
Network Pharmacology Analysis of the Potential Pharmacological Mechanism of a Sleep Cocktail


Abstract
1. Introduction
2. Materials and Methods
2.1. Candidate Therapeutic Targets of Sleep Cocktail
2.2. Gene Collection of Insomnia
2.3. Common Targets and Compound–Target Network Construction
2.4. Protein–Protein Interaction (PPI), Network Construction, and Network Analysis
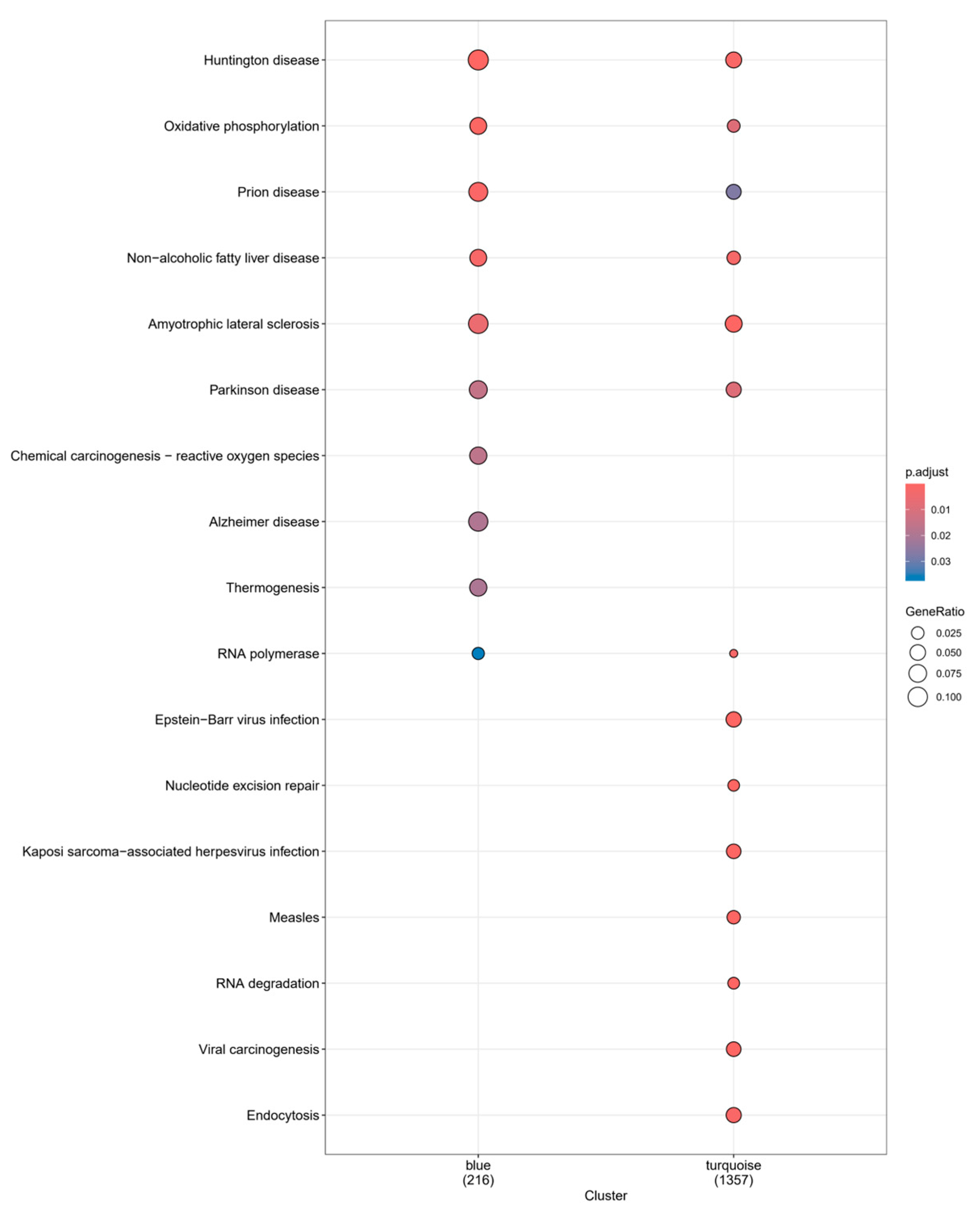
2.5. GO and KEGG Enrichment Analysis and Compound–Target-Pathway Network Construction
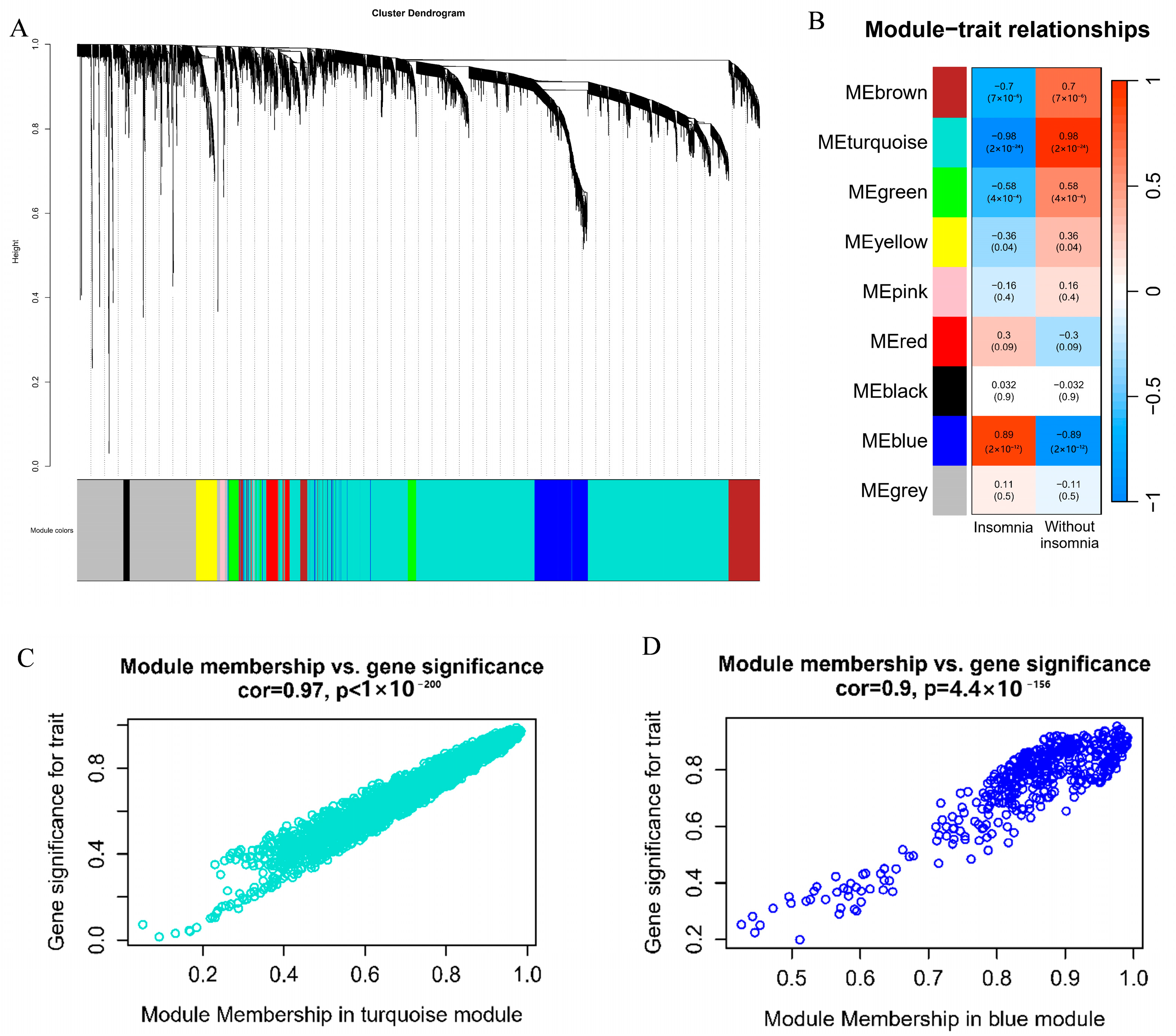
2.6. Weighted Correlation Network Analysis
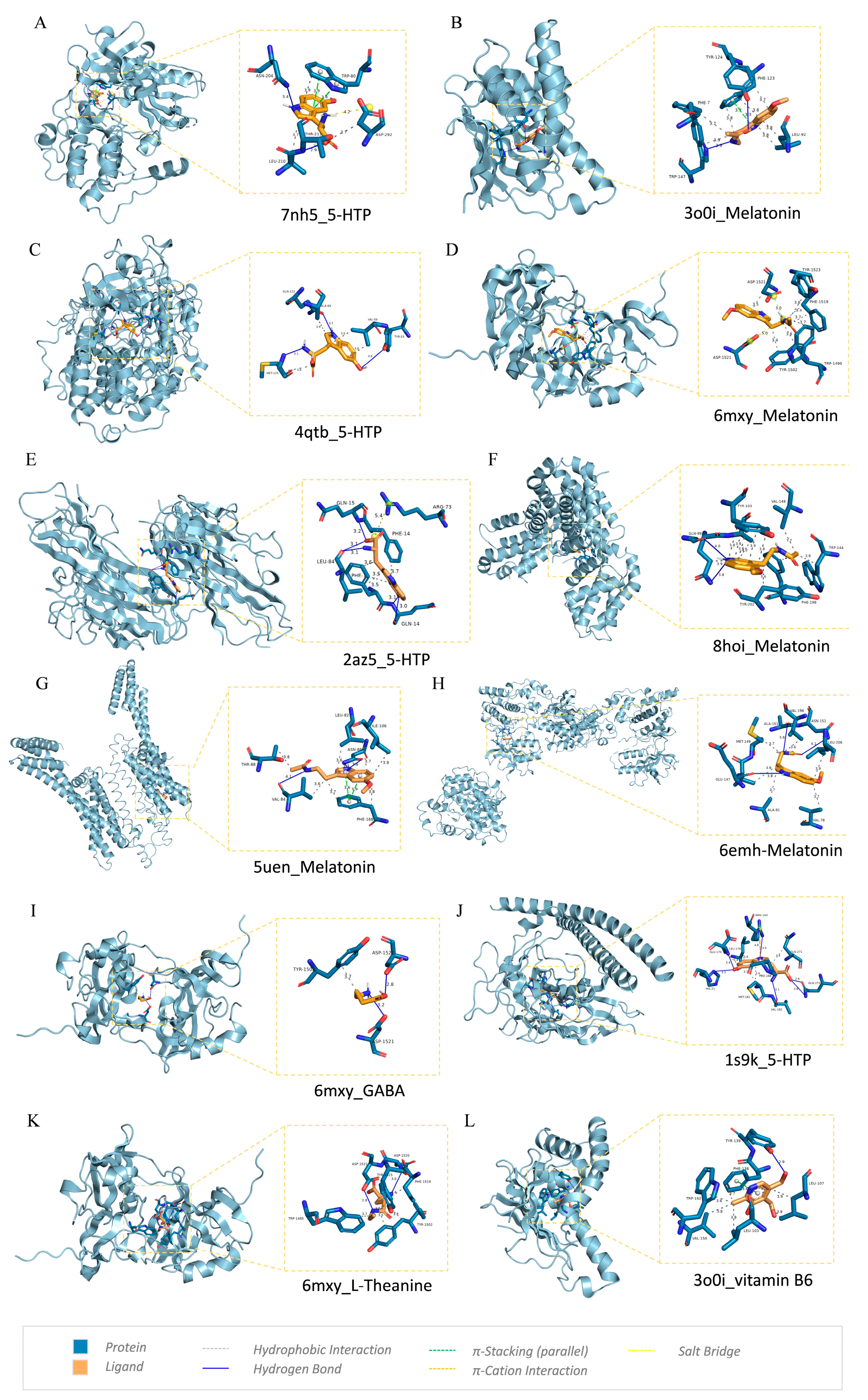
2.7. Molecular Docking
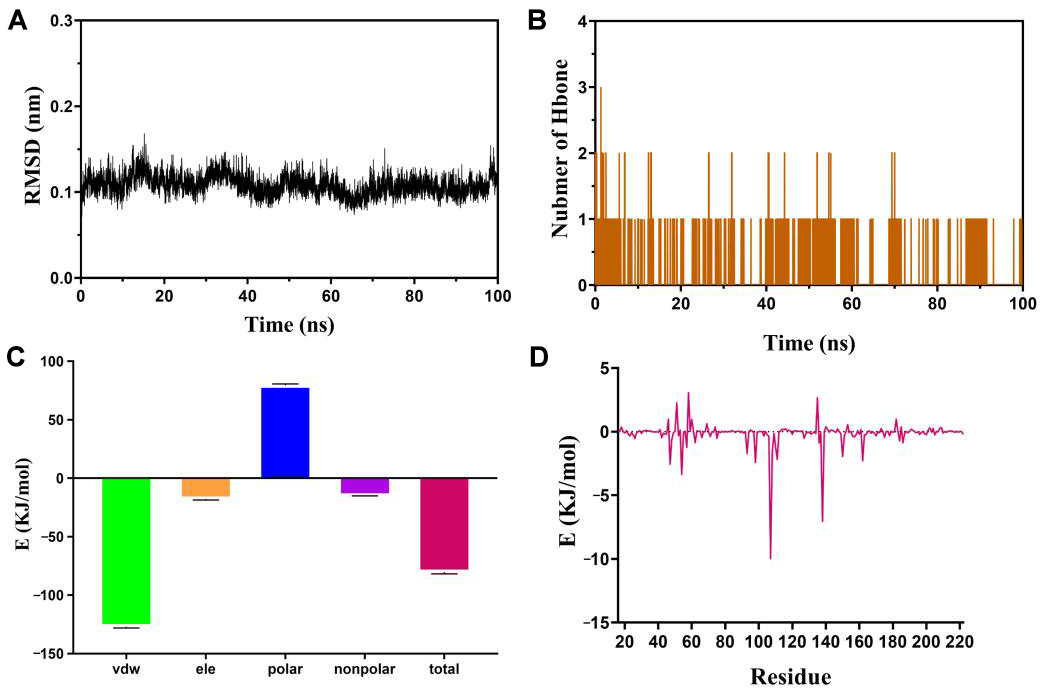
2.8. Molecular Dynamics Simulation
3. Results
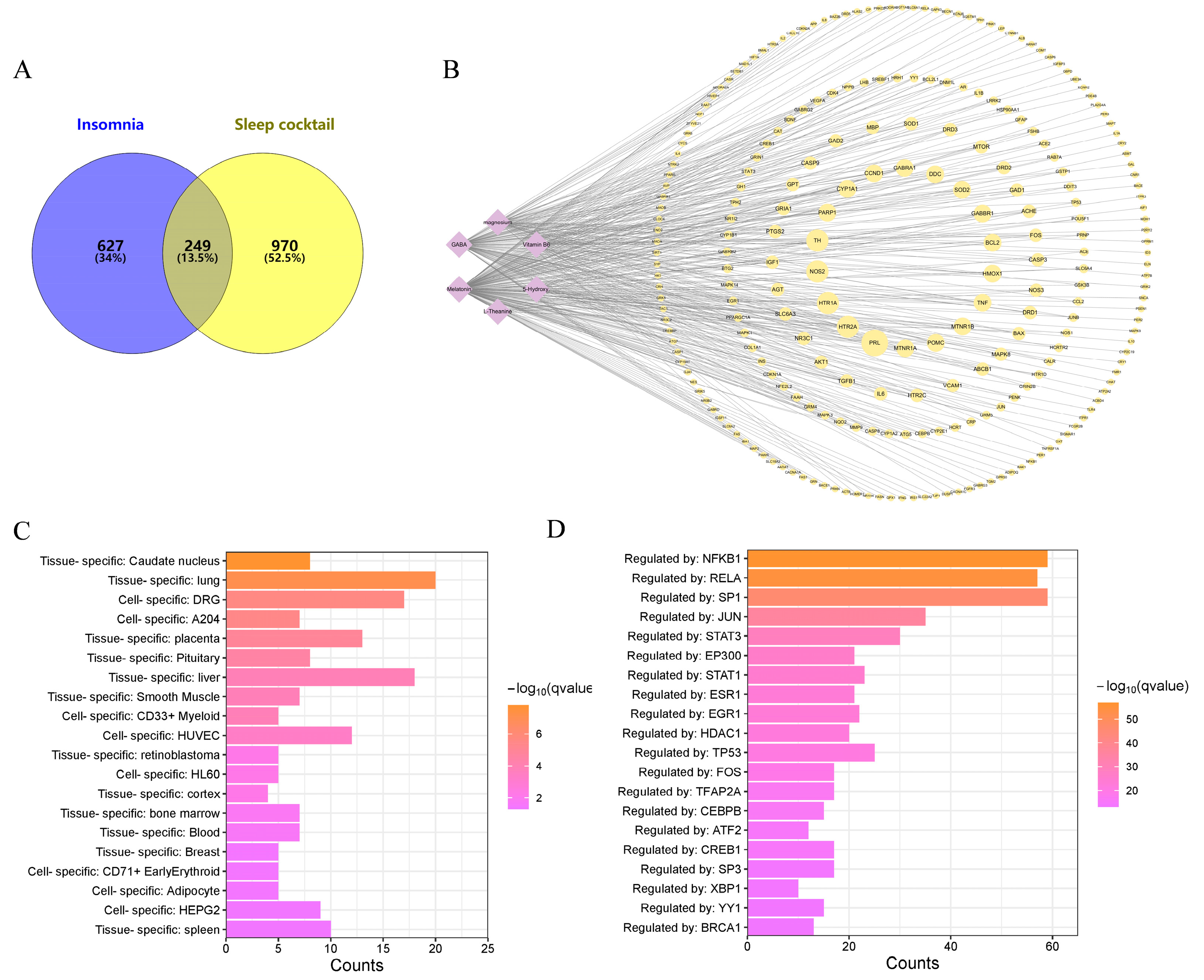
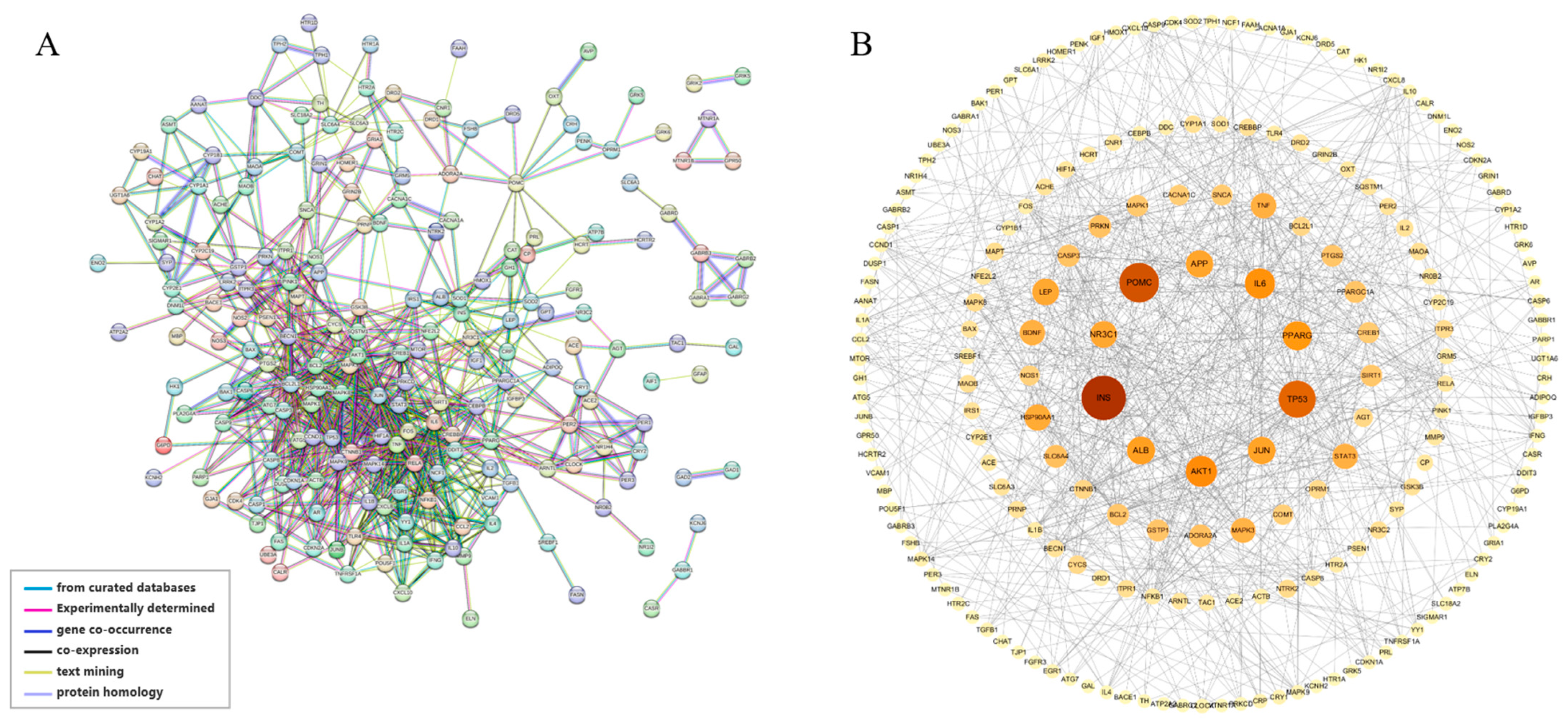
3.1. Common Targets between Sleep Cocktail and Insomnia and PPI Network Construction
3.2. GO, KEGG Pathway Enrichment Analysis, and Compound–Target Pathway Network
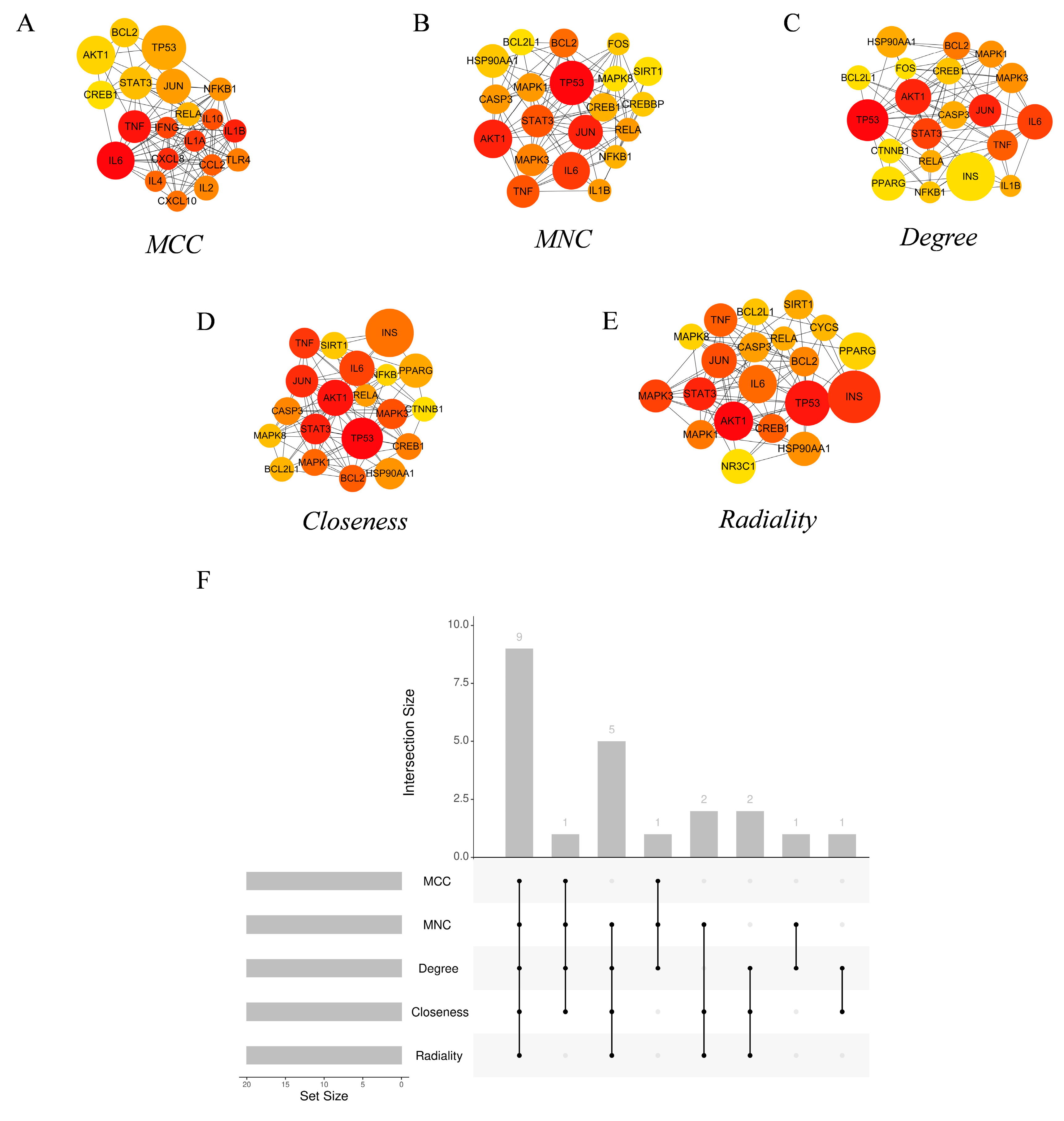
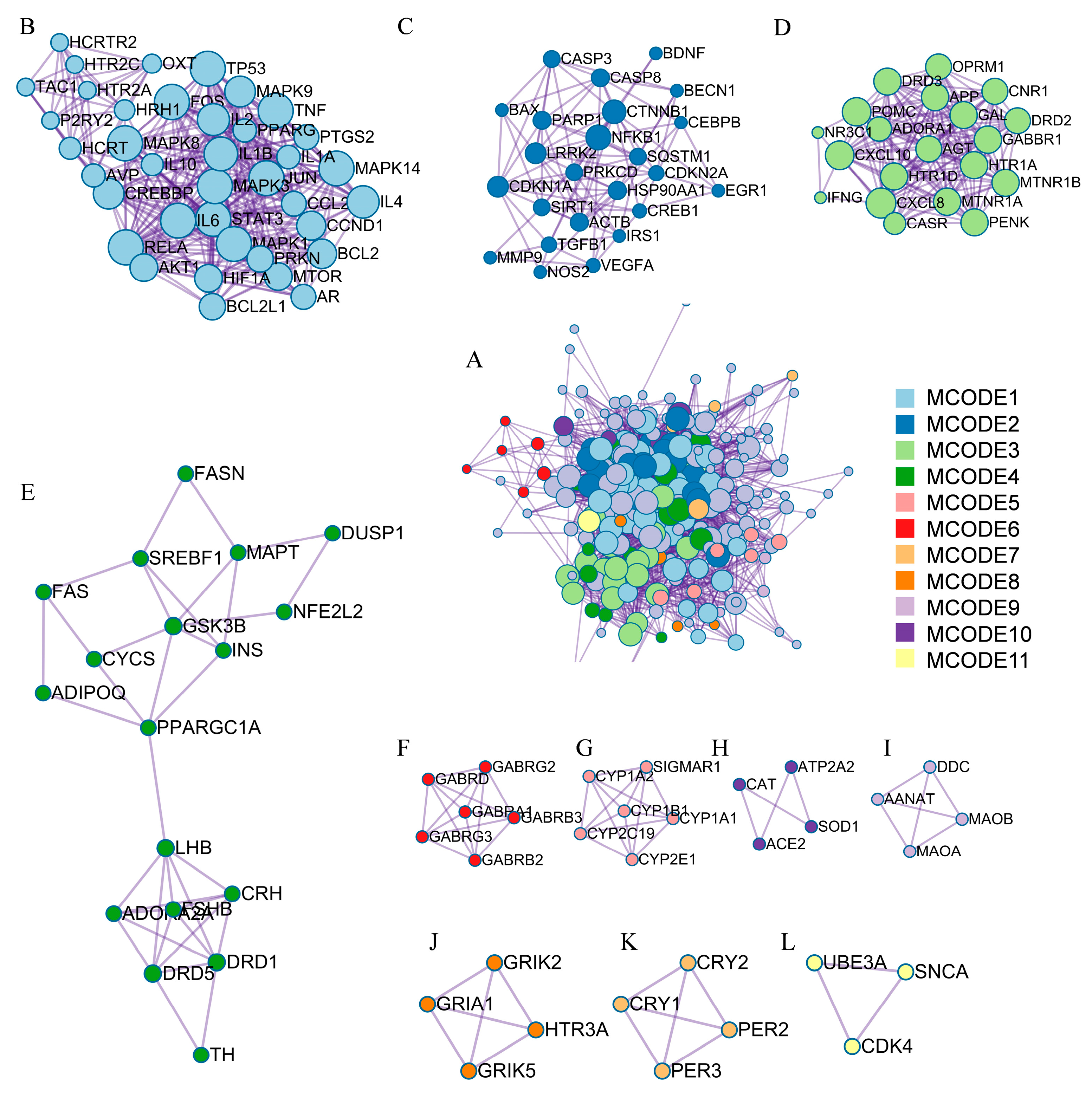
3.3. PPI Network Hub Genes and Functional Cluster Analysis
3.4. Weighted Correlation Network Analysis and Functional Analysis of PBMC Transcriptome
3.5. Molecular Docking Verification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roth, T. Insomnia: Definition, Prevalence, Etiology, and Consequences. J. Clin. Sleep Med. 2007, 3 (Suppl. 5), S7–S10. [Google Scholar] [CrossRef] [PubMed]
- Gardani, M.; Bradford, D.R.R.; Russell, K.; Allan, S.; Beattie, L.; Ellis, J.G.; Akram, U. A Systematic Review and Meta-Analysis of Poor Sleep, Insomnia Symptoms and Stress in Undergraduate Students. Sleep Med. Rev. 2022, 61, 101565. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.M.; Jarrin, D.C. Epidemiology of Insomnia. Sleep Med. Clin. 2022, 17, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Sutton, E.L. Insomnia. Ann. Intern. Med. 2021, 174, ITC33–ITC48. [Google Scholar] [CrossRef] [PubMed]
- DiBonaventura, M.; Richard, L.; Kumar, M.; Forsythe, A.; Flores, N.M.; Moline, M. The Association between Insomnia and Insomnia Treatment Side Effects on Health Status, Work Productivity, and Healthcare Resource Use. PLoS ONE 2015, 10, e0137117. [Google Scholar] [CrossRef] [PubMed]
