

Globospiramine Exhibits Inhibitory and Fungicidal Effects against Candida albicans via Apoptotic Mechanisms

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
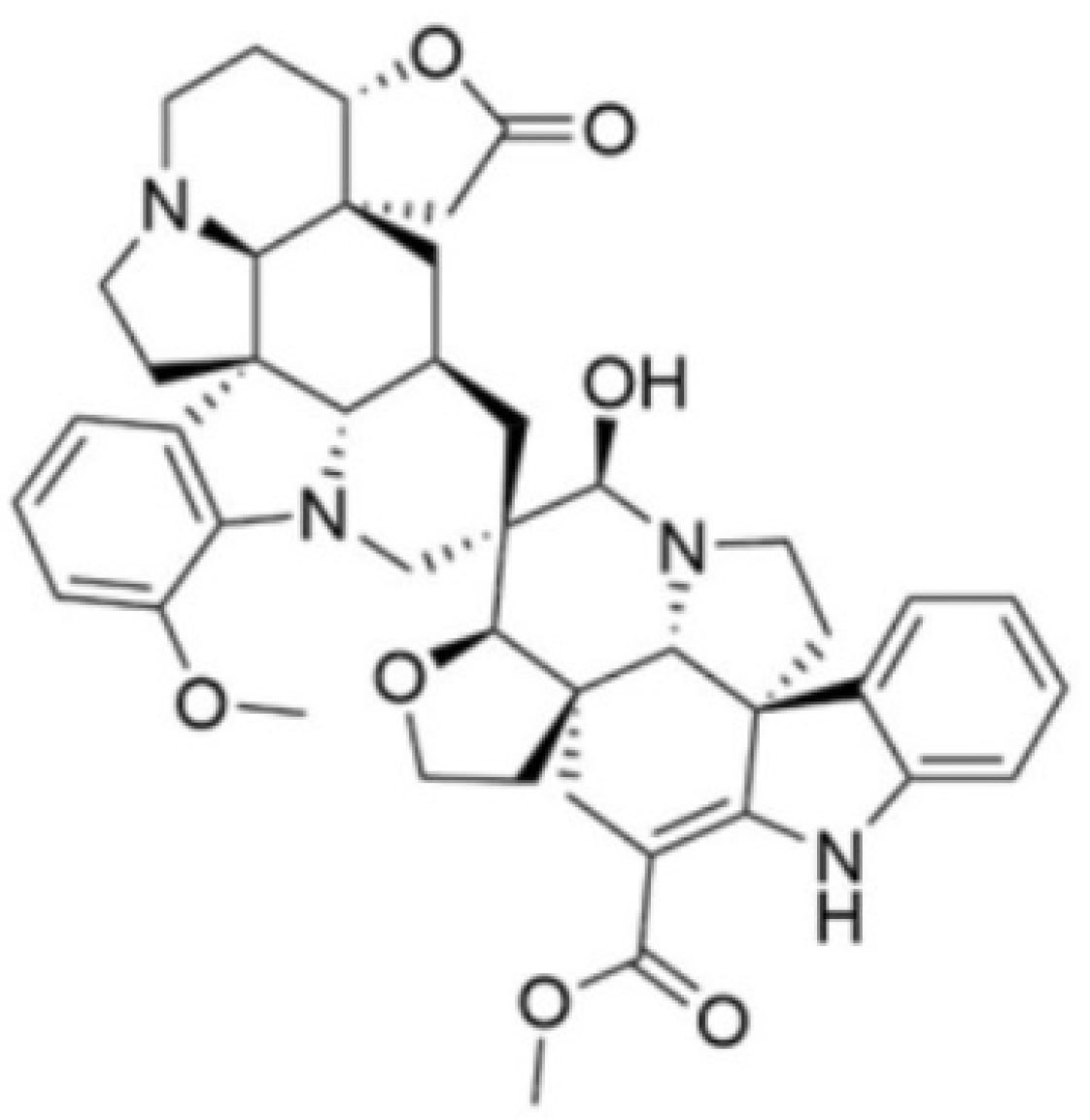
2.1. Test Compound
2.2. Colony-Forming Unit Viability Assay
2.3. MIC and MFC Determination
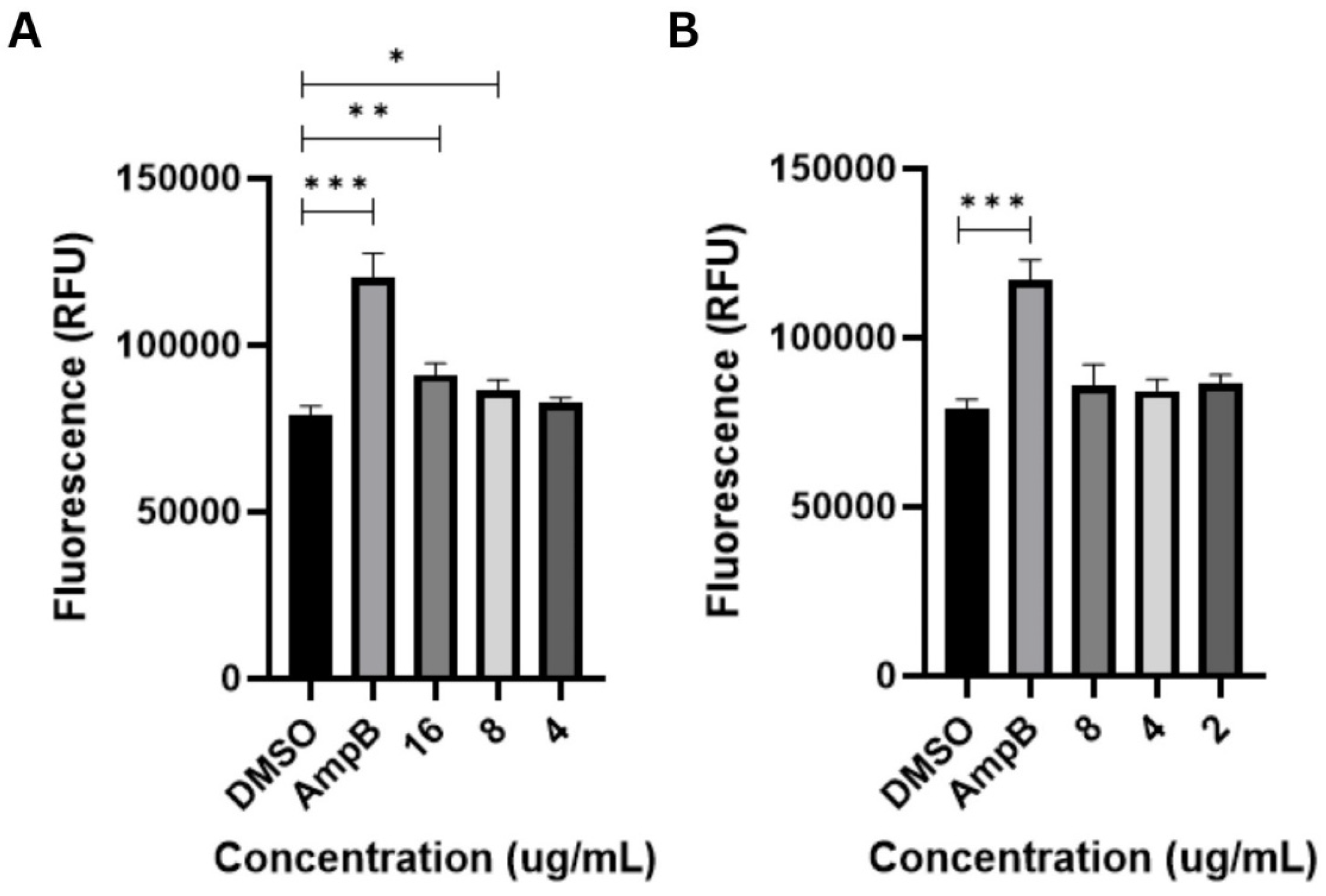
2.4. FAM-FLICA Poly-Caspase Assay
2.5. Molecular Docking against C. albicans Proteins
2.5.1. Ligand and Protein Preparation
2.5.2. Molecular Docking and Visualization of Interactions
2.6. Molecular Dynamics Simulations
3. Results
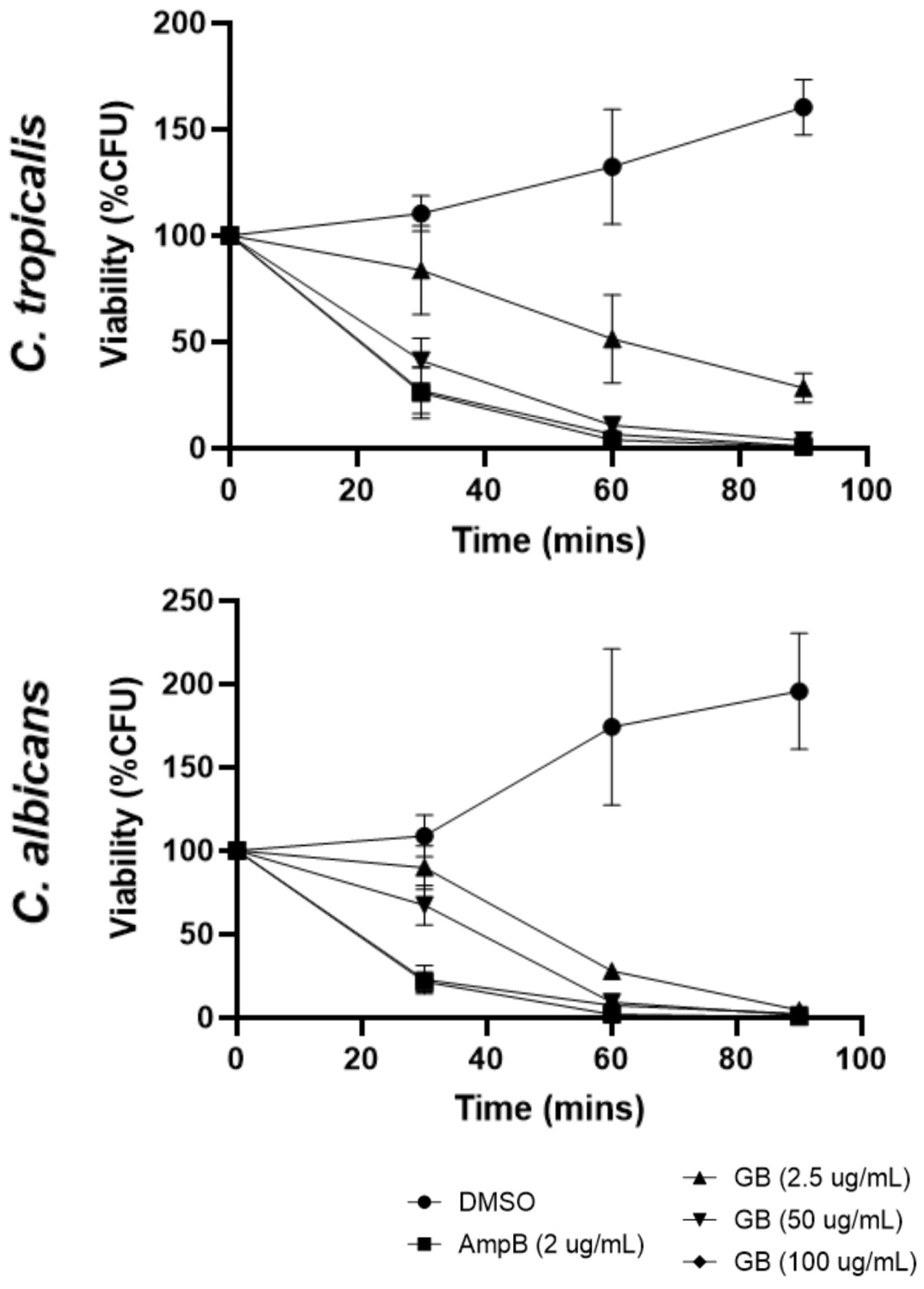
3.1. Effects of Globospiramine on C. albicans and C. tropicalis CFU Viability
3.2. MIC and MFC of Globospiramine versus C. albicans and C. tropicalis
3.3. Apoptosis-Inducing Activities of Globospiramine vs. C. albicans and C. tropicalis
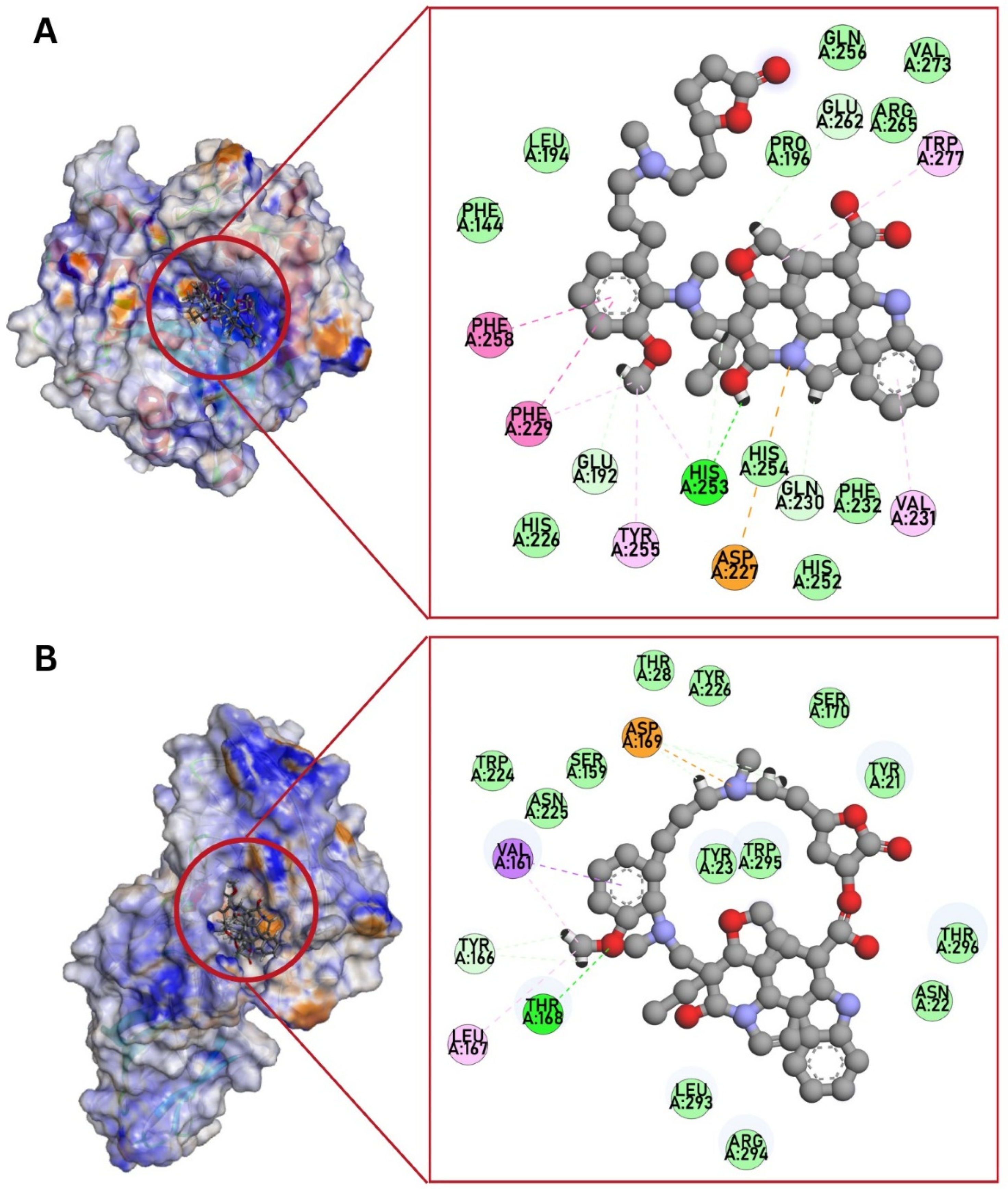
3.4. Molecular Docking against C. albicans Targets
3.5. Molecular Dynamics Simulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vandeputte, P.; Ferrari, S.; Coste, A.T. Antifungal resistance and new strategies to control fungal infections. Int. J. Microbiol. 2012, 2012, 713687. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-L.; Yu, S.-J.; Heitman, J.; Wellington, M.; Chen, Y.-L. New facets of antifungal therapy. Virulence 2016, 8, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Campoy, S.; Adrio, J.L. Antifungals. Biochem. Pharmacol. 2017, 133, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A. Antifungal drug resistance: Mechanisms, epidemiology, and consequences for treatment. Am. J. Med. 2012, 125, S3–S13. [Google Scholar] [CrossRef] [PubMed]
- Tobudic, S.; Kratzer, C.; Presterl, E. Azole-resistant Candida spp.—Emerging pathogens? Mycoses 2012, 55, 24–32. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed]
- Miceli, M.H.; Díaz, J.A.; Lee, S.A. Emerging opportunistic yeast infections. Lancet Infect. Dis. 2011, 11, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Eggimann, P.; Garbino, J.; Pittet, D. Epidemiology of Candida species infections in critically ill non-immunosuppressed patients. Lancet Infect. Dis. 2003, 3, 685–702. [Google Scholar] [CrossRef] [PubMed]
