynthesis. Cell Rep. 2020, 33, 108370. [Google Scholar] [CrossRef]
- Marra, A.N.; Cheng, C.N.; Adeeb, B.; Addiego, A.; Wesselman, H.M.; Chambers, B.E.; Chambers, J.M.; Wingert, R.A. Iroquois transcription factor irx2a is required for multiciliated and transporter cell fate decisions during zebrafish pronephros development. Sci. Rep. 2019, 9, 6454. [Google Scholar] [CrossRef]
- Weaver, N.E.; Healy, A.; Wingert, R.A. gldc is essential for renal progenitor patterning during kidney development. Biomedicines 2022, 10, 3220. [Google Scholar] [CrossRef] [PubMed]
- Wesselman, H.M.; Gatz, A.E.; Pfaff, M.R.; Arceri, L.; Wingert, R.A. Estrogen signaling influences nephron segmentation of the zebrafish embryonic kidney. Cells 2023, 12, 666. [Google Scholar] [CrossRef]
- Tang, Q.; Iyer, S.; Lobbardi, R.; Moore, J.C.; Chen, H.; Lareau, C.; Hebert, C.; Shaw, M.L.; Neftel, C.; Suva, M.L.; et al. Dissecting hematopoietic and renal cell heterogeneity in adult zebrafish at single-cell resolution using RNA sequencing. J. Exp. Med. 2017, 214, 2875–2887. [Google Scholar] [CrossRef] [PubMed]
- Sugano, Y.; Cianciolo Cosentino, C.; Loffing-Cueni, D.; Neuhauss, S.C.F.; Loffing, J. Comparative transcriptomic analysis identifies evolutionarily conserved gene products in the vertebrate renal distal convoluted tubule. Pflügers Arch.-Eur. J. Physiol. 2017, 469, 859–867. [Google Scholar] [CrossRef]
- Sander, V.; Salleh, L.; Naylor, R.W.; Schierding, W.; Sontam, D.; O’Sullivan, J.M.; Davidson, A.J. Transcriptional profiling of the zebrafish proximal tubule. Am. J. Physiol. Renal Physiol. 2019, 317, F478–F488. [Google Scholar] [CrossRef]
- Brown, C.; Mullins, L.J.; Wesencraft, K.; McConnell, G.; Beltran, M.; Henderson, N.C.; Conway, B.; Hoffmann, S.; Rider, S.; Mullins, J.J. scRNA transcription profile of adult zebrafish podocytes using a novel reporter strain. Cell Physiol. Biochem. 2021, 55, 35–47. [Google Scholar] [PubMed]
- Corkins, M.E.; Achieng, M.; DeLay, B.D.; Krneta-Stankic, V.; Cain, M.P.; Walker, B.L.; Chen, J.; Lindström, N.O.; Miller, R.K. A comparative study of cellular diversity between the Xenopus pronephric and mouse metanephric nephron. Kidney Int. 2023, 103, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Chambers, B.E.; Weaver, N.E.; Wingert, R.A. The “3Ds” of Growing Kidney Organoids: Advances in Nephron Development, Disease Modeling, and Drug Screening. Cells 2023, 12, 549. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.K.; Petrikas, M.; Chambers, B.E.; Wingert, R.A. Principles of Zebrafish Nephron Segment Development. J. Dev. Biol. 2023, 11, 14. https://doi.org/10.3390/jdb11010014
Nguyen TK, Petrikas M, Chambers BE, Wingert RA. Principles of Zebrafish Nephron Segment Development. Journal of Developmental Biology. 2023; 11(1):14. https://doi.org/10.3390/jdb11010014
Chicago/Turabian StyleNguyen, Thanh Khoa, Madeline Petrikas, Brooke E. Chambers, and Rebecca A. Wingert. 2023. "Principles of Zebrafish Nephron Segment Development" Journal of Developmental Biology 11, no. 1: 14. https://doi.org/10.3390/jdb11010014
APA StyleNguyen, T. K., Petrikas, M., Chambers, B. E., & Wingert, R. A. (2023). Principles of Zebrafish Nephron Segment Development. Journal of Developmental Biology, 11(1), 14. https://doi.org/10.3390/jdb11010014