- Atkin, T.; Comai, S.; Gobbi, G. Drugs for Insomnia beyond Benzodiazepines: Pharmacology, Clinical Applications, and Discovery. Pharmacol. Rev. 2018, 70, 197–245. [Google Scholar] [CrossRef] [PubMed]
- Birdsall, T.C. 5-Hydroxytryptophan: A Clinically-Effective Serotonin Precursor. Altern. Med. Rev. 1998, 3, 271–280. [Google Scholar]
- Rahman, M.K.; Nagatsu, T.; Sakurai, T.; Hori, S.; Abe, M.; Matsuda, M. Effect of Pyridoxal Phosphate Deficiency on Aromatic L-Amino Acid Decarboxylase Activity With L-Dopa and L-5-Hydroxytryptophan as Substrates in Rats. Jpn. J. Pharmacol. 1982, 32, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Kennaway, D.J. A Critical Review of Melatonin Assays: Past and Present. J. Pineal Res. 2019, 67, e12572. [Google Scholar] [CrossRef]
- Kakuda, T. Neuroprotective Effects of Theanine and Its Preventive Effects on Cognitive Dysfunction. Pharmacol. Res. 2011, 64, 162–168. [Google Scholar] [CrossRef]
- Ngo, D.H.; Vo, T.S. An Updated Review on Pharmaceutical Properties of Gamma-Aminobutyric Acid. Molecules 2019, 24, 2678. [Google Scholar] [CrossRef] [PubMed]
- Liwinski, T.; Lang, U.E.; Brühl, A.B.; Schneider, E. Exploring the Therapeutic Potential of Gamma-Aminobutyric Acid in Stress and Depressive Disorders through the Gut–Brain Axis. Biomedicines 2023, 11, 3128. [Google Scholar] [CrossRef] [PubMed]
- Mills, D.J. The Aging GABAergic System and Its Nutritional Support. J. Nutr. Metab. 2021, 2021, 6655064. [Google Scholar] [CrossRef]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of Oral Gamma-Aminobutyric Acid (GABA) Administration on Stress and Sleep in Humans: A Systematic Review. Front. Neurosci. 2020, 14, 559962. [Google Scholar] [CrossRef] [PubMed]
- Erdman, J.W.; MacDonald, I.A.; Zeisel, S.H. Present Knowledge in Nutrition, 10th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Combs, G.F.; McClung, J.P. The Vitamins: Fundamental Aspects in Nutrition and Health, 6th ed.Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Abbasi, B.; Kimiagar, M.; Sadeghniiat, K.; Shirazi, M.M.; Hedayati, M.; Rashidkhani, B. The Effect of Magnesium Supplementation on Primary Insomnia in Elderly: A Double-Blind Placebo-Controlled Clinical Trial. J. Res. Med. Sci. 2012, 17, 1161. [Google Scholar]