- Wächtler, B.; Citiulo, F.; Jablonowski, N.; Förster, S.; Dalle, F.; Schaller, M.; Wilson, D.; Hube, B. Candida albicans-epithelial interactions: Dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS ONE 2012, 7, e36952. [Google Scholar] [CrossRef]
- Martins, N.; Ferreira, I.C.; Barros, L.; Silva, S.; Henriques, M. Candidiasis: Predisposing factors, prevention, diagnosis and alternative treatment. Mycopathologia 2014, 177, 223–240. [Google Scholar] [CrossRef]
- Fuentefria, A.M.; Pippi, B.; Dalla Lana, D.F.; Donato, K.K.; de Andrade, S.F. Antifungals discovery: An insight into new strategies to combat antifungal resistance. Lett. Appl. Microbiol. 2018, 66, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Heard, S.C.; Wu, G.; Winter, J.M. Antifungal natural products. Curr. Opin. Biotechnol. 2021, 69, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Onishi, J.; Meinz, M.; Thompson, J.; Curotto, J.; Dreikorn, S.; Rosenbach, M.; Douglas, C.; Abruzzo, G.; Flattery, A.; Kong, L.; et al. Discovery of novel antifungal (1,3)-β-glucan synthase inhibitors. Antimicrob. Agents Chemother. 2000, 44, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Long, S.-Y.; Li, C.-L.; Hu, J.; Zhao, Q.-J.; Chen, D. Indole alkaloids from the aerial parts of Kopsia fruticosa and their cytotoxic, antimicrobial and antifungal activities. Fitoterapia 2018, 129, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Yordanov, M.; Dimitrova, P.; Patkar, S.; Saso, L.; Ivanovska, N. Inhibition of Candida albicans extracellular enzyme activity by selected natural substances and their application in Candida infection. Can. J. Microbiol. 2008, 54, 435–440. [Google Scholar] [CrossRef]
- Xu, G.-B.; He, G.; Bai, H.-H.; Yang, T.; Zhang, G.-L.; Wu, L.-W.; Li, G.-Y. Indole alkaloids from Chaetomium globosum. J. Nat. Prod. 2015, 78, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Li, W.; Chen, F.-F.; Zhang, J.-S.; Tang, Y.-Q.; Chen, L.; Tang, G.-H.; Yin, S. Monoterpene indole alkaloids from Rhazya stricta. Fitoterapia 2018, 128, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Anjum, K.; Kaleem, S.; Yi, W.; Zheng, G.; Lian, X.; Zhang, Z. Novel antimicrobial indolepyrazines A and B from the marine-associated Acinetobacter sp. ZZ1275. Mar. Drugs 2019, 17, 89. [Google Scholar] [CrossRef]
