
N-acetyl-l-histidine, a Prominent Biomolecule in Brain and Eye of Poikilothermic Vertebrates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Background
1.2. NAH Metabolism
1.3. NAH Molecular Water Pump Hypothesis
2. Materials and Methods
2.1. 1HMRS Methods
2.2. Phantoms
2.3. Animals and Tissues
3. Results
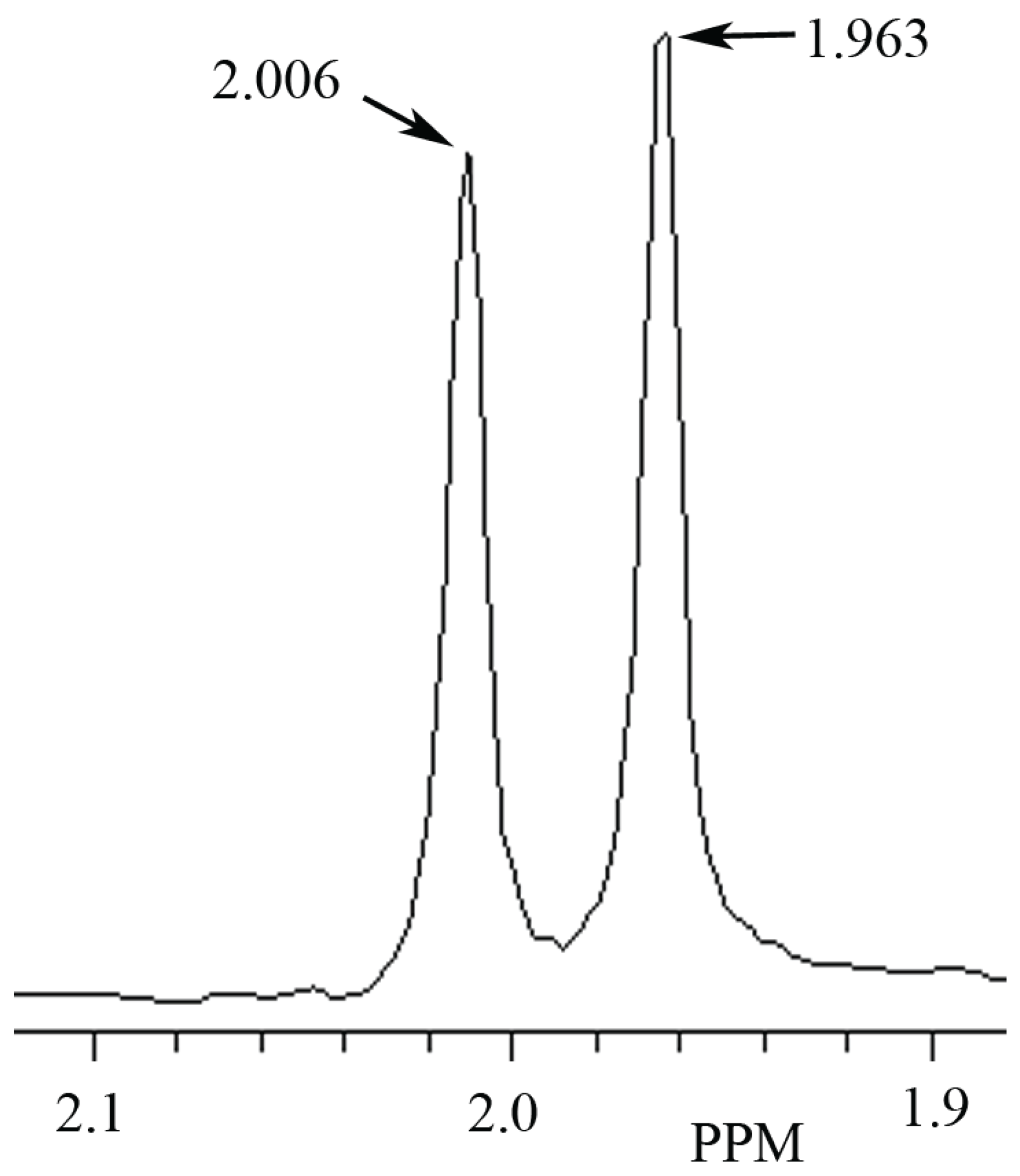
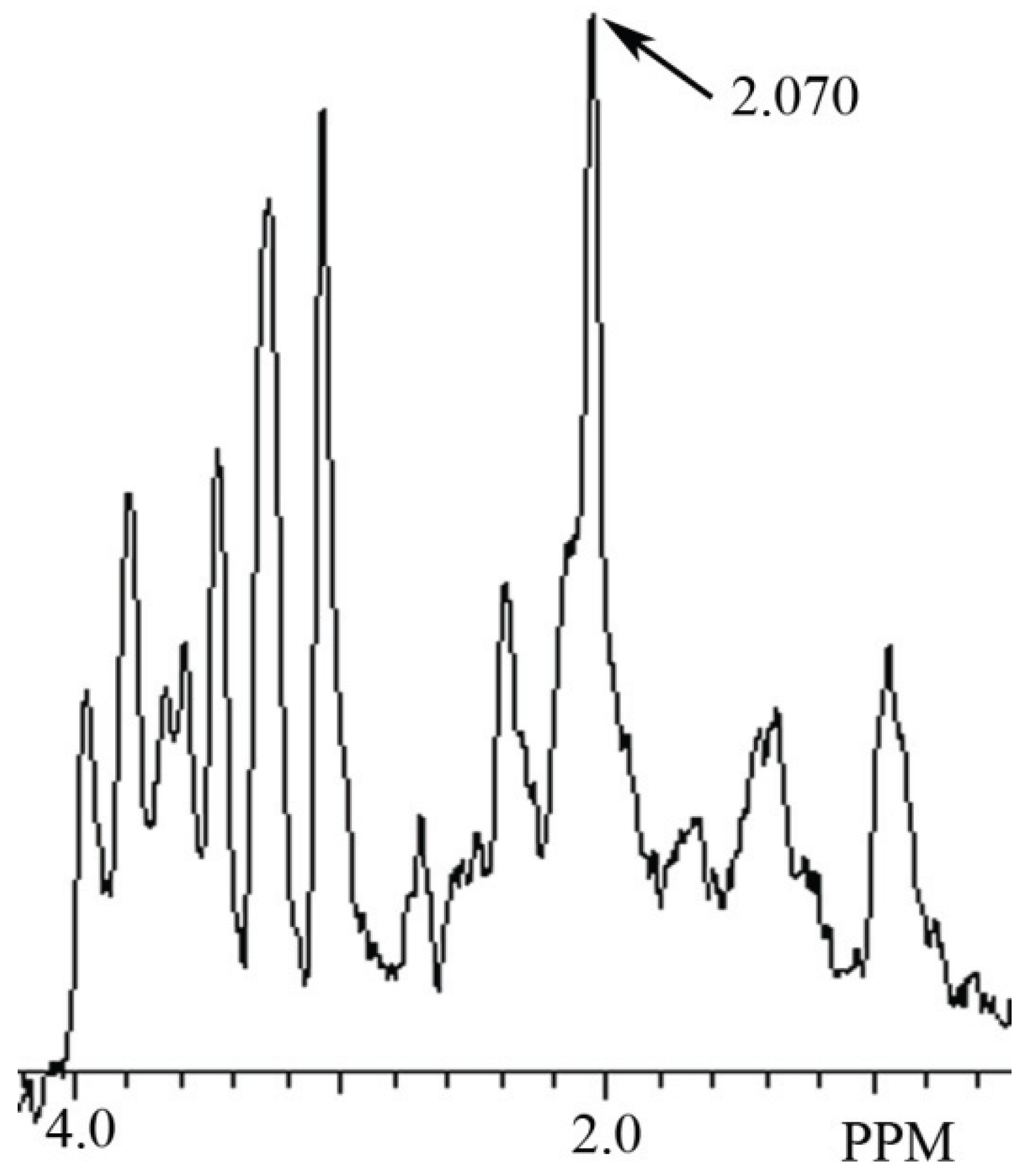
3.1. Identification ofNAA and NAH in Phantoms Using MRS
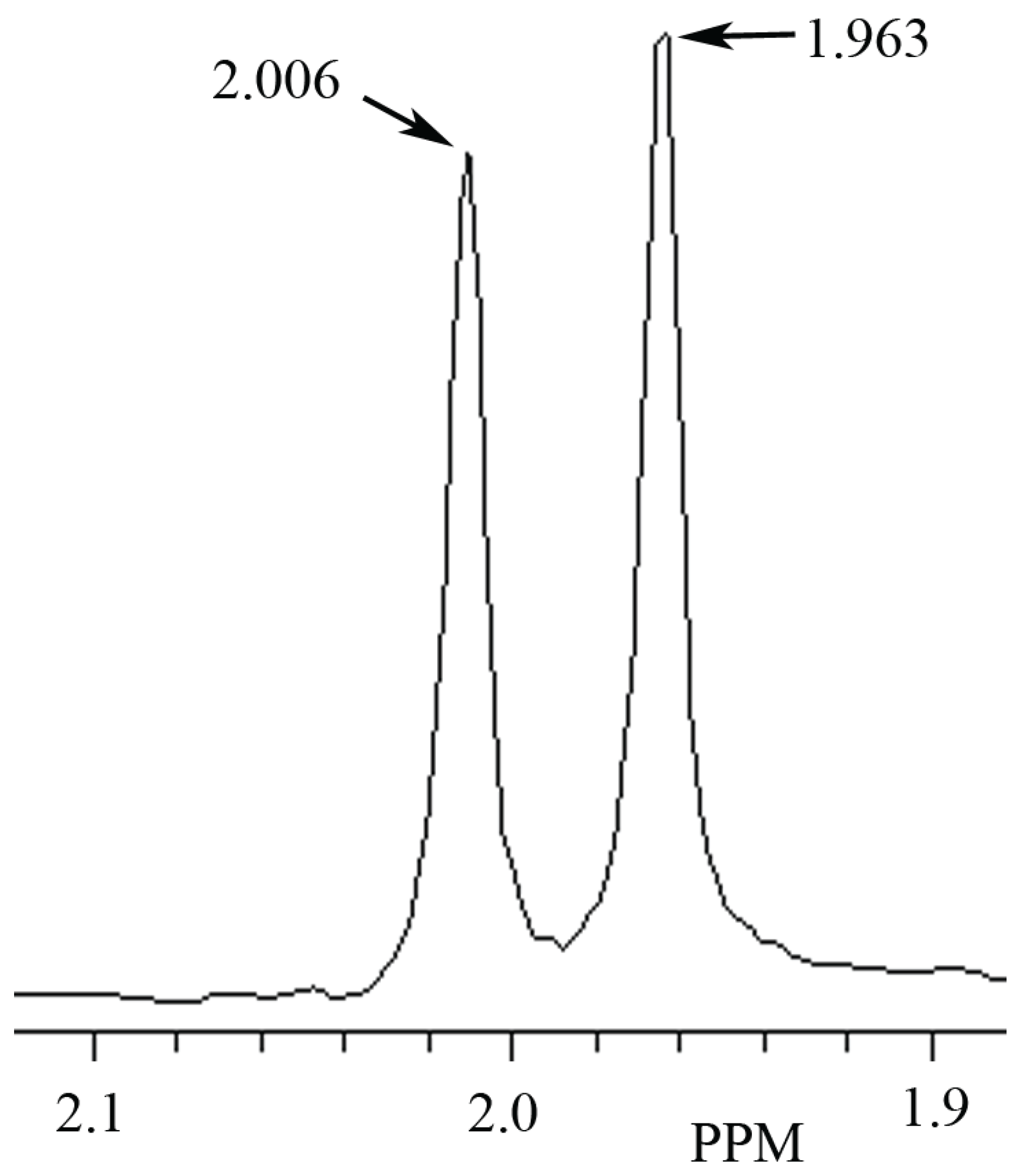
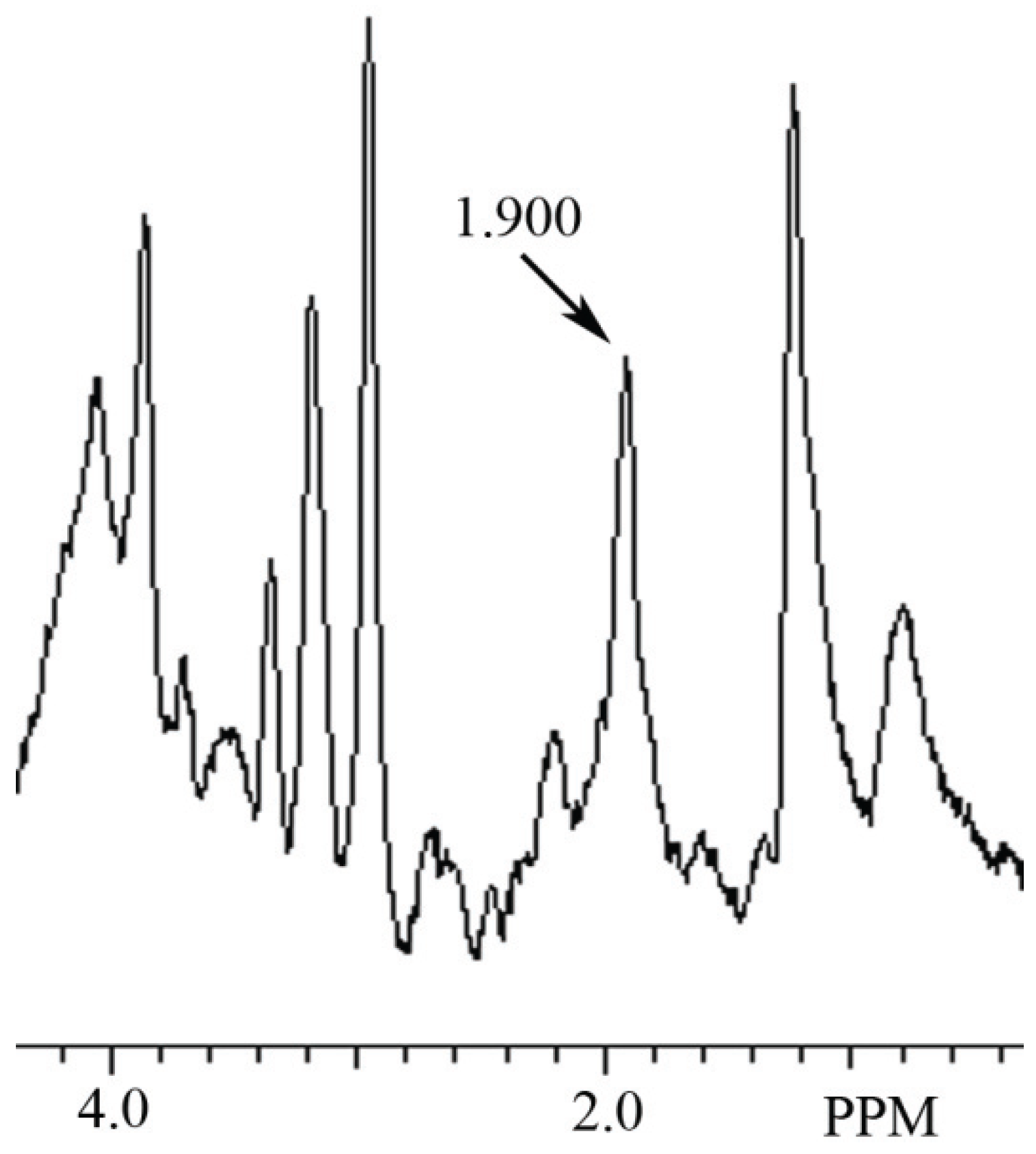
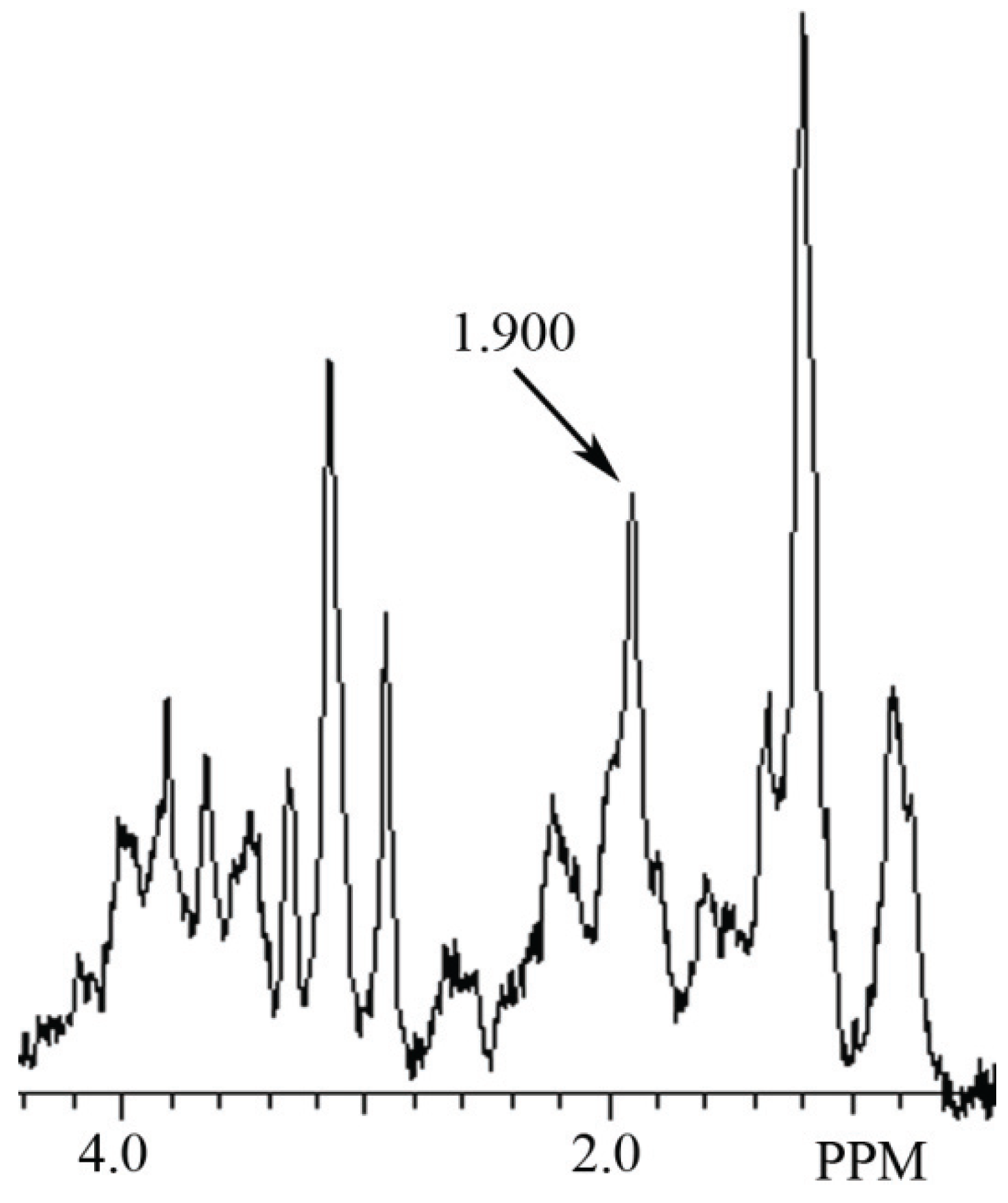
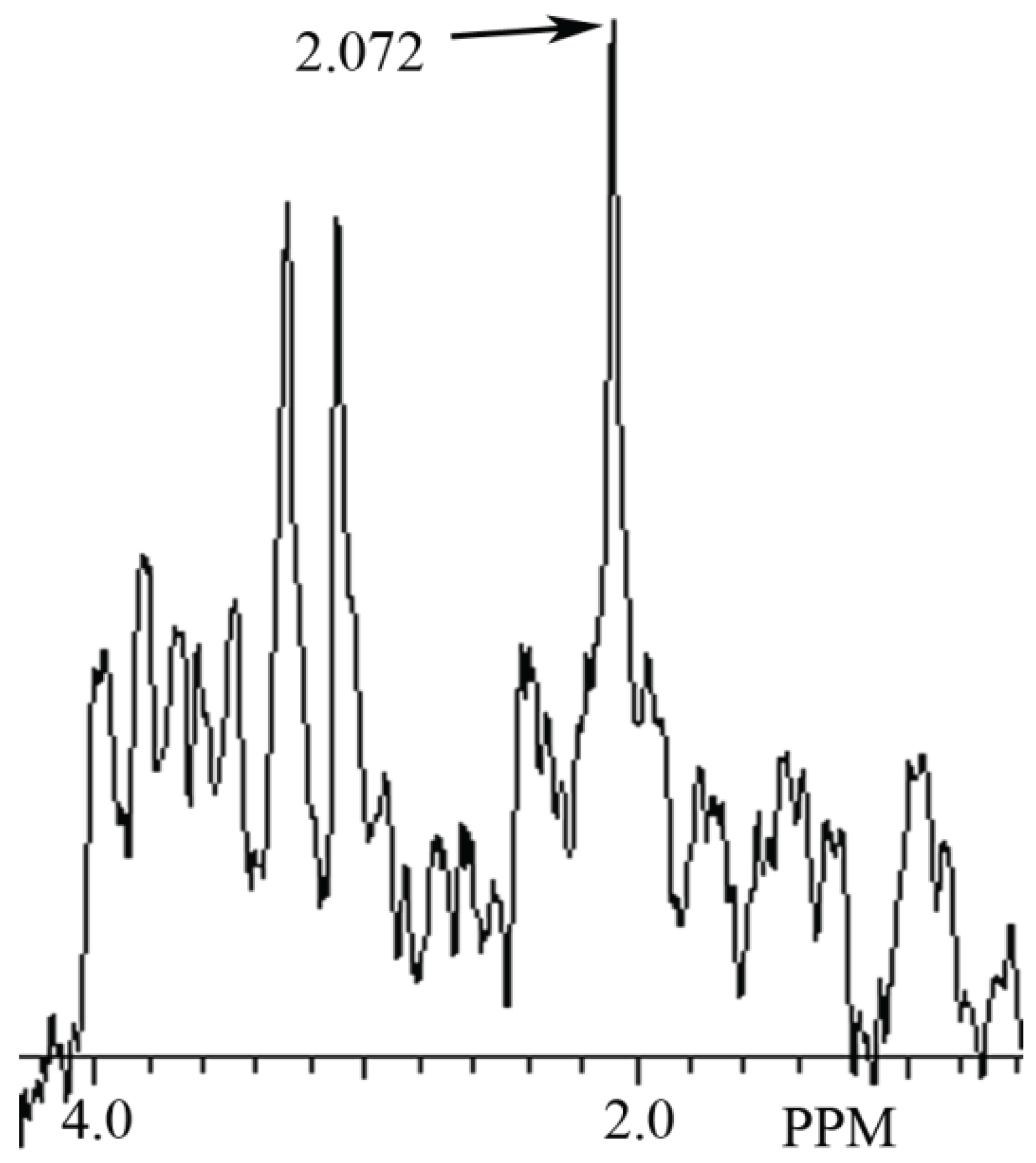
3.2. Identification of NAH in Fish Brain, and NAA in Mammalian Brain using MRS
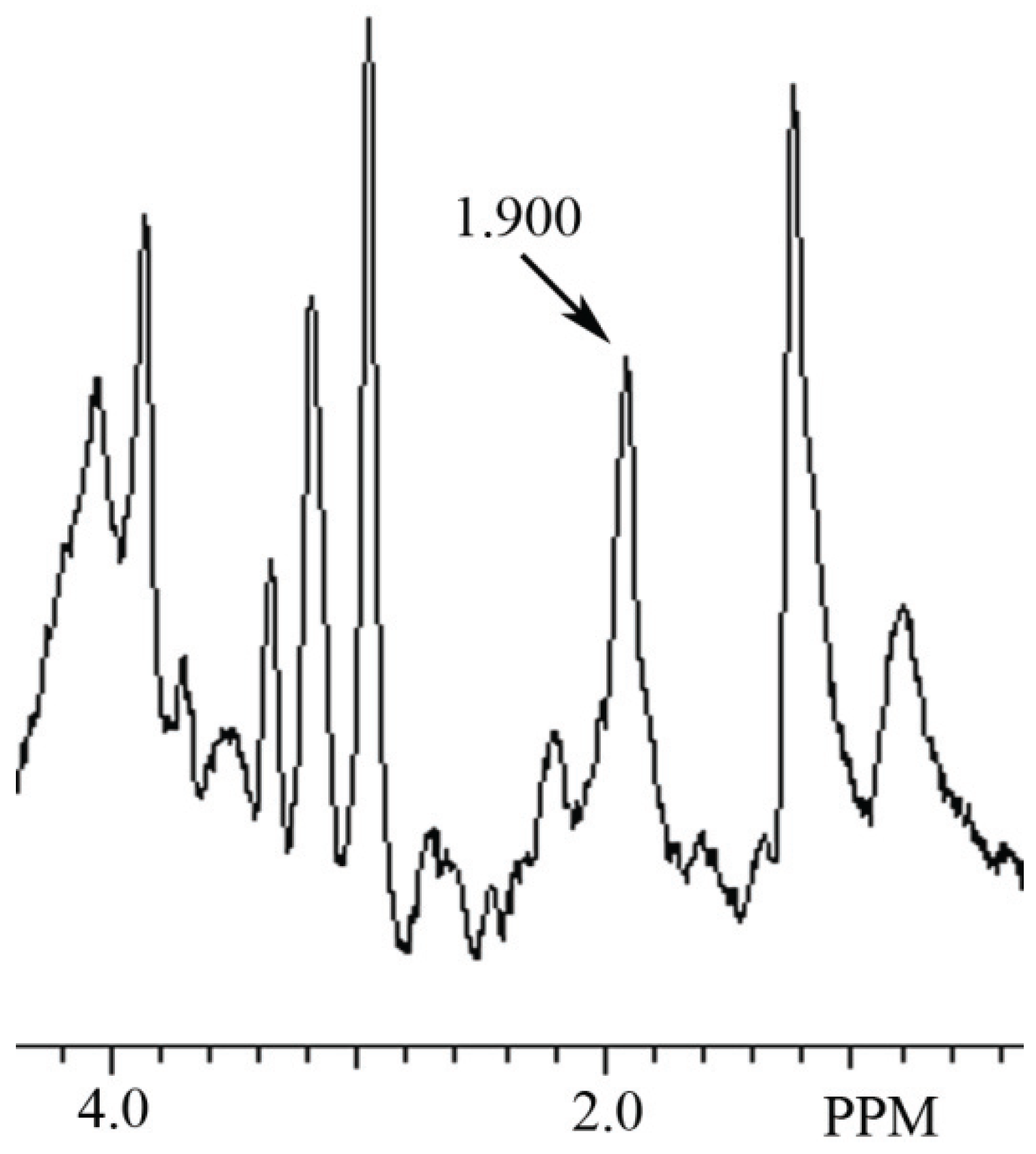
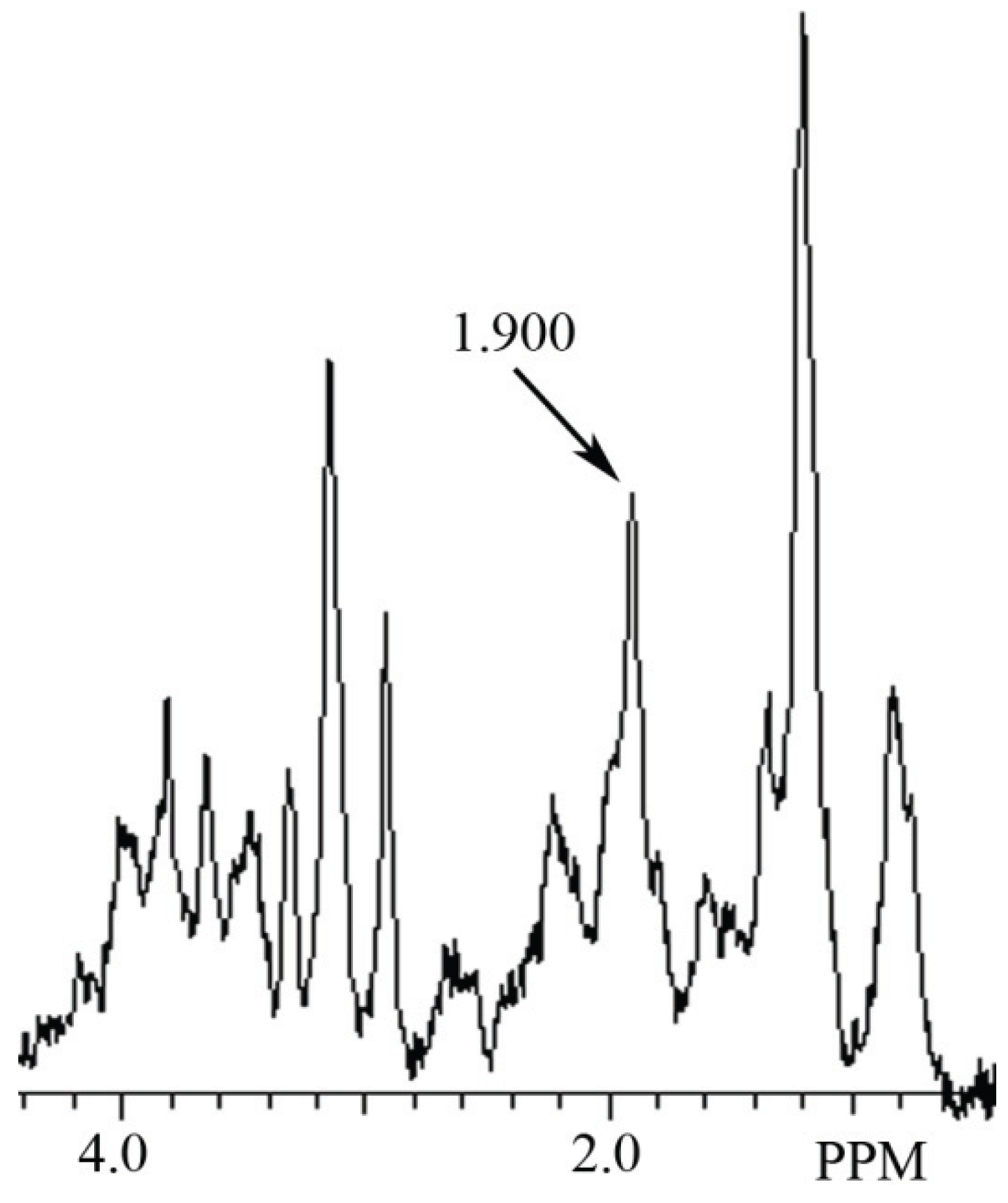


4. Discussion
4.1. The Use of MRS to Determine the Presence of NAH in Fish Brain
4.2. Recent Evidence in Support of the NAH Efflux MWP Hypothesis in Fish Lens and Eye
5. Conclusions
5.1. Identification of NAH in Fish Brain Using MRS
5.2. Support for the NAH MWP Hypothesis