- Cao, Y.; Zhen, S.; Taylor, A.W.; Appleton, S.; Atlantis, E.; Shi, Z. Magnesium Intake and Sleep Disorder Symptoms: Findings from the Jiangsu Nutrition Study of Chinese Adults at Five-Year Follow-Up. Nutrients 2018, 10, 1354. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jo, K.; Hong, K.B.; Han, S.H.; Suh, H.J. GABA and L-Theanine Mixture Decreases Sleep Latency and Improves NREM Sleep. Pharm. Biol. 2019, 57, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.B.; Park, Y.; Suh, H.J. Sleep-Promoting Effects of the GABA/5-HTP Mixture in Vertebrate Models. Behav. Brain Res. 2016, 310, 36–41. [Google Scholar] [CrossRef]
- Hong, K.B.; Park, Y.; Suh, H.J. Two Combined Amino Acids Promote Sleep Activity in Caffeine-Induced Sleepless Model Systems. Nutr. Res. Pract. 2018, 12, 208. [Google Scholar] [CrossRef]
- Hong, K.B.; Park, Y.; Suh, H.J. Sleep-Promoting Effects of a GABA/5-HTP Mixture: Behavioral Changes and Neuromodulation in an Invertebrate Model. Life Sci. 2016, 150, 42–49. [Google Scholar] [CrossRef]
- Djokic, G.; Vojvodić, P.; Korcok, D.; Agic, A.; Rankovic, A.; Djordjevic, V.; Vojvodic, A.; Vlaskovic-Jovicevic, T.; Peric-Hajzler, Z.; Matovic, D.; et al. The Effects of Magnesium—Melatonin-Vit B Complex Supplementation in Treatment of Insomnia. Open Access Maced. J. Med. Sci. 2019, 7, 3101. [Google Scholar] [CrossRef] [PubMed]
- Bravaccio, C.; Terrone, G.; Rizzo, R.; Gulisano, M.; Tosi, M.; Curatolo, P.; Gialloreti, L.E. Use of Nutritional Supplements Based on Melatonin, Tryptophan and Vitamin B6 (Melamil Tripto®) in Children with Primary Chronic Headache, with or without Sleep Disorders: A Pilot Study. Minerva Pediatr. 2020, 72, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Sutanto, C.N.; Xia, X.; Heng, C.W.; Tan, Y.S.; Lee, D.P.S.; Fam, J.; Kim, J.E. The Impact of 5-Hydroxytryptophan Supplementation on Sleep Quality and Gut Microbiota Composition in Older Adults: A Randomized Controlled Trial. Clin. Nutr. 2024, 43, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Bhandage, A.K.; Jin, Z.; Korol, S.V.; Shen, Q.; Pei, Y.; Deng, Q.; Espes, D.; Carlsson, P.O.; Kamali-Moghaddam, M.; Birnir, B. GABA Regulates Release of Inflammatory Cytokines From Peripheral Blood Mononuclear Cells and CD4+ T Cells and Is Immunosuppressive in Type 1 Diabetes. EBioMedicine 2018, 30, 283–294. [Google Scholar] [CrossRef]
