Role of Cop9 Signalosome Subunits in the Environmental and Hormonal Balance of Plant


Abstract
1. Introduction
2. Role of CSN in Hormonal Signaling
3. Importance of CSN in Biotic and Abiotic Stress
4. Characteristics of CSN Hypomorphic Mutants and Their Importance to Study Plant Growth and Development
4.1. csn1-10
4.2. csn2-5
4.3. csn3-3
4.4. csn4-2035
4.5. csn5a-1, csn5a-2 and csn5b-1
4.6. csn6a-1 and csn6b-1
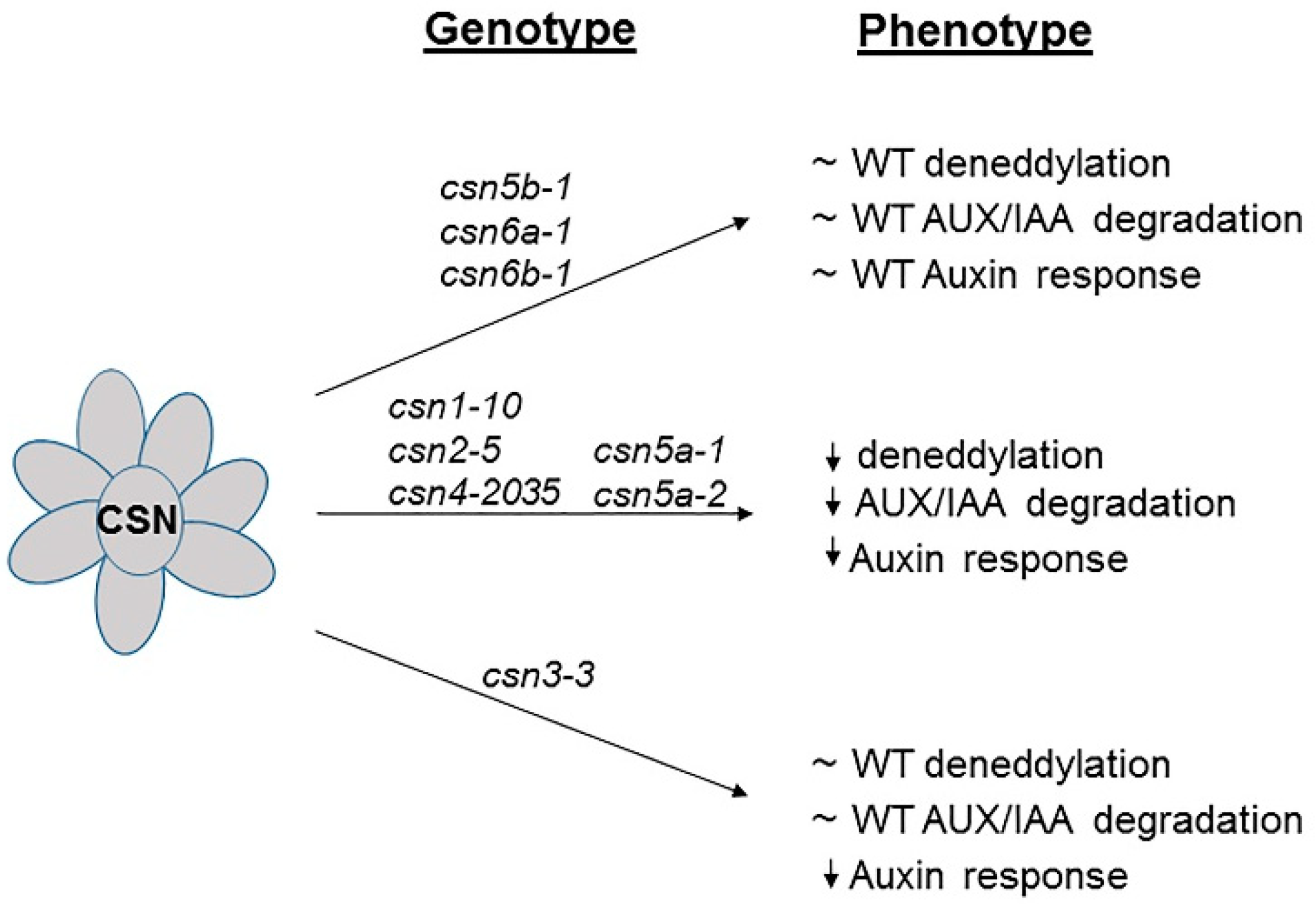
5. Comparison of Hypomorphic Mutants on the Basis of Developmental and Deneddylation Activity
6. Conclusions and Future Perspective
Funding
Conflicts of Interest
References
- Chamovitz, D.A.; Wei, N.; Osterlund, M.T.; Von Arnim, A.G.; Staub, J.M.; Matsui, M.; Deng, X.-W. The COP9 Complex, a Novel Multisubunit Nuclear Regulator Involved in Light Control of a Plant Developmental Switch. Cell 1996, 86, 115–121. [Google Scholar] [CrossRef]
- Wei, N.; Chamovitz, D.A.; Deng, X.-W. Arabidopsis COP9 is a component of a novel signaling complex mediating light control of development. Cell 1994, 78, 117–124. [Google Scholar] [CrossRef]
- Kim, T.-H.; Hofmann, K.; Von Arnim, A.G.; Chamovitz, D.A. PCI complexes: pretty complex interactions in diverse signaling pathways. Trends Plant Sci. 2001, 6, 379–386. [Google Scholar] [CrossRef]
- Schwechheimer, C. Interactions of the COP9 Signalosome with the E3 Ubiquitin Ligase SCFTIR1 in Mediating Auxin Response. Science 2001, 292, 1379–1382. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Serino, G.; Deng, X.W. A role of Arabidopsis COP9 signalosome in multifaceted developmental processes revealed by the characterization of its subunit 3. Development 2001, 128, 4277–4288. [Google Scholar] [PubMed]
- Wang, X.; Feng, S.; Nakayama, N.; Crosby, W.L.; Irish, V.; Deng, X.W.; Wei, N. The COP9 Signalosome Interacts with SCFUFO and Participates in Arabidopsis Flower Development. Plant Cell 2003, 15, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Hind, S.; Pulliam, S.E.; Veronese, P.; Shantharaj, D.; Nazir, A.; Jacobs, N.S.; Stratmann, J.W. The COP9 signalosome controls jasmonic acid synthesis and plant responses to herbivory and pathogens. Plant J. 2011, 65, 480–491. [Google Scholar] [CrossRef]
- Meir, M.; Galanty, Y.; Kashani, L.; Blank, M.; Khosravi, R.; Fernández-Ávila, M.J.; Cruz-García, A.; Star, A.; Shochot, L.; Thomas, Y.; et al. The COP9 signalosome is vital for timely repair of DNA double-strand breaks. Nucleic Acids Res. 2015, 43, 4517–4530. [Google Scholar] [CrossRef]
- Groisman, R.; Polanowska, J.; Kuraoka, I.; Sawada, J.-I.; Saijo, M.; Drapkin, R.; Kisselev, A.F.; Tanaka, K.; Nakatani, Y. The Ubiquitin Ligase Activity in the DDB2 and CSA Complexes Is Differentially Regulated by the COP9 Signalosome in Response to DNA Damage. Cell 2003, 113, 357–367. [Google Scholar] [CrossRef]