5.3. A Problem of Metabolite Identification and Quantification when Using Automated MRS Programs in Different Species
5.4. Retrospective Identification of NAH in Published Fish Brain MRS Studies
5.5. The Thermal Dimension in Poikilotherms Provides a Unique Investigative Tool to Study the Physiological Functions of NAH and NAA
5.6. A Possible Dietary Approach for Treatment of some Brain and Eye Disorders in Homeotherms
Author Contributions
Abbreviations
| Ac | Acetate |
| AcCoA | Acetyl Co-enzyme A |
| Carn | Carnosine |
| ECF | Extra-cellular fluid |
| Glc | d-glucose |
| His | l-histidine |
| Hcarn | Homocarnosine |
| HPLC | High pressure liquid chromatography |
| MWP | Molecular water pump |
| MRS | Magnetic resonance spectroscopy |
| NAA | N-acetyl-l-aspartate |
| NAH | N-acetyl-l-histidine |
Conflicts of Interest
References
- Baslow, M.H. A review of phylogenetic and metabolic relationships between the acylamino acids, N-acetyl-l-aspartic acid and N-acetyl-l-histidine in the vertebrate nervous system. J. Neurochem. 1997, 68, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H. N-acetyl-l-histidine synthetase activity from the brain of the killifish. Brain Res. 1966, 3, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Arikawa, S. An ectotherm homologue of human predicted gene NAT16 encodes histidine N-acetyltransferase responsible for Nα-acetylhistidine synthesis. Biochim. Biophys. Acta 2014, 1840, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H. N-acetyl-l-histidine metabolism in the fish eye: Evidence for ocular fluid-lens l-histidine recycling. Exp. Eye Res. 1967, 6, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H.; Lenney, J.F. α-N-acetyl-l-histidine amidohydrolase activity from the brain of the skipjack tuna Katsuwonus pelamis. Can. J. Biochem. 1967, 45, 337–340. [Google Scholar]
- Lenney, J.F.; Baslow, M.H.; Sugiyama, G.H. Similarity of tuna N-acetylhistidine deacetylase and cod fish anserinase. Comp. Biochem. Physiol. 1978, 61B, 253–258. [Google Scholar]
- Yamada, S.; Tanaka, Y.; Sameshima, M.; Furuichi, M. Properties of Nα-acetylhistidine deacetylase in brain of trout Oncorhynchus mykiss. Comp. Biochem. Physiol. B 1993, 106, 309–315. [Google Scholar] [CrossRef]