- Tian, J.; Yong, J.; Dang, H.; Kaufman, D.L. Oral GABA Treatment Downregulates Inflammatory Responses in a Mouse Model of Rheumatoid Arthritis. Autoimmunity 2011, 44, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Lu, Y.; Zhang, H.; Chau, C.H.; Dang, H.N.; Kaufman, D.L. γ-Aminobutyric Acid Inhibits T Cell Autoimmunity and the Development of Inflammatory Responses in a Mouse Type 1 Diabetes Model. J. Immunol. 2004, 173, 5298–5304. [Google Scholar] [CrossRef]
- Li, S.; Hu, Y.J. Network Pharmacology: An Approach to the Analysis of Complex Systems Underlying Traditional Chinese Medicine. World J. Tradit. Chin. Med. 2018, 4, 135–136. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New Data Content and Improved Web Interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated Data and New Features for Efficient Prediction of Protein Targets of Small Molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Santos, A.; Von Mering, C.; Jensen, L.J.; Bork, P.; Kuhn, M. STITCH 5: Augmenting Protein-Chemical Interaction Networks with Tissue and Affinity Data. Nucleic Acids Res. 2016, 44, D380–D384. [Google Scholar] [CrossRef]
- Davis, A.P.; Wiegers, T.C.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2023. Nucleic Acids Res 2023, 51, D1257–D1262. [Google Scholar] [CrossRef]
- Schriml, L.M.; Arze, C.; Nadendla, S.; Chang, Y.W.W.; Mazaitis, M.; Felix, V.; Feng, G.; Kibbe, W.A. Disease Ontology: A Backbone for Disease Semantic Integration. Nucleic Acids Res. 2012, 40, D940–D946. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Yan, G.R.; He, Q.Y. DOSE: An R/Bioconductor Package for Disease Ontology Semantic and Enrichment Analysis. Bioinformatics 2015, 31, 608–609. [Google Scholar] [CrossRef] [PubMed]