- Halimi, Y.; Dessau, M.; Pollak, S.; Ast, T.; Erez, T.; Livnat-Levanon, N.; Karniol, B.; Hirsch, J.A.; Chamovitz, D.A. COP9 signalosome subunit 7 from Arabidopsis interacts with and regulates the small subunit of ribonucleotide reductase (RNR2). Plant Mol. Boil. 2011, 77, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Gusmaroli, G.; Figueroa, P.; Serino, G.; Deng, X.W. Role of the MPN Subunits in COP9 Signalosome Assembly and Activity, and Their Regulatory Interaction with Arabidopsis Cullin3-Based E3 Ligases. Plant Cell 2007, 19, 564–581. [Google Scholar] [CrossRef] [PubMed]
- Dohmann, E.M.N.; Levesque, M.P.; De Veylder, L.; Reichardt, I.; Jürgens, G.; Schmid, M.; Schwechheimer, C. The Arabidopsis COP9 signalosome is essential for G2 phase progression and genomic stability. Dev. 2008, 135, 2013–2022. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Ma, L.; Wang, X.; Xie, D.; Dinesh-Kumar, S.P.; Wei, N.; Deng, X.W. The COP9 Signalosome Interacts Physically with SCFCOI1 and Modulates Jasmonate Responses. Plant Cell 2003, 15, 1083–1094. [Google Scholar] [CrossRef]
- Schwechheimer, C. The COP9 signalosome (CSN): an evolutionary conserved proteolysis regulator in eukaryotic development. Biochim Biophys ACTA 2004, 1695, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Martínez, C.; Gusmaroli, G.; Wang, Y.; Zhou, J.; Wang, F.; Chen, L.; Yu, L.; Iglesias-Pedraz, J.M.; Kircher, S.; et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins. Nature 2008, 451, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Christians, M.J.; Robles, L.M.; Zeller, S.M.; Larsen, P.B. The eer5 mutation, which affects a novel proteasome-related subunit, indicates a prominent role for the COP9 signalosome in resetting the ethylene-signaling pathway in Arabidopsis. Plant J. 2008, 55, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Wu, M.; Li, B.; Bücker, B.; Keil, P.; Zhang, S.; Li, J.; Kang, D.; Liu, J.; Dong, J.; et al. The COP9 Signalosome regulates seed germination by facilitating protein degradation of RGL2 and ABI5. PLoS Genet. 2018, 14, e1007237. [Google Scholar] [CrossRef]
- Fu, X.; Richards, D.E.; Fleck, B.; Xie, D.; Burton, N.; Harberd, N.P. The Arabidopsis Mutant sleepy1gar2-1 Protein Promotes Plant Growth by Increasing the Affinity of the SCFSLY1 E3 Ubiquitin Ligase for DELLA Protein Substrates. Plant Cell 2004, 16, 1406–1418. [Google Scholar] [CrossRef]
- Dill, A.; Thomas, S.G.; Hu, J.; Steber, C.M.; Sun, T.-P. The Arabidopsis F-Box Protein SLEEPY1 Targets Gibberellin Signaling Repressors for Gibberellin-Induced Degradation. Plant Cell 2004, 16, 1392–1405. [Google Scholar] [CrossRef]
- Lozano-Durán, R.; Rosas-Díaz, T.; Gusmaroli, G.; Luna, A.P.; Taconnat, L.; Deng, X.W.; Bejarano, E.R. Geminiviruses Subvert Ubiquitination by Altering CSN-Mediated Derubylation of SCF E3 Ligase Complexes and Inhibit Jasmonate Signaling in Arabidopsis thaliana. Plant Cell 2011, 23, 1014–1032. