
A Neuroprotective Peptide Modulates Retinal cAMP Response Element-Binding Protein (CREB), Synapsin I (SYN1), and Growth-Associated Protein 43 (GAP43) in Rats with Silicone Oil-Induced Ocular Hypertension

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Peptide
2.3. Silicone Oil (SO) Model
2.4. Intravitreal Injections of CPP-P1 or Vehicle
2.5. SO Removal Protocol and Tissue Collection Timeline
2.6. IOP Measurements
2.7. Pattern Electroretinography (PERG) Measurements
2.8. Immunohistochemistry of Retina Cross-Sections
2.9. Immunohistochemistry of Whole Retinal Flat Mounts
2.10. Hematoxylin and Eosin Staining
2.11. Statistical Analysis
3. Results
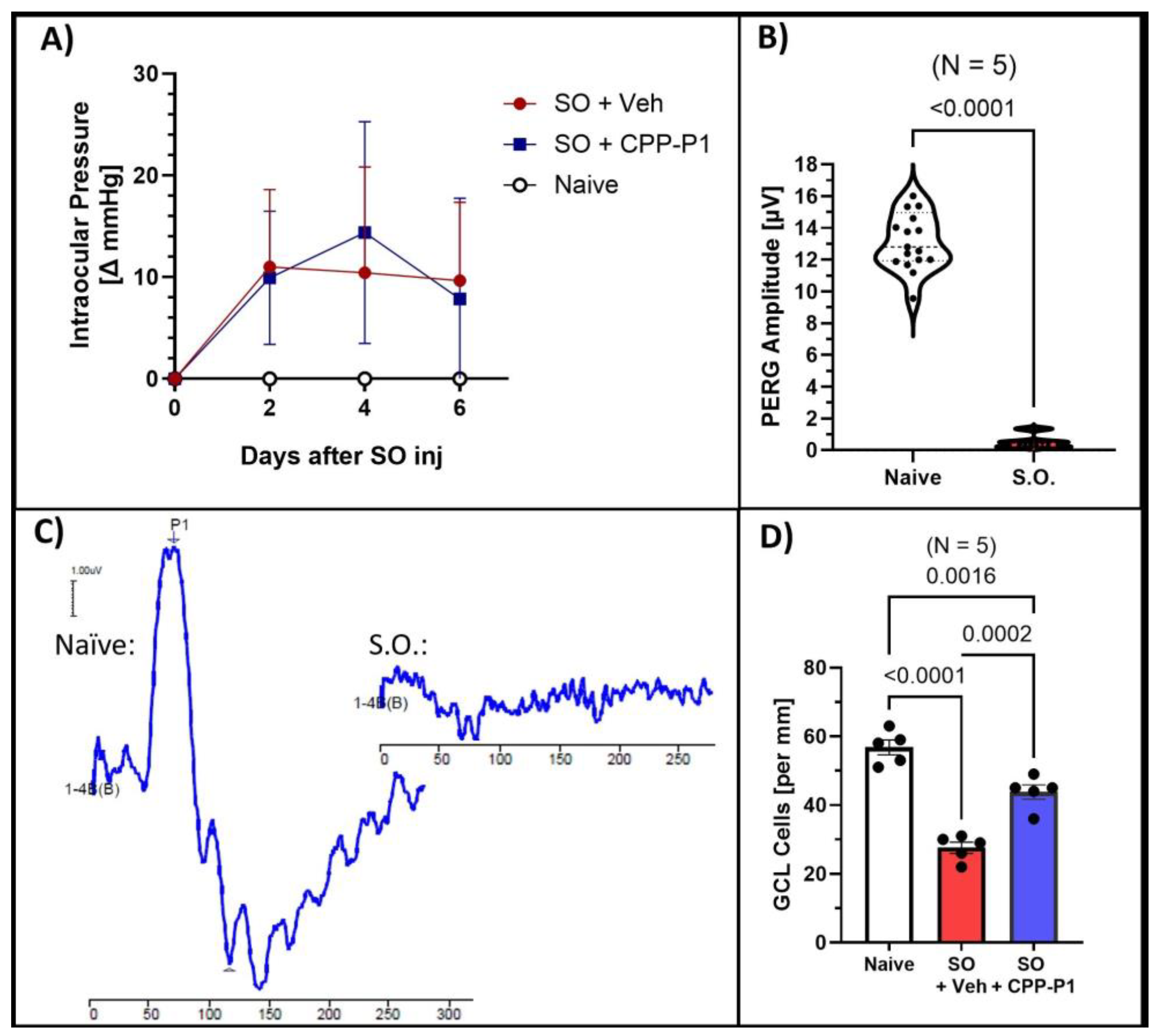
3.1. Intraocular Pressure (IOP)
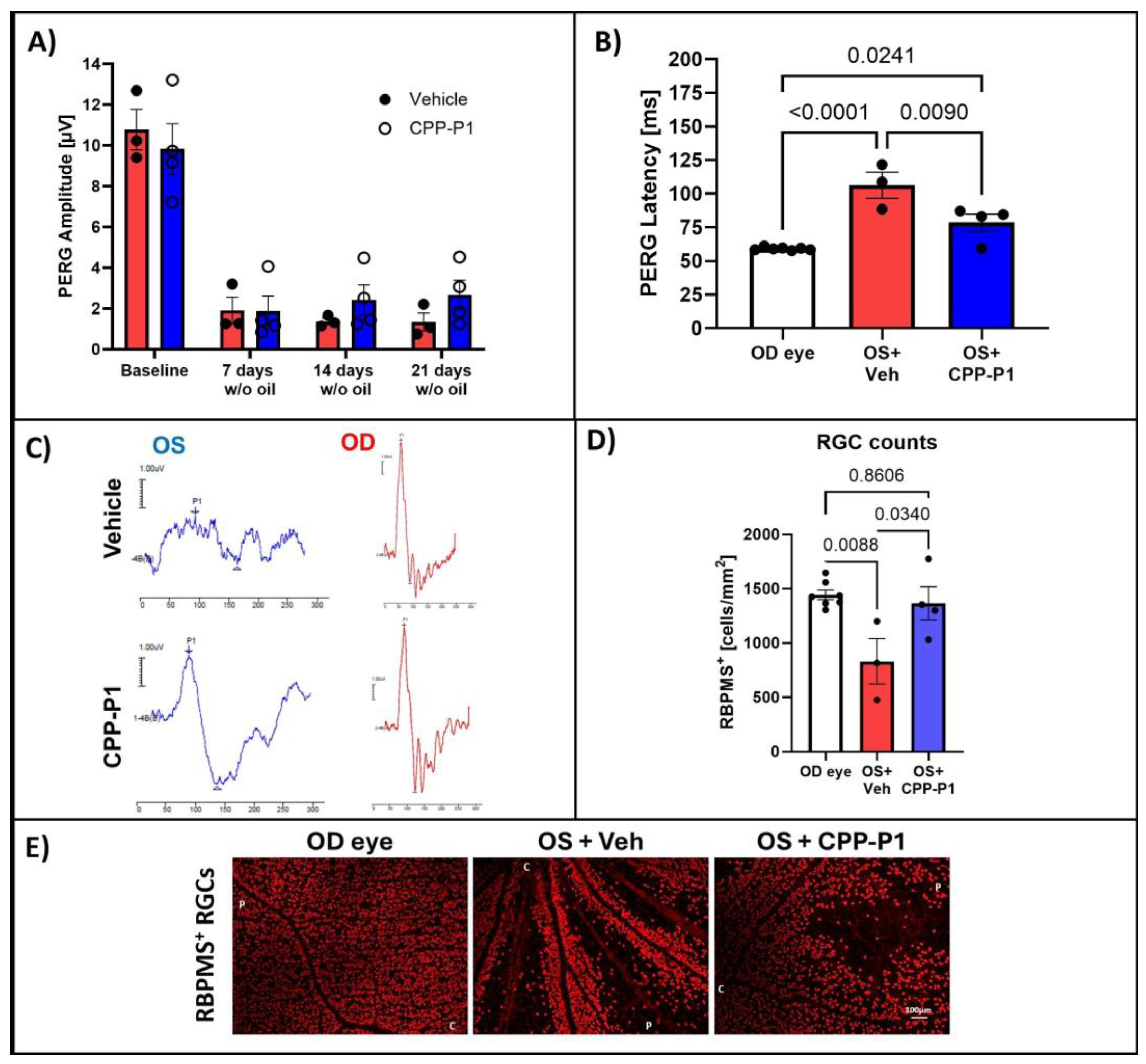
3.2. Pattern Electroretinography (PERG)
3.3. CPP-P1 Prevented the Loss of Cells in the GCL
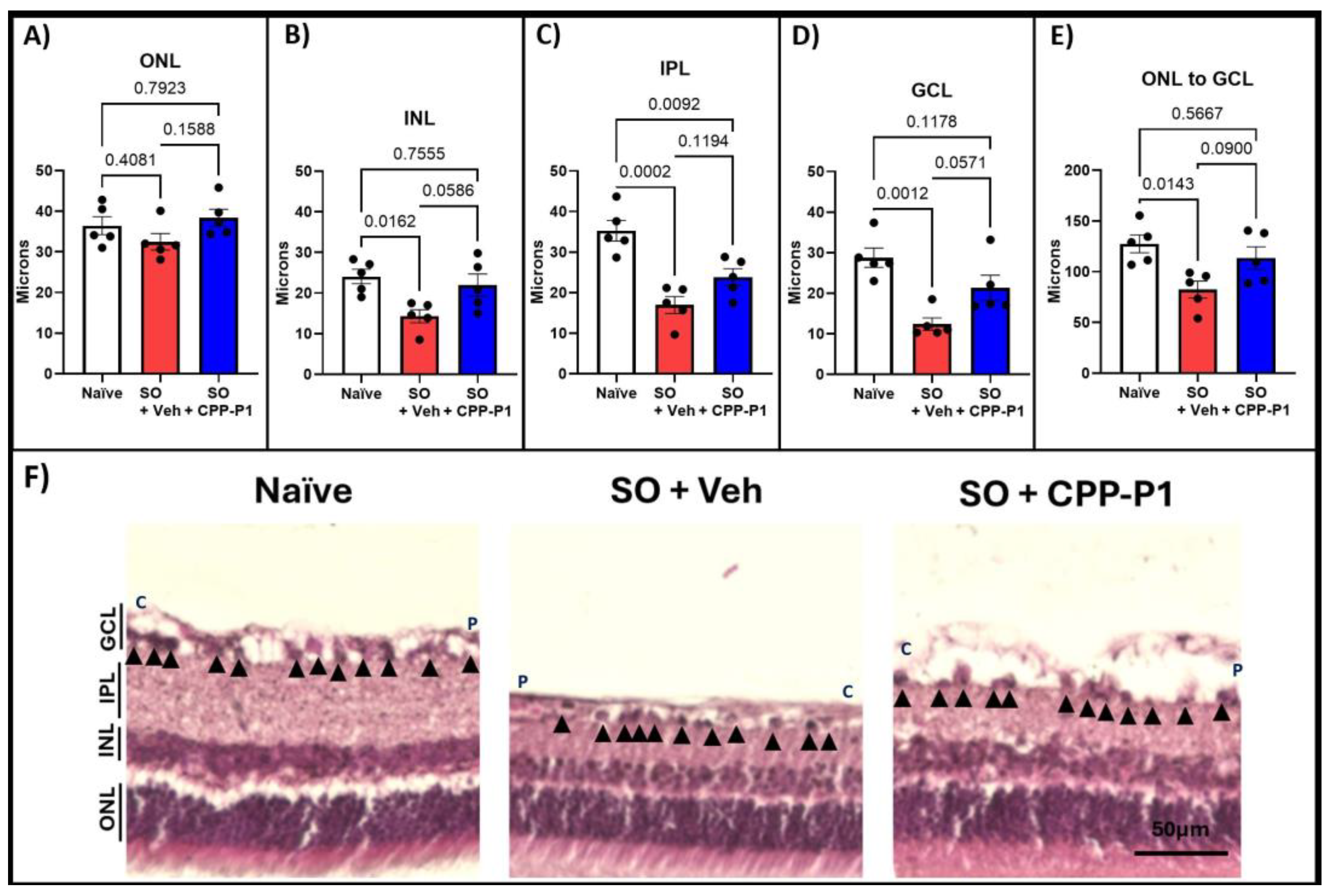
3.4. Retinal-Layer Thickness
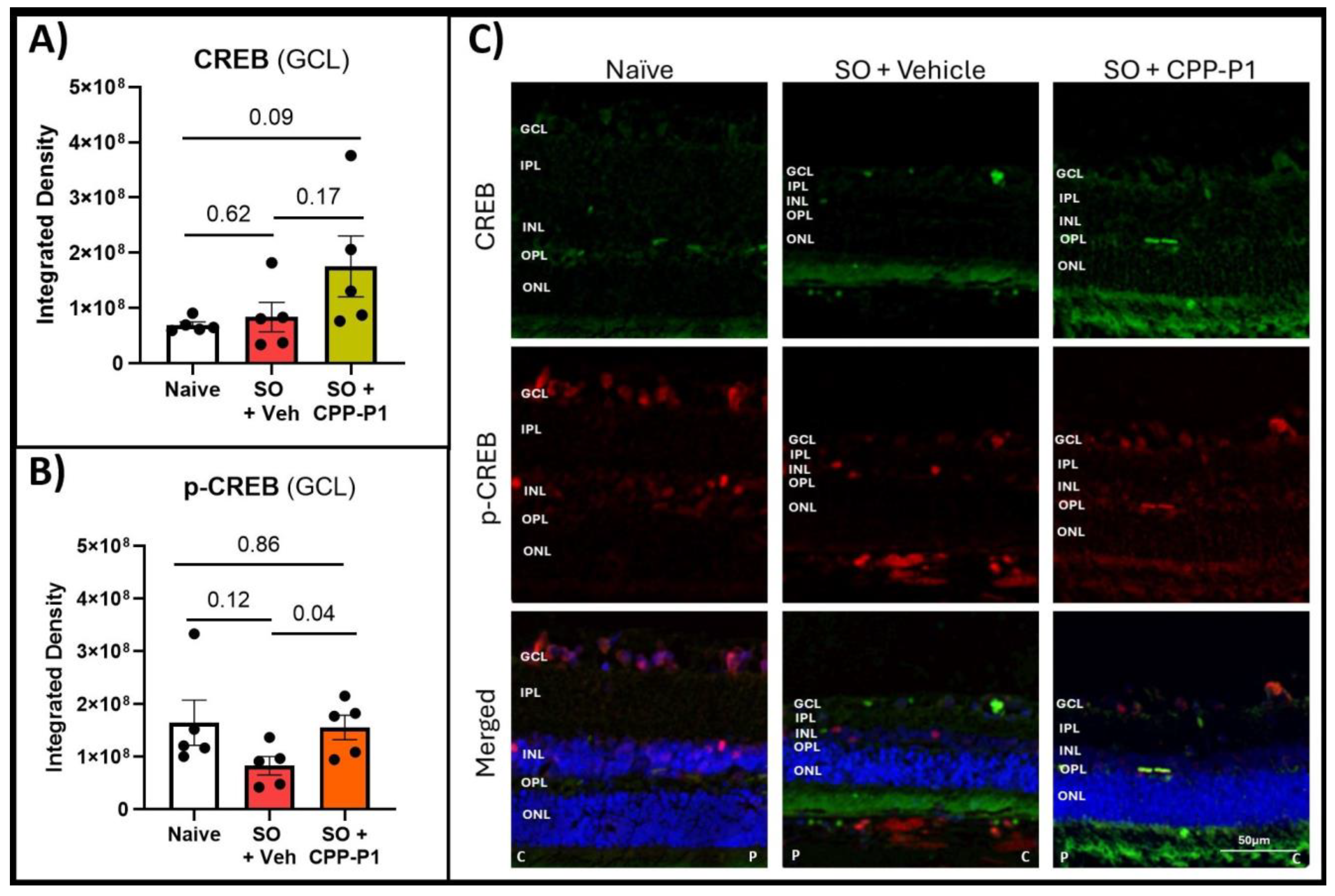
3.5. CREB and Phospho-CREB Analysis
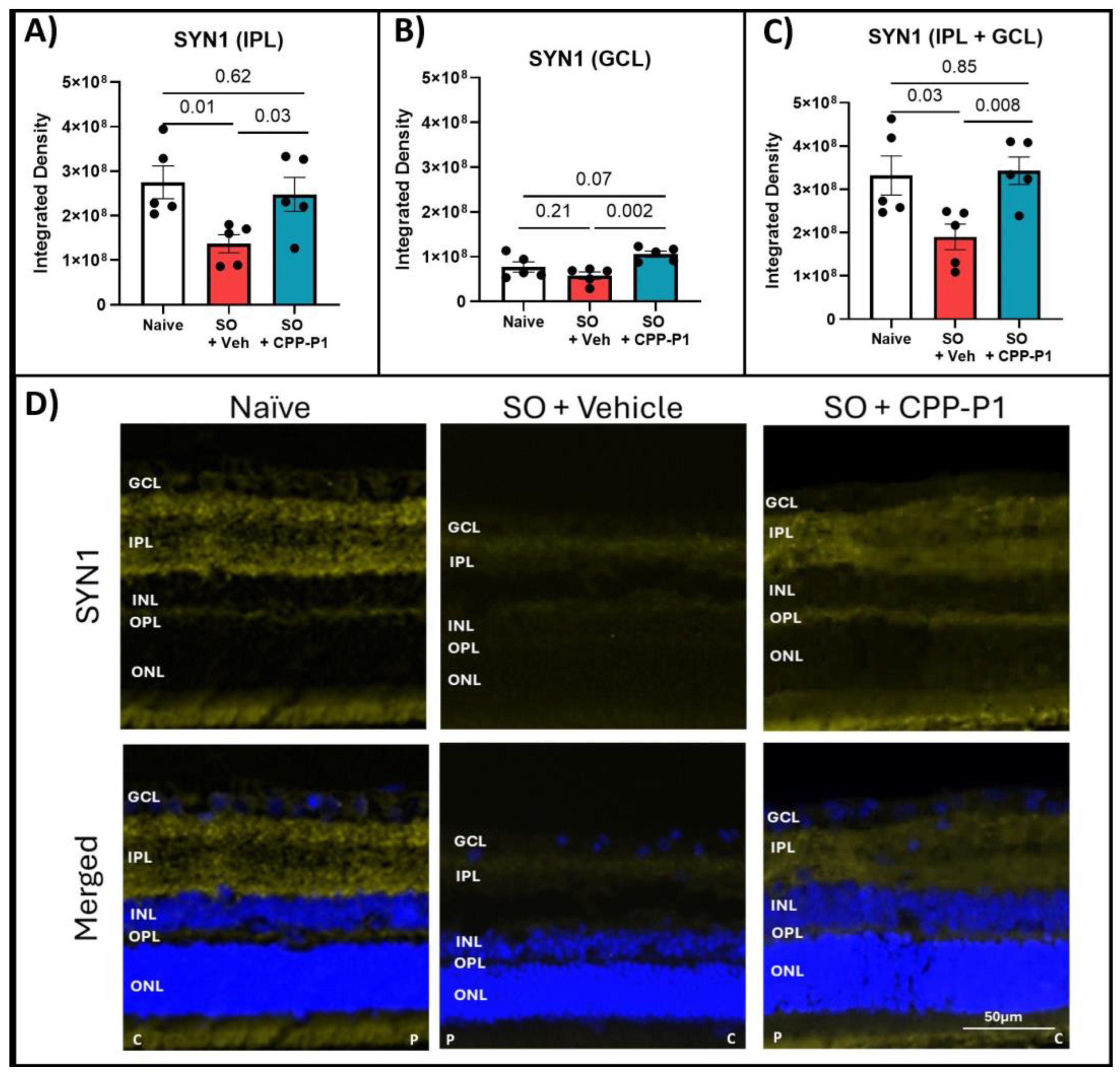
3.6. Expression of GAP43 and SYN1s Following IOP Elevation and Treatment with CPP-P1
3.7. Mitochondrial SOD2-Level Evaluation
3.8. Assessment of RGC Survival via RBPMS-Positive Cell Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allison, K.; Patel, D.; Alabi, O. Epidemiology of Glaucoma: The Past, Present, and Predictions for the Future. Cureus 2020, 12, e11686. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, C.G.; Liebmann, J.M.; Levin, L.A. Detection and measurement of clinically meaningful visual field progression in clinical trials for glaucoma. Prog. Retin. Eye Res. 2017, 56, 107–147. [Google Scholar] [CrossRef] [PubMed]
- Erb, C.; Eckert, S.; Gindorf, P.; Köhler, M.; Köhler, T.; Neuhann, L.; Ellrich, J. Electrical neurostimulation in glaucoma with progressive vision loss. Bioelectron. Med. 2022, 8, 6. [Google Scholar] [CrossRef]
- Quigley, H.A. Clinical trials for glaucoma neuroprotection are not impossible. Curr. Opin. Ophthalmol. 2012, 23, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Akerfelt, M.; Morimoto, R.I.; Sistonen, L. Heat shock factors: Integrators of cell stress, development and lifespan. Nat. Rev. Mol. Cell Biol. 2010, 11, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Andley, U.P. The lens epithelium: Focus on the expression and function of the alpha-crystallin chaperones. Int. J. Biochem. Cell Biol. 2008, 40, 317–323. [Google Scholar] [CrossRef]