- Amberger, J.S.; Bocchini, C.A.; Scott, A.F.; Hamosh, A. OMIM.Org: Leveraging Knowledge across Phenotype-Gene Relationships. Nucleic Acids Res. 2019, 47, D1038–D1043. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Iny Stein, T.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Martin, M.J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The Universal Protein Knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams.—References—Scientific Research Publishing. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 23 February 2024).
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING Database in 2023: Protein-Protein Association Networks and Functional Enrichment Analyses for Any Sequenced Genome of Interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Shim, H.; Shin, D.; Shim, J.E.; Ko, Y.; Shin, J.; Kim, H.; Cho, A.; Kim, E.; Lee, T.; et al. TRRUST: A Reference Database of Human Transcriptional Regulatory Interactions. Sci. Rep. 2015, 5, 11432. [Google Scholar] [CrossRef]
- Pan, J.B.; Hu, S.C.; Shi, D.; Cai, M.C.; Li, Y.B.; Zou, Q.; Ji, Z.L. PaGenBase: A Pattern Gene Database for the Global and Dynamic Understanding of Gene Function. PLoS ONE 2013, 8, e80747. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. CytoHubba: Identifying Hub Objects and Sub-Networks from Complex Interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Bader, G.D.; Hogue, C.W.V. An Automated Method for Finding Molecular Complexes in Large Protein Interaction Networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for Integration and Interpretation of Large-Scale Molecular Data Sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [PubMed]
- Sean, D.; Meltzer, P.S. GEOquery: A Bridge between the Gene Expression Omnibus (GEO) and BioConductor. Bioinformatics 2007, 23, 1846–1847. [Google Scholar] [CrossRef] [PubMed]
- Piber, D.; Cho, J.H.; Lee, O.; Lamkin, D.M.; Olmstead, R.; Irwin, M.R. Sleep Disturbance and Activation of Cellular and Transcriptional Mechanisms of Inflammation in Older Adults. Brain Behav. Immun. 2022, 106, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. Fast R Functions for Robust Correlations and Hierarchical Clustering. J. Stat. Softw. 2012, 46, i11. [Google Scholar] [CrossRef]