- Macabeo, A.P.; Vidar, W.S.; Chen, X.; Decker, M.; Heilmann, J.; Wan, B.; Franzblau, S.G.; Galvez, E.V.; Aguinaldo, M.A.; Cordell, G.A. Mycobacterium tuberculosis and cholinesterase inhibitors from Voacanga globosa. Eur. J. Med. Chem. 2011, 46, 3118–3123. [Google Scholar] [CrossRef]
- de Jesus, M.; Macabeo, A.; Ramos, J.; de Leon, V.; Asamitsu, K.; Okamoto, T. Voacanga globosa spirobisindole alkaloids exert antiviral activity in HIV latently infected cell lines by targeting the NF-ΚB CASCADE: In vitro and in silico investigations. Molecules 2022, 27, 1078. [Google Scholar] [CrossRef]
- Acebedo, A.R.; Amor, E.C.; Jacinto, S.D. Apoptosis-inducing activity of HPLC fraction from Voacanga globosa (Blanco) Merr. on the human colon carcinoma cell line, HCT116. Asian Pac. J. Cancer Prev. 2014, 15, 617–622. [Google Scholar] [CrossRef]
- Vital, P.G.; Rivera, W.L. Antimicrobial activity, cytotoxicity, and phytochemical screening of Voacanga globosa (Blanco) Merr. leaf extract (Apocynaceae). Asian Pac. J. Trop. Med. 2011, 4, 824–828. [Google Scholar] [CrossRef]
- Cascio, V.; Gittings, D.; Merloni, K.; Hurton, M.; Laprade, D.; Austriaco, N. S-adenosyl-L-methionine protects the probiotic yeast, Saccharomyces boulardii, from acid-induced cell death. BMC Microbiol. 2013, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J. The Effect of Acute Heavy Metal (Cu And Cd) Toxicity on ROS Generation, Apoptosis, and Intracellular Glutathione Levels in Saccharomyces cerevisiae. Master’s Thesis, Georgia State University, Atlanta, GA, USA, 2019. [Google Scholar]
- Poling, B.M. Differential Effects of Acute Cadmium, Copper, and Chromium Assault on Glutathione and Transcription Profiles in Saccharomyces cerevisiae. Master’s Thesis, Georgia State University, Atlanta, GA, USA, 2021. [Google Scholar]
- da Nóbrega Alves, D.; Monteiro, A.F.; Andrade, P.N.; Lazarini, J.G.; Abílio, G.M.; Guerra, F.Q.; Scotti, M.T.; Scotti, L.; Rosalen, P.L.; Castro, R.D. Docking Prediction, antifungal activity, anti-biofilm effects on Candida spp., and toxicity against human cells of cinnamaldehyde. Molecules 2020, 25, 5969. [Google Scholar] [CrossRef]