- Fischer, D.; Hauk, T.G.; Müller, A.; Thanos, S. Crystallins of the beta/gamma-superfamily mimic the effects of lens injury and promote axon regeneration. Mol. Cell Neurosci. 2008, 37, 471–479. [Google Scholar] [CrossRef]
- Fort, P.E.; Lampi, K.J. New focus on alpha-crystallins in retinal neurodegenerative diseases. Exp. Eye Res. 2011, 92, 98–103. [Google Scholar] [CrossRef]
- Piri, N.; Kwong, J.M.; Caprioli, J. Crystallins in retinal ganglion cell survival and regeneration. Mol. Neurobiol. 2013, 48, 819–828. [Google Scholar] [CrossRef]
- Rajeswaren, V.; Wong, J.O.; Yabroudi, D.; Nahomi, R.B.; Rankenberg, J.; Nam, M.H.; Nagaraj, R.H. Small Heat Shock Proteins in Retinal Diseases. Front. Mol. Biosci. 2022, 9, 860375. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Chothe, P.; Sharma, K.K.; Baid, R.; Kompella, U.; Spee, C.; Kannan, N.; Manh, C.; Ryan, S.J.; Ganapathy, V.; et al. Antiapoptotic properties of α-crystallin-derived peptide chaperones and characterization of their uptake transporters in human RPE cells. Investig. Ophthalmol. Vis. Sci. 2013, 54, 2787–2798. [Google Scholar] [CrossRef] [PubMed]
- Vendredy, L.; Adriaenssens, E.; Timmerman, V. Small heat shock proteins in neurodegenerative diseases. Cell Stress. Chaperones 2020, 25, 679–699. [Google Scholar] [CrossRef] [PubMed]
- Anders, F.; Liu, A.; Mann, C.; Teister, J.; Lauzi, J.; Thanos, S.; Grus, F.H.; Pfeiffer, N.; Prokosch, V. The Small Heat Shock Protein α-Crystallin B Shows Neuroprotective Properties in a Glaucoma Animal Model. Int. J. Mol. Sci. 2017, 18, 2418. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.; Sreekumar, P.G.; Hinton, D.R. Novel roles for α-crystallins in retinal function and disease. Prog. Retin. Eye Res. 2012, 31, 576–604. [Google Scholar] [CrossRef]
- Kannan, R.; Sreekumar, P.G.; Kannan, N.; Ryan, S.J.; Kompella, U.B.; Sharma, K.; Hinton, D.R. Alpha Crystallin Derived Peptide Chaperone Protects Human RPE Cells From Oxidative Injury. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1441. [Google Scholar]