- Young, J.Y.; Westbrook, J.D.; Feng, Z.; Peisach, E.; Persikova, I.; Sala, R.; Sen, S.; Berrisford, J.M.; Swaminathan, G.J.; Oldfield, T.J.; et al. Worldwide Protein Data Bank Biocuration Supporting Open Access to High-Quality 3D Structural Biology Data. Database 2018, 2018, bay002. [Google Scholar] [CrossRef]
- Lill, M.A.; Danielson, M.L. Computer-Aided Drug Design Platform Using PyMOL. J. Comput. Aided Mol. Des. 2011, 25, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Feinstein, W.P.; Brylinski, M. Calculating an Optimal Box Size for Ligand Docking and Virtual Screening against Experimental and Predicted Binding Pockets. J. Cheminform. 2015, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand-Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the Scope of the Protein-Ligand Interaction Profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, Flexible, and Free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Hollingsworth, S.A.; Dror, R.O. Molecular Dynamics Simulation for All. Neuron 2018, 99, 1129–1143. [Google Scholar] [CrossRef]
- Cho, H.J.; Seeman, T.E.; Kiefe, C.I.; Lauderdale, D.S.; Irwin, M.R. Sleep Disturbance and Longitudinal Risk of Inflammation: Moderating Influences of Social Integration and Social Isolation in the Coronary Artery Risk Development in Young Adults (CARDIA) Study. Brain Behav. Immun. 2015, 46, 319–326. [Google Scholar] [CrossRef]
- Smagula, S.F.; Stone, K.L.; Redline, S.; Ancoli-Israel, S.; Barrett-Connor, E.; Lane, N.E.; Orwoll, E.S.; Cauley, J.A. Actigraphy- and Polysomnography-Measured Sleep Disturbances, Inflammation, and Mortality among Older Men. Psychosom. Med. 2016, 78, 686–696. [Google Scholar] [CrossRef]
- Carroll, J.E.; Carrillo, C.; Olmstead, R.; Witarama, T.; Breen, E.C.; Yokomizo, M.; Seeman, T.E.; Irwin, M.R. Sleep Deprivation and Divergent Toll-like Receptor-4 Activation of Cellular Inflammation in Aging. Sleep 2015, 38, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.R.; Carrillo, C.; Olmstead, R. Sleep Loss Activates Cellular Markers of Inflammation: Sex Differences. Brain Behav. Immun. 