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Nezames, C.D.; Sheng, L.; Deng, X.; Wei, N. Cullin-RING Ubiquitin Ligase Family in Plant Abiotic Stress PathwaysF. J. Integr. Plant Boil. 2013, 55, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.; Ma, S.L.; Bai, L.P.; Zhang, L.; Ma, H.; Jia, P.; Liu, J.; Zhong, M.; Guo, Z.F. Signal transduction during cold, salt, and drought stresses in plants. Mol. Biol. Rep. 2012, 39, 969–987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-Y.; Lei, G.; Zhou, H.-W.; He, C.; Liao, J.-L.; Huang, Y.-J. Quantitative iTRAQ-based proteomic analysis of rice grains to assess high night temperature stress. Proteomics 2017, 17, 1600365. [Google Scholar] [CrossRef] [PubMed]
- Serino, G.; Deng, X. THE COP9 S IGNALOSOME: Regulating Plant Development Through the Control of Proteolysis. Annu. Rev. Plant Biol. 2003, 54, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Oron, E.; Tuller, T.; Li, L.; Rozovsky, N.; Yekutieli, D.; Segal, D.; Chor, B.; Edgar, B.A.; Chamovitz, D.A.; Rencus-Lazar, S.; et al. Genomic analysis of COP9 signalosome function in Drosophila melanogaster reveals a role in temporal regulation of gene expression. Mol. Syst. Boil. 2007, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Seibert, V.; Geyer, R.; Rhee, E.; Lyapina, S.; Cope, G.; Deshaies, R.J.; Wolf, D.A. The fission yeast COP9/signalosome is involved in cullin modification by ubiquitin-related Ned8p. BMC Biochem. 2001, 2, 7. [Google Scholar] [CrossRef][Green Version]
- Zhang, W.; Ito, H.; Quint, M.; Huang, H.; Noël, L.D.; Gray, W.M. Genetic analysis of CAND1–CUL1 interactions in Arabidopsis supports a role for CAND1-mediated cycling of the SCFTIR1 complex. Proc. Natl. Acad. Sci. 2008, 105, 8470–8475. [Google Scholar] [CrossRef] [PubMed]
- Stuttmann, J.; Lechner, E.; Guérois, R.; Nussaume, L.; Genschik, P.; Parker, J.E.; Noel, L.D. COP9 signalosome- and 26S proteasome-dependent regulation of SCFTIR1 accumulation in Arabidopsis. J. Boil. Chem. 2009, 284, 7920–7930. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Quint, M.; Gray, W.M. The eta7/csn3-3 Auxin Response Mutant of Arabidopsis Defines a Novel Function for the CSN3 Subunit of the COP9 Signalosome. PLoS ONE 2013, 8, e66578. [Google Scholar] [CrossRef] [PubMed]
- Pacurar, D.I.; Pacurar, M.L.; Lakehal, A.; Pacurar, A.M.; Ranjan, A.; Bellini, C. The Arabidopsis Cop9 signalosome subunit 4 (CSN4) is involved in adventitious root formation. Sci. Rep. 2017, 7, 261. [Google Scholar]
- Kotiguda, G.G.; Weinberg, D.; Dessau, M.; Salvi, C.; Serino, G.; Chamovitz, D.A.; Hirsch, J.A. The Organization of a CSN5-containing Subcomplex of the COP9 Signalosome. J. Boil. Chem. 2012, 287, 42031–42041. [Google Scholar] [CrossRef] [PubMed]
- Kwok, S.F.; Solano, R.; Tsuge, T.; Chamovitz, D.A.; Ecker, J.R.; Matsui, M.; Deng, X.-W. Arabidopsis Homologs of a c-Jun Coactivator Are Present Both in Monomeric Form and in the COP9 Complex, and Their Abundance Is Differentially Affected by the Pleiotropic cop/det/fus Mutations. Plant Cell 2007, 10, 1779. [Google Scholar] [CrossRef] [PubMed]