- Kourtis, N.; Nikoletopoulou, V.; Tavernarakis, N. Small heat-shock proteins protect from heat-stroke-associated neurodegeneration. Nature 2012, 490, 213–218. [Google Scholar] [CrossRef]
- Liu, H.; Bell, K.; Herrmann, A.; Arnhold, S.; Mercieca, K.; Anders, F.; Nagel-Wolfrum, K.; Thanos, S.; Prokosch, V. Crystallins Play a Crucial Role in Glaucoma and Promote Neuronal Cell Survival in an In Vitro Model Through Modulating Muller Cell Secretion. Invest. Ophthalmol. Vis. Sci. 2022, 63, 3. [Google Scholar] [CrossRef]
- Liu, J.P.; Schlosser, R.; Ma, W.Y.; Dong, Z.; Feng, H.; Lui, L.; Huang, X.Q.; Liu, Y.; Li, D.W. Human alphaA- and alphaB-crystallins prevent UVA-induced apoptosis through regulation of PKCalpha, RAF/MEK/ERK and AKT signaling pathways. Exp. Eye Res. 2004, 79, 393–403. [Google Scholar] [CrossRef]
- McGreal, R.S.; Kantorow, W.L.; Chauss, D.C.; Wei, J.; Brennan, L.A.; Kantorow, M. αB-crystallin/sHSP protects cytochrome c and mitochondrial function against oxidative stress in lens and retinal cells. Biochim. Biophys. Acta 2012, 1820, 921–930. [Google Scholar] [CrossRef]
- Munemasa, Y.; Kwong, J.M.; Caprioli, J.; Piri, N. The role of alphaA- and alphaB-crystallins in the survival of retinal ganglion cells after optic nerve axotomy. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3869–3875. [Google Scholar] [CrossRef]
- Nahomi, R.B.; Wang, B.; Raghavan, C.T.; Voss, O.; Doseff, A.I.; Santhoshkumar, P.; Nagaraj, R.H. Chaperone peptides of α-crystallin inhibit epithelial cell apoptosis, protein insolubilization, and opacification in experimental cataracts. J. Biol. Chem. 2013, 288, 13022–13035. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.-H.; Stankowska, D.L.; Johnson, G.A.; Nahomi, R.B.; Pantcheva, M.B.; Nagaraj, R.H. Peptains block retinal ganglion cell death in animal models of ocular hypertension: Implications for neuroprotection in glaucoma. Cell Death Dis. 2022, 13, 958. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.S.; Reddy, G.B. Emerging therapeutic roles of small heat shock protein-derived mini-chaperones and their delivery strategies. Biochimie 2022, 208, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, P.G.; Li, Z.; Wang, W.; Spee, C.; Hinton, D.R.; Kannan, R.; MacKay, J.A. Intra-vitreal αB crystallin fused to elastin-like polypeptide provides neuroprotection in a mouse model of age-related macular degeneration. J. Control. Release 2018, 283, 94–104. [Google Scholar] [CrossRef]