2010, 24, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.R.; Wang, M.; Campomayor, C.O.; Collado-Hidalgo, A.; Cole, S. Sleep Deprivation and Activation of Morning Levels of Cellular and Genomic Markers of Inflammation. Arch. Intern. Med. 2006, 166, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.R.; Wang, M.; Ribeiro, D.; Cho, H.J.; Olmstead, R.; Breen, E.C.; Martinez-Maza, O.; Cole, S. Sleep Loss Activates Cellular Inflammatory Signaling. Biol. Psychiatry 2008, 64, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Fujita, N.; Tsuruo, T. Modulation of Akt Kinase Activity by Binding to Hsp90. Proc. Natl. Acad. Sci. USA 2000, 97, 10832–10837. [Google Scholar] [CrossRef] [PubMed]
- Bohonowych, J.E.; Hance, M.W.; Nolan, K.D.; Defee, M.; Parsons, C.H.; Isaacs, J.S. Extracellular Hsp90 Mediates an NF-ΚB Dependent Inflammatory Stromal Program: Implications for the Prostatetumor Microenvironment. Prostate 2014, 74, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Ammirante, M.; Rosati, A.; Gentilella, A.; Festa, M.; Petrella, A.; Marzullo, L.; Pascale, M.; Belisario, M.A.; Leone, A.; Turco, M.C. The Activity of Hsp90 Alpha Promoter Is Regulated by NF-Kappa B Transcription Factors. Oncogene 2008, 27, 1175–1178. [Google Scholar] [CrossRef]
- Bieging, K.T.; Mello, S.S.; Attardi, L.D. Unravelling Mechanisms of P53-Mediated Tumour Suppression. Nat. Rev. Cancer 2014, 14, 359–370. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | UniProt ID | Gene Symbol | Protein Name |
|---|---|---|---|
| 1 | P31749 | AKT1 | RAC-alpha serine/threonine-protein kinase, 2.7.11.1, protein kinase B, PKB, protein kinase B alpha, PKB alpha, proto-oncogene c-Akt, RAC-PK-alpha |
| 2 | P10415 | BCL2 | apoptosis regulator Bcl-2 |
| 3 | P16220 | CREB1 | Cyclic AMP-responsive element-binding protein 1, CREB-1, cAMP-responsive element-binding protein 1 |
| 4 | P05231 | IL6 | Interleukin-6, IL-6, B-cell stimulatory factor 2, BSF-2, CTL differentiation factor, CDF, hybridoma growth factor, Interferon beta-2, IFN-beta-2 |
| 5 | P05412 | JUN | Transcription factor Jun, activator protein 1, AP1, proto-oncogene c-Jun, transcription factor AP-1 subunit Jun, V-jun avian sarcoma virus 17 oncogene homolog, p39 |
| 6 | Q04206 | RELA | Transcription factor p65, Nuclear factor NF-kappa-B p65 subunit, nuclear factor of kappa light polypeptide gene enhancer in B-cells 3 |