- Gurgel do Amaral Valente Sá, L.; da Silva, C.R.; Neto, J.B.; do Nascimento, F.B.; Barroso, F.D.; da Silva, L.J.; Cabral, V.P.; Barbosa, A.D.; Silva, J.; Marinho, E.S.; et al. Antifungal activity of etomidate against growing biofilms of fluconazole-resistant Candida spp. strains, binding to mannoproteins and molecular docking with the ALS3 protein. J. Med. Microbiol. 2020, 69, 1221–1227. [Google Scholar] [CrossRef]
- Bouamrane, S.; Khaldan, A.; Hajji, H.; El-mernissi, R.; Alaqarbeh, M.; Alsakhen, N.; Maghat, H.; Ajana, M.A.; Sbai, A.; Bouachrine, M.; et al. In silico identification of 1,2,4-triazoles as potential Candida albicans inhibitors using 3D-QSAR, molecular docking, molecular dynamics simulations, and ADMET profiling. Mol. Divers. 2022, 27, 2111–2132. [Google Scholar] [CrossRef]
- Manzano, J.A.; Cruz, C.L.; Quimque, M.T.; Macabeo, A.P. In silico potentials of Alpinia galanga constituents against human placental aromatase vital in postmenopausal estrogen-dependent breast cancer pathogenesis. Philipp. J. Sci. 2022, 151, 2101–2115. [Google Scholar] [CrossRef]
- Manzano, J.A.; Llames, L.C.; Macabeo, A.P. Tetrahydrobisbenzylisoquinoline alkaloids from Phaeanthus ophthalmicus inhibit target enzymes associated with type 2 diabetes and obesity. J. Appl. Pharm. Sci. 2023, 14, 230–237. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Brogi, S.; Rossi, S.; Ibba, R.; Butini, S.; Calderone, V.; Campiani, G.; Gemma, S. In silico analysis of peptide-based derivatives containing bifunctional warheads engaging prime and non-prime subsites to covalent binding SARS-COV-2 main protease (mpro). Computation 2022, 10, 69. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and testing of the OPLS all-atom force field on conformational energetics and properties of organic liquids. J. Am. Chem. Soc. 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1695–1697. [Google Scholar] [CrossRef] [PubMed]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Humphreys, D.D.; Friesner, R.A.; Berne, B.J. A multiple-time-step molecular dynamics algorithm for Macromolecules. J. Phys. Chem. 1994, 98, 6885–6892. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- da Silva, E.R.; Brogi, S.; Lucon-Júnior, J.F.; Campiani, G.; Gemma, S.; Maquiaveli, C. Dietary polyphenols rutin, taxifolin and quercetin related compounds target Leishmania amazonensis arginase. Food Funct. 2019, 10, 3172–3180. [Google Scholar] [CrossRef] [PubMed]