- Sreekumar, P.G.; Reddy, S.T.; Hinton, D.R.; Kannan, R. Mechanisms of RPE senescence and potential role of αB crystallin peptide as a senolytic agent in experimental AMD. Exp. Eye Res. 2022, 215, 108918. [Google Scholar] [CrossRef]
- Stankowska, D.L.; Nam, M.H.; Nahomi, R.B.; Chaphalkar, R.M.; Nandi, S.K.; Fudala, R.; Krishnamoorthy, R.R.; Nagaraj, R.H. Systemically administered peptain-1 inhibits retinal ganglion cell death in animal models: Implications for neuroprotection in glaucoma. Cell Death Discov. 2019, 5, 112. [Google Scholar] [CrossRef]
- Webster, J.M.; Darling, A.L.; Uversky, V.N.; Blair, L.J. Small Heat Shock Proteins, Big Impact on Protein Aggregation in Neurodegenerative Disease. Front. Pharmacol. 2019, 10, 1047. [Google Scholar] [CrossRef]
- Wu, N.; Yu, J.; Chen, S.; Xu, J.; Ying, X.; Ye, M.; Li, Y.; Wang, Y. α-Crystallin protects RGC survival and inhibits microglial activation after optic nerve crush. Life Sci. 2014, 94, 17–23. [Google Scholar] [CrossRef]
- Yan, H.; Peng, Y.; Huang, W.; Gong, L.; Li, L. The Protective Effects of αB-Crystallin on Ischemia-Reperfusion Injury in the Rat Retina. J. Ophthalmol. 2017, 2017, 7205408. [Google Scholar] [CrossRef]
- Johnson, G.A.; Kodati, B.; Nahomi, R.B.; Pham, J.H.; Krishnamoorthy, V.R.; Phillips, N.R.; Krishnamoorthy, R.R.; Nagaraj, R.H.; Stankowska, D.L. Mechanisms contributing to inhibition of retinal ganglion cell death by cell permeable peptain-1 under glaucomatous stress. Cell Death Discov. 2024, 10, 305. [Google Scholar] [CrossRef]
- Arthur, J.S.; Fong, A.L.; Dwyer, J.M.; Davare, M.; Reese, E.; Obrietan, K.; Impey, S. Mitogen- and stress-activated protein kinase 1 mediates cAMP response element-binding protein phosphorylation and activation by neurotrophins. J. Neurosci. 2004, 24, 4324–4332. [Google Scholar] [PubMed]