| 7 | P40763 | STAT3 | Signal transducer and activator of transcription 3, acute-phase response factor |
| 8 | P01375 | TNF | Tumor necrosis factor, cachectin, TNF-alpha, tumor necrosis factor ligand superfamily member 2, TNF-a |
| 9 | P04637 | TP53 | Cellular tumor antigen p53, antigen NY-CO-13, phosphoprotein p53, tumor suppressor p53 |
| Protein | Ligand | Binding Affinity | Interacting Residues |
|---|---|---|---|
| HSP90AA1 | Melatonin | −8.2 | TYR-124, PHE-123, PHE-7, TRP-147, LEU-92 |
| AKT1 | 5-HTP | −7.9 | ASN-204, THR-211, TRP-80, ASP-292, LEU-210 |
| TP53 | Melatonin | −7.7 | ASP-1521, TYR-1502, TYR-1523, PHE-1519, TRP-1495 |
| MAPK3 | 5-HTP | −7.5 | MET-125, GLN-122, ALA-69, VAL-56, TYR-53 |
| TNF | 5-HTP | −7.2 | GLN-15, ARG-73, LEU-84, PHE-14, PHE-14, GLN-14 |
| ADORA1 | Melatonin | −6.9 | THR-88, LEU-82, ILE-106, ASN-86, PHE-168 |
| JUN | Melatonin | −6.9 | VAL-196, ASN-152, GLU-147, MET-149, LEU-206, ALA-91, VAL-78 |
| BCL2 | Melatonin | −6.8 | GLN-99, TYR-103, VAL-148, TYR-202, PHE-198, TRP-144 |
| FOS | Melatonin | −6.6 | ARG-143, GLU-170, LEU-179, GLN-271, HIS-21, MET-181, PRO-168, VAL-182, GLN-273 |
| TP53 | L-Theanine | −6.2 | ASP-1520, TRP-1495, ASP-1521, PHE-1519, TYR-1502 |
| HSP90AA1 | vitaminB6 | −5.7 | TYR-139, PHE-138, TRP-162, LEU-107, VAL-150, LEU-103 |
| TP53 | GABA | −4.8 | ASP-1521, TYR-1502, ASP-1521 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Lv, Y.; Qin, J.; Deng, W. Network Pharmacology Analysis of the Potential Pharmacological Mechanism of a Sleep Cocktail. Biomolecules 2024, 14, 630. https://doi.org/10.3390/biom14060630
Liang Y, Lv Y, Qin J, Deng W. Network Pharmacology Analysis of the Potential Pharmacological Mechanism of a Sleep Cocktail. Biomolecules. 2024; 14(6):630. https://doi.org/10.3390/biom14060630
Chicago/Turabian StyleLiang, Yuyun, Yanrong Lv, Jing Qin, and Wenbin Deng. 2024. "Network Pharmacology Analysis of the Potential Pharmacological Mechanism of a Sleep Cocktail" Biomolecules 14, no. 6: 630. https://doi.org/10.3390/biom14060630
APA StyleLiang, Y., Lv, Y., Qin, J., & Deng, W. (2024). Network Pharmacology Analysis of the Potential Pharmacological Mechanism of a Sleep Cocktail. Biomolecules, 14(6), 630. https://doi.org/10.3390/biom14060630