- Manzano, J.A.; Abellanosa, E.A.; Aguilar, J.P.; Brogi, S.; Yen, C.H.; Macabeo, A.P.; Austriaco, N. Globospiramine from Voacanga globosa Exerts Robust Cytotoxic and Antiproliferative Activities on Cancer Cells by Inducing Caspase-Dependent Apoptosis in A549 Cells and Inhibiting MAPK14 (p38α): In Vitro and Computational Investigations. Cells 2024, 13, 772. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Hou, S.-Y.; Zhang, T.-Y.; Wu, Y.-Y.; Zhang, M.-Y.; Yan, X.-M.; Xia, M.-Y.; Zhang, Y.-X. Cytotoxic and antimicrobial indole alkaloids from an endophytic fungus Chaetomium sp. SYP-F7950 of Panax notoginseng. RSC Adv. 2019, 9, 28754–28763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bai, X.; Yuwen, H.-S.; Guo, L.-L.; Liu, J.-W.; Hao, X.-J. Alkaloids from Tabernaemontana divaricata combined with fluconazole to overcome fluconazole resistance in Candida albicans. Bioorg. Chem. 2021, 107, 104515. [Google Scholar] [CrossRef]
- Lee, H.-S.; Yoon, K.-M.; Han, Y.-R.; Lee, K.J.; Chung, S.-C.; Kim, T.-I.; Lee, S.-H.; Shin, J.; Oh, K.-B. 5-hydroxyindole-type alkaloids, as Candida albicans isocitrate lyase inhibitors, from the tropical sponge Hyrtios sp. Bioorg. Med. Chem. Lett. 2009, 19, 1051–1053. [Google Scholar] [CrossRef]
- Leadsham, J.E.; Kotiadis, V.N.; Tarrant, D.J.; Gourlay, C.W. Apoptosis and the yeast actin cytoskeleton. Cell Death Differ. 2009, 17, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Al-Dhaheri, R.S.; Douglas, L.J. Apoptosis in Candida biofilms exposed to amphotericin B. J. Med. Microbiol. 2010, 59, 149–157. [Google Scholar] [CrossRef] [PubMed]
- da Silva, C.R.; de Andrade Neto, J.B.; de Sousa Campos, R.; Figueiredo, N.S.; Sampaio, L.S.; Magalhães, H.I.; Cavalcanti, B.C.; Gaspar, D.M.; de Andrade, G.M.; Lima, I.S.; et al. Synergistic effect of the flavonoid catechin, quercetin, or epigallocatechin gallate with fluconazole induces apoptosis in Candida tropicalis resistant to fluconazole. Antimicrob. Agents Chemother. 