- Caracciolo, L.; Marosi, M.; Mazzitelli, J.; Latifi, S.; Sano, Y.; Galvan, L.; Kawaguchi, R.; Holley, S.; Levine, M.S.; Coppola, G.; et al. CREB controls cortical circuit plasticity and functional recovery after stroke. Nat. Commun. 2018, 9, 2250. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R. The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Mol. Brain 2012, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.R.; Dragunow, I. Is CREB a key to neuronal survival? Trends Neurosci. 2000, 23, 48–53. [Google Scholar] [CrossRef]
- Wang, L.; Nie, Q.; Gao, M.; Yang, L.; Xiang, J.W.; Xiao, Y.; Liu, F.Y.; Gong, X.D.; Fu, J.L.; Wang, Y.; et al. The transcription factor CREB acts as an important regulator mediating oxidative stress-induced apoptosis by suppressing αB-crystallin expression. Aging 2020, 12, 13594–13617. [Google Scholar] [CrossRef]
- Guo, X.J.; Tian, X.S.; Ruan, Z.; Chen, Y.T.; Wu, L.; Gong, Q.; Wang, W.; Zhang, H.Y. Dysregulation of neurotrophic and inflammatory systems accompanied by decreased CREB signaling in ischemic rat retina. Exp. Eye Res. 2014, 125, 156–163. [Google Scholar] [CrossRef]
- Guo, X.; Zhou, J.; Starr, C.; Mohns, E.J.; Li, Y.; Chen, E.P.; Yoon, Y.; Kellner, C.P.; Tanaka, K.; Wang, H.; et al. Preservation of vision after CaMKII-mediated protection of retinal ganglion cells. Cell 2021, 184, 4299–4314.e12. [Google Scholar] [CrossRef]
- Zhu, S.; Chen, L.; Wang, M.; Zhang, J.; Chen, G.; Yao, Y.; Song, S.; Li, T.; Zu, S.; Yu, Z.; et al. Schwann cell-derived extracellular vesicles as a potential therapy for retinal ganglion cell degeneration. J. Control Release 2023, 363, 641–656. [Google Scholar] [CrossRef]
- Eastwood, S.L.; Harrison, P.J. Synaptic pathology in the anterior cingulate cortex in schizophrenia and mood disorders. A review and a Western blot study of synaptophysin, GAP-43 and the complexins. Brain Res. Bull. 2001, 55, 569–578. [Google Scholar] [CrossRef]
- Jin, Y. Synaptogenesis. WormBook Online Rev. C. Elegans Biol. 2005, 16, 1–11. [Google Scholar] [CrossRef]
- Kaneda, M.; Nagashima, M.; Mawatari, K.; Nunome, T.; Muramoto, K.; Sugitani, K.; Kato, S. Growth-associated protein43 (GAP43) is a biochemical marker for the whole period of fish optic nerve regeneration. Adv. Exp. Med. Biol. 2010, 664, 97–104. [Google Scholar] [PubMed]