- Baslow, M.H.; Yamada, S. Identification of N-acetylaspartate in the lens of the vertebrate eye: A new model for the investigation of the function of N-acetylated amino acids in vertebrates. Exp. Eye Res. 1997, 64, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H. Function of the N-acetyl-l-histidine system in the vertebrate eye. Evidence in support of a role as a molecular water pump. J. Mol. Neurosci. 1998, 10, 193–208. [Google Scholar]
- Meinild, A.-K.; Klaerke, D.A.; Loo, D.D.F.; Wright, E.M.; Zeuthen, T. The human Na+-glucose cotransporter is a molecular water pump. J. Physiol. 1998, 508, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H. The existence of molecular water pumps in the nervous system. A review of the evidence. Neurochem. Int. 1999, 34, 77–90. [Google Scholar] [CrossRef]
- Loo, D.D.F.; Wright, E.M.; Zeuthen, T. Water pumps. J. Physiol. 2002, 542, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Choe, S.; Rosenberg, J.M.; Abramson, J.; Wright, E.M.; Grabe, M. Water permeation through sodium-dependent galactose cotransporter vSGLT. Biophys. J. 2010, 99, L56–L58. [Google Scholar] [CrossRef] [PubMed]
- Swietach, P.; Leem, C.-H.; Spitzer, K.W.; Vaughan-Jones, R.D. Pumping Ca2+ up H+gradients: A Ca2+-H+ exchanger without a membrane. J. Physiol. 2014, 592, 3179–3188. [Google Scholar] [CrossRef] [PubMed]
- Kabli, S.; Spaink, H.P.; de Groot, H.J.M.; Alia, A. In vivo metabolite profile of adult zebrafish brain obtained by high-resolution localized magnetic resonance spectroscopy. J. Magn. Reson. Imaging 2009, 29, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Provencher, S.W. Automatic quantification of localized in vivo 1H spectra with LCModel. NMR Biomed. 2001, 14, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H.; Ruggieri, G.D. N-acetylhistidine in developing embryos of the killifish Fundulus heteroclitus and the zebrafish Brachydanio rerio. Life Sci. 1967, 6, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Wang, J.; Li, M.; Liu, Q.; Wei, D.; Yang, M.; Kong, L. 