2014, 58, 1468–1478. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.; Alnajdy, D.; El-Keblawy, A.; Mosa, K.; Khoder, G.; Noreddin, A. Plants’ natural products as alternative promising anti-candida drugs. Pharmacogn. Rev. 2017, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Zhang, J.; Yu, L.; Wang, C.; Yang, Y.; Rong, X.; Xu, K.; Chu, M. Antifungal activity of coumarin against Candida albicans is related to apoptosis. Front. Cell. Infect. Microbiol. 2019, 8, 445. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-R.; You, Z.-L.; Chen, D.-D.; Hang, J.; Wang, Z.-B.; Ji, M.; Wang, L.-X.; Zhao, P.; Qiao, J.; Yun, C.-H.; et al. Structure of a fungal 1,3-β-glucan synthase. Sci. Adv. 2023, 9, eadh7820. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W. Echinocandin antifungal drugs. Lancet 2003, 362, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C.; Brown, A.J.P.; Gow, N.A.R. Antifungal agents: Mechanisms of action. Trends Microbiol. 2003, 11, 272–279. [Google Scholar] [CrossRef]
- Gow, N.A.; Latge, J.-P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 28513415. [Google Scholar] [CrossRef]
- Lima, S.L.; Colombo, A.L.; de Almeida Junior, J.N. Fungal cell wall: Emerging antifungals and drug resistance. Front. Microbiol. 2019, 10, 2573. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, Y. Antifungal activity of Salvia miltiorrhiza against Candida albicans is associated with the alteration of membrane permeability and (1,3)-β-D-glucan synthase activity. J. Microbiol. Biotechnol. 2016, 26, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S. Resistance to echinocandin-class antifungal drugs. Drug Resist. Updates 2007, 10, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.E.; Edlind, T.D. Topological and mutational analysis of Saccharomyces cerevisiae FKS1. Eukaryot. Cell 2012, 11, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Filler, S.G. Candida albicans ALS3, a multifunctional adhesin and Invasin. Eukaryot. Cell 2011, 10, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, L.L.; Cota, E. Candida albicans agglutinin-like sequence (ALS) Family Vignettes: A review of ALS protein structure and function. Front. Microbiol. 2016, 7, 280. [Google Scholar] [CrossRef] [PubMed]