- Cho, K.S.; Yang, L.; Lu, B.; Feng Ma, H.; Huang, X.; Pekny, M.; Chen, D.F. Re-establishing the regenerative potential of central nervous system axons in postnatal mice. J. Cell Sci. 2005, 118 Pt 5, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Leon, S.; Yin, Y.; Nguyen, J.; Irwin, N.; Benowitz, L.I. Lens injury stimulates axon regeneration in the mature rat optic nerve. J. Neurosci. 2000, 20, 4615–4626. [Google Scholar] [CrossRef] [PubMed]
- John, A.; Ng-Cordell, E.; Hanna, N.; Brkic, D.; Baker, K. The neurodevelopmental spectrum of synaptic vesicle cycling disorders. J. Neurochem. 2021, 157, 208–228. [Google Scholar] [CrossRef]
- Parenti, I.; Leitao, E.; Kuechler, A.; Villard, L.; Goizet, C.; Courdier, C.; Bayat, A.; Rossi, A.; Julia, S.; Bruel, A.L.; et al. The different clinical facets of SYN1-related neurodevelopmental disorders. Front. Cell Dev. Biol. 2022, 10, 1019715. [Google Scholar] [CrossRef]
- Ferreira, S.M.; Lerner, S.F.; Brunzini, R.; Evelson, P.A.; Llesuy, S.F. Oxidative stress markers in aqueous humor of glaucoma patients. Am. J. Ophthalmol. 2004, 137, 62–69. [Google Scholar] [CrossRef]
- Ghanem, A.A.; Arafa, L.F.; El-Baz, A. Oxidative Stress Markers in Patients with Primary Open-Angle Glaucoma. Curr. Eye Res. 2010, 35, 295–301. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Huang, H.; Fang, F.; Webber, H.C.; Zhuang, P.; Liu, L.; Dalal, R.; Tang, P.H.; Mahajan, V.B.; et al. Silicone oil-induced ocular hypertension and glaucomatous neurodegeneration in mouse. Elife 2019, 8, e45881. [Google Scholar] [CrossRef]
- Fang, F.; Zhang, J.; Zhuang, P.; Liu, P.; Li, L.; Huang, H.; Webber, H.C.; Xu, Y.; Liu, L.; Dalal, R.; et al. Chronic mild and acute severe glaucomatous neurodegeneration derived from silicone oil-induced ocular hypertension. Sci. Rep. 2021, 11, 9052. [Google Scholar] [CrossRef]
- Zhang, J.; Fang, F.; Li, L.; Huang, H.; Webber, H.C.; Sun, Y.; Mahajan, V.B.; Hu, Y. A Reversible Silicon Oil-Induced Ocular Hypertension Model in Mice. J. Vis. Exp. 2019, 153, 10-3791. [Google Scholar] [CrossRef]
- Kodati, B.; McGrady, N.R.; Jefferies, H.B.; Stankowska, D.L.; Krishnamoorthy, R.R. Oral administration of a dual ET(A)/ET(B) receptor antagonist promotes neuroprotection in a rodent model of glaucoma. Mol. Vis. 2022, 28, 165–177. [Google Scholar] [PubMed]