1H-NMR-based metabolomics study on a goldfish model of Parkinson’s disease induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Chem. Biol. Interact. 2014, 223, 18–26. [Google Scholar] [CrossRef]
- Breck, O.; Bjerkas, E.; Campbell, P.; Rhodes, J.D.; Sanderson, J.; Waagbo, R. Histidine nutrition and genotype affect cataract development in Atlantic salmon, Salmo salar L. J. Fish. Dis. 2005, 28, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Trosse, C.; Waagbo, R.; Breck, O.; Stavrum, A.-K.; Petersen, K.; Olsvik, P.A. Genome-wide transcription analysis of histidine-related cataract in Atlantic salmon (Salmo salar L). Mol. Vis. 2009, 15, 1332–1350. [Google Scholar] [PubMed]
- Rhodes, J.D.; Breck, O.; Waagbo, R.; Bjerkas, E.; Sanderson, J. N-acetylhistidine, a novel osmolyte in the lens of the Atlantic salmon (Salmo salar L.). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, 1075–1081. [Google Scholar] [CrossRef]
- Remo, S.C.; Olsvik, P.A.; Torstensen, B.E.; Amlund, H.; Breck, O.; Waagbo, R. Susceptibility of Atlantic salmon lenses to hydrogen peroxide oxidation ex vivo after being fed diets with vegetable oil and methylmercury. Exp. Eye Res. 2011, 92, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Remo, S.C.; Hevroy, E.M.; Olsvik, P.A.; Fontanillas, R.; Breck, O.; Waagbo, R. Dietary histidine requirement to reduce the risk and severity of cataracts is higher than the requirement for growth in Atlantic salmon smolts, independently of the dietary lipid source. Br. J. Nutr. 2014, 111, 1759–1772. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H.; Hrabe, J.; Guilfoyle, D.N. Dynamic relationship between neurostimulation and N-acetylaspartate metabolism in the human visual cortex. J. Mol. Neurosci. 2007, 32, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Baslow, M.H. Temperature adaptation and the nervous system of fish. In Molecular Mechanisms of Temperature Adaptation; Prosser, C.L., Ed.; AAAS Symposium: Washington, D.C., USA, 1967; Volume 84, pp. 205–226. [Google Scholar]