- Kioshima, E.S.; Shinobu-Mesquita, C.S.; Abadio, A.K.; Felipe, M.S.; Svidzinski, T.I.; Maigret, B. Selection of potential anti-adhesion drugs by in silico approaches targeted to ALS3 from Candida albicans. Biotechnol. Lett. 2019, 41, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.R.; de Sardi, J.; Freires, I.A.; Silva, A.C.; Rosalen, P.L. In silico approaches for screening molecular targets in Candida albicans: A proteomic insight into drug discovery and development. Eur. J. Pharmacol. 2019, 842, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Cheng, S.; Clancy, C.J.; Nguyen, M.H. Caspofungin kills Candida albicans by causing both cellular apoptosis and necrosis. Antimicrob. Agents Chemother. 2013, 57, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhang, B.; Li, J.; Zhang, B.; Wang, H.; Li, M. Endoplasmic reticulum-derived reactive oxygen species (ROS) is involved in toxicity of cell wall stress to Candida albicans. Free Radic. Biol. Med. 2016, 99, 572–583. [Google Scholar] [CrossRef]
- Maurya, I.K.; Pathak, S.; Sharma, M.; Sanwal, H.; Chaudhary, P.; Tupe, S.; Deshpande, M.; Chauhan, V.S.; Prasad, R. Antifungal activity of novel synthetic peptides by accumulation of reactive oxygen species (ROS) and disruption of cell wall against Candida albicans. Peptides 2011, 32, 1732–1740. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, J.; Sun, J.; Zhu, X.; Zhou, L.; Lu, Z.; Lu, Y. C16-Fengycin a affect the growth of Candida albicans by destroying its cell wall and accumulating reactive oxygen species. Appl. Microbiol. Biotechnol. 2019, 103, 8963–8975. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.J.; Sudbery, I.; Ramsdale, M. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proc. Natl. Acad. Sci. USA 2003, 100, 14327–14332. [Google Scholar] [CrossRef] [PubMed]
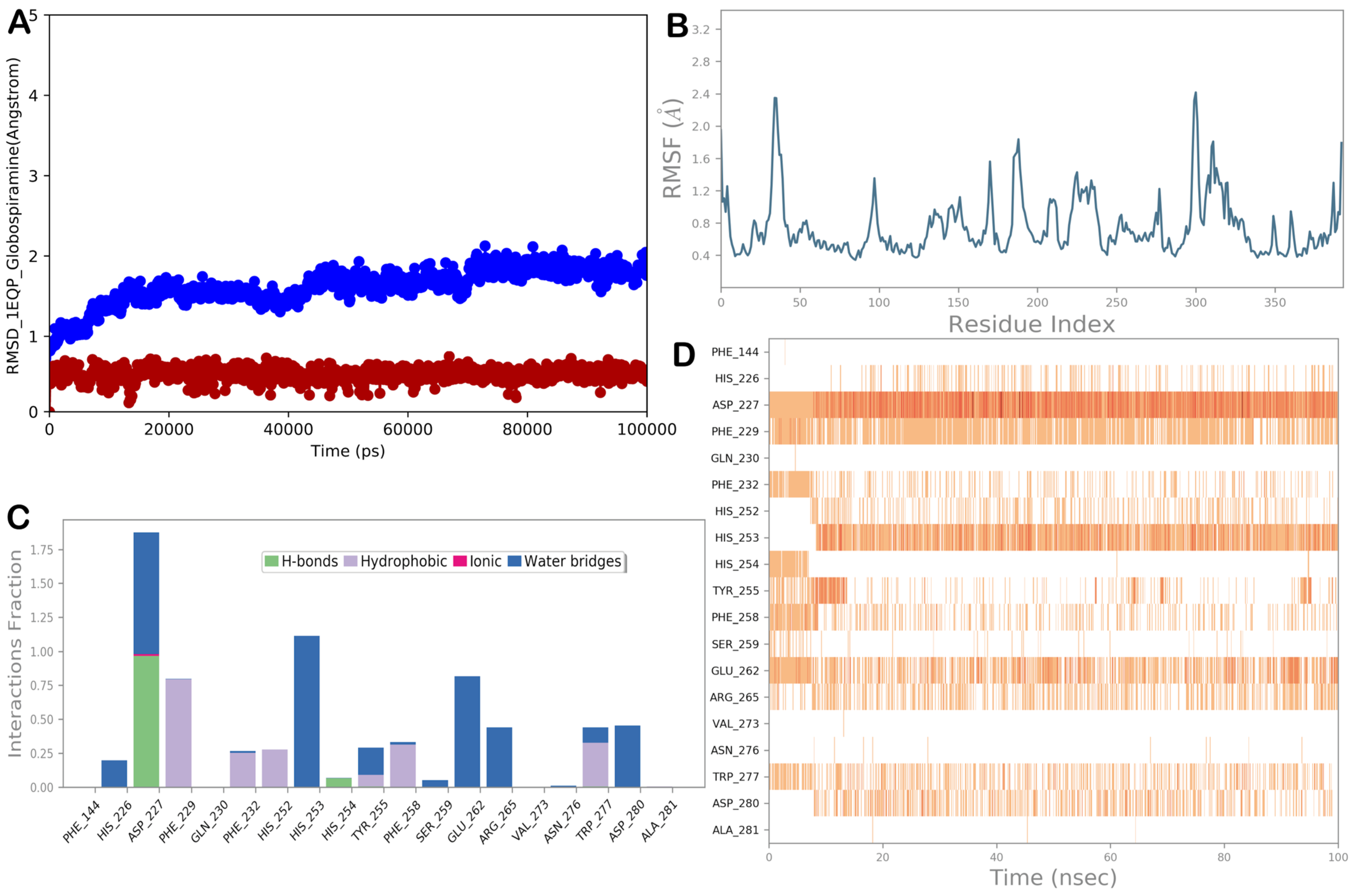
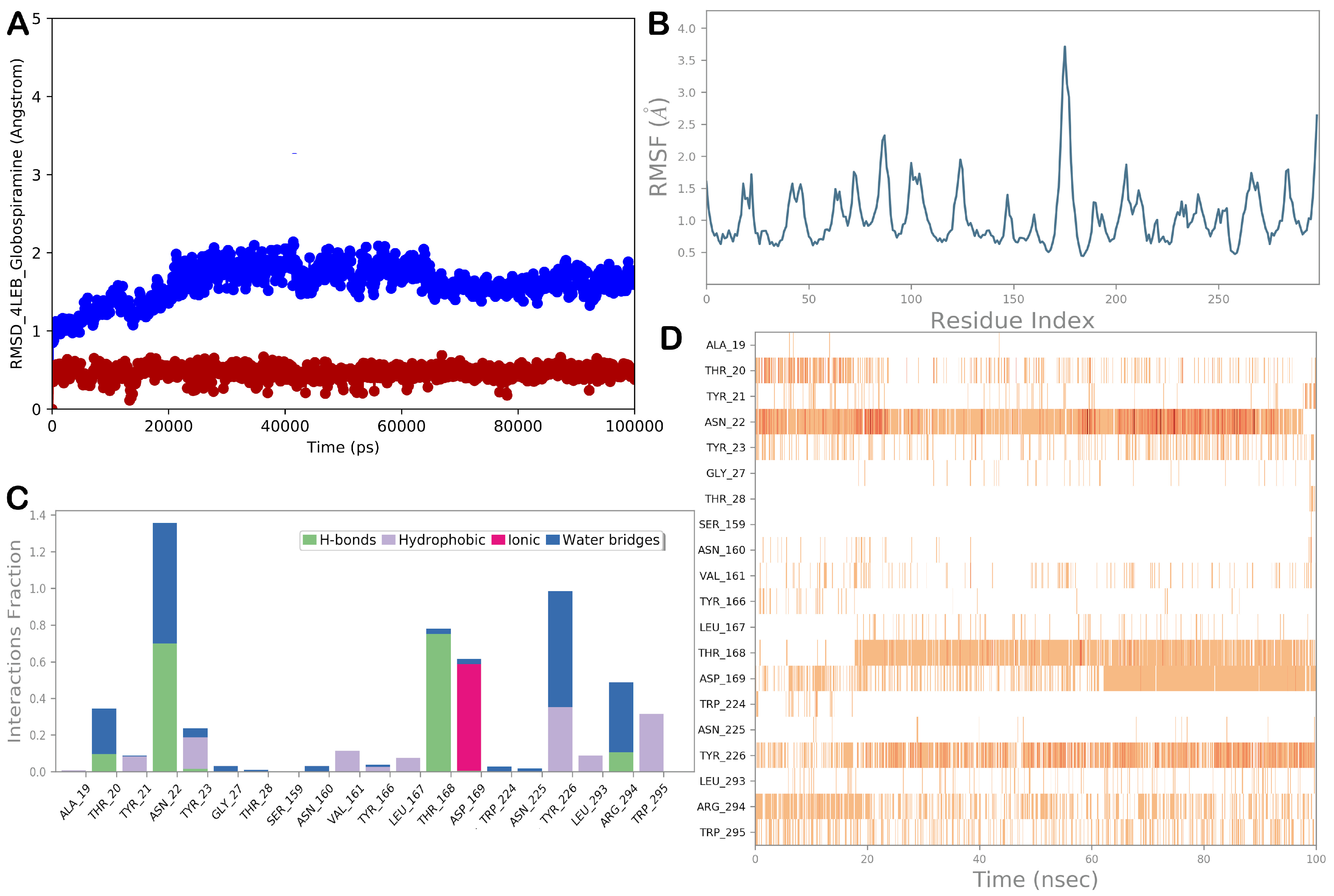

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Globospiramine | Amphotericin B | |
|---|---|---|
| MIC (µg/mL) | ||
| C. albicans | 8.0 | 0.5 |
| C. tropicalis | 4.0 | 0.5 |
| MFC (µg/mL) | ||
| C. albicans | 8.0 | 0.5 |
| C. tropicalis | >64.0 | 1.0 |