- Dattilo, M.; Newman, N.J.; Biousse, V. Acute retinal arterial ischemia. Ann. Eye Sci. 2018, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Khakha, N.; Khan, H.; Kaur, A.; Singh, T.G. Therapeutic implications of phosphorylation-and dephosphorylation-dependent factors of cAMP-response element-binding protein (CREB) in neurodegeneration. Pharmacol. Rep. 2023, 75, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Fontanella, R.A.; Ghosh, P.; Pesapane, A.; Taktaz, F.; Puocci, A.; Franzese, M.; Feliciano, M.F.; Tortorella, G.; Scisciola, L.; Sommella, E.; et al. Tirzepatide prevents neurodegeneration through multiple molecular pathways. J. Transl. Med. 2024, 22, 114. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Wei, R.Y.; Tang, K.; Wang, Z.; Tan, N.H. Ginsenoside Rg1 promotes neurite growth of retinal ganglion cells through cAMP/PKA/CREB pathways. J. Ginseng Res. 2024, 48, 163–170. [Google Scholar] [CrossRef]
- Yan, D.; Dai, L.; Zhang, X.; Wang, Y.; Yan, H. Subchronic Acrylamide Exposure Activates PERK-eIF2α Signaling Pathway and Induces Synaptic Impairment in Rat Hippocampus. ACS Chem. Neurosci. 2022, 13, 1370–1381. [Google Scholar] [CrossRef]
- Naguib, S.; Backstrom, J.R.; Gil, M.; Calkins, D.J.; Rex, T.S. Retinal oxidative stress activates the NRF2/ARE pathway: An early endogenous protective response to ocular hypertension. Redox Biol. 2021, 42, 101883. [Google Scholar] [CrossRef]
- Jiang, W.; Tang, L.; Zeng, J.; Chen, B. Adeno-associated virus mediated SOD gene therapy protects the retinal ganglion cells from chronic intraocular pressure elevation induced injury via attenuating oxidative stress and improving mitochondrial dysfunction in a rat model. Am. J. Transl. Res. 2016, 8, 799–810. [Google Scholar]
- Chae, J.J.; Prausnitz, M.R.; Ethier, C.R. Effects of General Anesthesia on Intraocular Pressure in Rabbits. J. Am. Assoc. Lab. Anim. Sci. 2021, 60, 91–95. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Higashide, T.; Hatake, S.; Sugiyama, K. Effect of inhalation anesthesia with isoflurane on circadian rhythm of murine intraocular pressure. Exp. Eye Res. 2021, 203, 108420. [Google Scholar] [CrossRef]