- Dohmann, E.M.; Kuhnle, C.; Schwechheimer, C. Loss of the CONSTITUTIVE PHOTOMORPHOGENIC9 Signalosome Subunit 5 Is Sufficient to Cause the cop/det/fus Mutant Phenotype in Arabidopsis. Plant Cell 2005, 17, 1967–1978. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Hypomorphic Mutants | Ecotype | Details of Mutation and Phenotype | Deneddylation Activity | References |
|---|---|---|---|---|
| csn1-10 | Col-0 | Transition mutation resulting in Ser-Asn substitution at amino acid 305. Mild dwarf phenotype. Show auxin-resistant root growth and reduced SCFTIR1 activity. | Affects | [29] |
| csn2-5 | Ler | Single base pair exchange in codon GGT-GAT results in amino acid substitution mutation from Gly-Asp at position 237 in CSN2. Hypomorphic mutant, viable and fertilize. Mild dwarf phenotype. Show auxin-resistant root growth and defective in Aux/IAA protein degradation. | Affects | [30] |
| csn3-3 | Col-0 | Missense mutation in Gly-293 to Glu of PCI domain of CSN3. Exhibit auxin related response like auxin-resistant root growth. But unlike csn mutants, it is not defective in Aux/IAA protein degradation. | Does not affect | [31] |
| csn4-2035 | Ws-4 | Point mutation of G to A at position 2592 in the tenth exon of CSN4 results in Ala-302 to Val amino acid substitution. Slightly resistant to exogenously applied auxin. Inefficient degradation of AUX/IAA protein. Produced similar amount of CSN4 protein as the wild type. Dwarf phenotype with reduction in the number of adventitious root formation. | Partially affects | [32] |
| csn5a-1 | Col-0 | T-DNA insertion (SALK_063436). Severe dwarf phenotype. Auxin-resistant root growth. | Affects | [35] |
| csn5a-2 | Col-0 | T-DNA insertion (SALK_027705). Dwarf phenotype. Affects deneddylation activity. Auxin and jasmonic acid resistant root growth. | Affects | [35] |
| csn5b-1 | Col-0 | T-DNA insertion (SALK_007134). Phenotype almost similar to wild type (Col-0). Auxin and jasmonic acid sensitive root growth. | Does not affect | [35] |
| csn6a-1 and csn6b-1 | Col-0 | T-DNA insertion SALK_146926 (csn6a-1), and SALK_036965 (csn6b-1), In the dark, loss of CSN6A affect hypocotyl elongation slightly more than loss of CSN6B. But both the mutants do not display any obvious morphological defects in white light. | Do not affect | [12] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.K.; Chamovitz, D.A. Role of Cop9 Signalosome Subunits in the Environmental and Hormonal Balance of Plant. Biomolecules 2019, 9, 224. https://doi.org/10.3390/biom9060224
Singh AK, Chamovitz DA. Role of Cop9 Signalosome Subunits in the Environmental and Hormonal Balance of Plant. Biomolecules. 2019; 9(6):224. https://doi.org/10.3390/biom9060224
Chicago/Turabian StyleSingh, Amit Kumar, and Daniel A. Chamovitz. 2019. "Role of Cop9 Signalosome Subunits in the Environmental and Hormonal Balance of Plant" Biomolecules 9, no. 6: 224. https://doi.org/10.3390/biom9060224
APA StyleSingh, A. K., & Chamovitz, D. A. (2019). Role of Cop9 Signalosome Subunits in the Environmental and Hormonal Balance of Plant. Biomolecules, 9(6), 224. https://doi.org/10.3390/biom9060224