- Baslow, M.H.; Suckow, R.F.; Berg, M.J.; Marks, N.; Saito, M.; Bhakoo, K.K. Differential expression of carnosine, homocarnosine and N-acetyl-l-histidine hydrolytic activities in cultured rat macroglial cells. J. Mol. Neurosci. 2001, 17, 87–95. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baslow, M.H.; Guilfoyle, D.N. N-acetyl-l-histidine, a Prominent Biomolecule in Brain and Eye of Poikilothermic Vertebrates. Biomolecules 2015, 5, 635-646. https://doi.org/10.3390/biom5020635
Baslow MH, Guilfoyle DN. N-acetyl-l-histidine, a Prominent Biomolecule in Brain and Eye of Poikilothermic Vertebrates. Biomolecules. 2015; 5(2):635-646. https://doi.org/10.3390/biom5020635
Chicago/Turabian StyleBaslow, Morris H., and David N. Guilfoyle. 2015. "N-acetyl-l-histidine, a Prominent Biomolecule in Brain and Eye of Poikilothermic Vertebrates" Biomolecules 5, no. 2: 635-646. https://doi.org/10.3390/biom5020635
APA StyleBaslow, M. H., & Guilfoyle, D. N. (2015). N-acetyl-l-histidine, a Prominent Biomolecule in Brain and Eye of Poikilothermic Vertebrates. Biomolecules, 5(2), 635-646. https://doi.org/10.3390/biom5020635