- Lonze, B.E.; Ginty, D.D. Function and Regulation of CREB Family Transcription Factors in the Nervous System. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | % Loss in Vehicle | p-Value Vehicle: Naïve | % Loss in CPP-P1 | p-Value CPP-P1: Vehicle | p-Value CPP-P1: Naïve |
|---|---|---|---|---|---|
| ONL | 11% | 0.408 | 0% | 0.159 | 0.792 |
| INL | 41% | * 0.016 | 9% | 0.059 | 0.756 |
| IPL | 52% | *** 0.0002 | 33% | 0.119 | ** 0.009 |
| GCL | 57% | *** 0.001 | 26% | 0.057 | 0.118 |
| Total | 65% | * 0.014 | 11% | 0.090 | 0.567 |
| Naïve | p-Value Vehicle: Naïve | Vehicle | p-Value CPP-P1: Vehicle | CPP-P1 | p-Value CPP-P1: Naive | |
|---|---|---|---|---|---|---|
| CREB | 6.9E7 | 0.62 | 8.3E7 | 0.17 | 1.8E8 | 0.09 |
| p-CREB | 1.6E8 | 0.12 | 8.3E7 | * 0.04 | 1.6E8 | 0.86 |
| p-CREB/CREB | 2.3 | 1 | 0.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, G.A.; Krishnamoorthy, R.R.; Nagaraj, R.H.; Stankowska, D.L. A Neuroprotective Peptide Modulates Retinal cAMP Response Element-Binding Protein (CREB), Synapsin I (SYN1), and Growth-Associated Protein 43 (GAP43) in Rats with Silicone Oil-Induced Ocular Hypertension. Biomolecules 2025, 15, 219. https://doi.org/10.3390/biom15020219
Johnson GA, Krishnamoorthy RR, Nagaraj RH, Stankowska DL. A Neuroprotective Peptide Modulates Retinal cAMP Response Element-Binding Protein (CREB), Synapsin I (SYN1), and Growth-Associated Protein 43 (GAP43) in Rats with Silicone Oil-Induced Ocular Hypertension. Biomolecules. 2025; 15(2):219. https://doi.org/10.3390/biom15020219
Chicago/Turabian StyleJohnson, Gretchen A., Raghu R. Krishnamoorthy, Ram H. Nagaraj, and Dorota L. Stankowska. 2025. "A Neuroprotective Peptide Modulates Retinal cAMP Response Element-Binding Protein (CREB), Synapsin I (SYN1), and Growth-Associated Protein 43 (GAP43) in Rats with Silicone Oil-Induced Ocular Hypertension" Biomolecules 15, no. 2: 219. https://doi.org/10.3390/biom15020219
APA StyleJohnson, G. A., Krishnamoorthy, R. R., Nagaraj, R. H., & Stankowska, D. L. (2025). A Neuroprotective Peptide Modulates Retinal cAMP Response Element-Binding Protein (CREB), Synapsin I (SYN1), and Growth-Associated Protein 43 (GAP43) in Rats with Silicone Oil-Induced Ocular Hypertension. Biomolecules, 15(2), 219. https://doi.org/10.3390/biom15020219