| PDB IDs | Globospiramine | Positive Controls | ||||||
|---|---|---|---|---|---|---|---|---|
| Caspofungin | Amphotericin B | Co-Crystallized Ligand/Inhibitor | ||||||
| BE (kcal/ mol) | Interactions | BE (kcal/ mol) | Interactions | BE (kcal/ mol) | Interactions | BE (kcal/ mol) | Interactions | |
| 1EQP (1,3-β-glucan synthase) | −10.5 | His253 (H-bond), Phe258, Phe229 (pi–pi stacked), Trp277, Val231, Tyr255, Phe229, His253 (pi–alkyl), Glu192, His253, Gln230, Glu262 (C-H bond) | −8.0 | Trp277, Gln230 (H-bond), Asp227 (salt bridge), His254, Glu262 (C-H bond), Val273, Phe258, Phe144, Tyr255, Trp373 (alkyl, pi–alkyl), His253, Arg265 (unfavorable interaction) | −8.8 | Asn305, Asp151 (H-bond), Phe258, Phe144 (pi–alkyl), Tyr 153 (unfavorable donor–donor, C-H bond) | - | - |
| 4QUV (δ-14-sterol reductase) | −9.5 | Arg324, His320 (H-bond), Val96, His320 (pi–sigma), Leu253, Met99 (alkyl), Arg106, Arg323 (unfavorable positive–positive) | −7.2 | Arg106, Arg323, Arg324, Lys406 (H-bond), Tyr414, Trp352, Leu346, Cys403, Trp411, Lys319, Val96 (alkyl, pi–alkyl), His320 (pi–pi stacked), Gln97 (C-H bond), Arg324 (unfavorable positive–positive) | −7.5 | Gln97, Glu250, Arg323, Arg324, Gly343 (H-bond), Met99, Leu253 (alkyl), Arg106 (unfavorable positive–positive) | −9.5 | His248, Arg313, Thr254, Lys259, Lys319, Trp256, Arg395, Asn316, Thr255 (H-bond), Asp244, Asp399, Arg395 (attractive charge, pi–cation), Glu201 (C-H bond), Lys319 (unfavorable positive–positive), Tyr245 (pi–pi T-shaped), Arg398, Val252 (pi–alkyl, alkyl) |
| 5TZ1 (lanosterol 14-alpha demethylase or CYP51) | −7.4 | Arg469 (H-bond), Glu444 (attractive charge), Val452, Val454 (alkyl), Ser453, Lys451 (C-H bond) | −5.7 | Met508, Pro462, His468, Leu439, Leu471, Gly303 (H-bond), His468 (C-H bond), Ile304 (pi–sigma), Leu87, Phe233, Tyr64, Phe380, Phe228, Val509, Leu150, Ile304, Ile131, His377, Pro230, Leu88, Lys90 (pi–alkyl, alkyl), Arg381, Tyr132, Lys143 (unfavorable interactions) | −3.3 | Phe463 (H-bond), His468 (C-H bond), Tyr118 (pi–lone pair), Leu376, Ile379, Ala146, Ile304, Leu204, Phe475 (alkyl, pi–alkyl), Cys470, Ile379, Gly464, Arg381, Thr311, Phe475, Leu150, Ile471, Tyr132 (unfavorable bonds) | −10.6 | Gly303, Ile304 (C-H bond), Ser507, His377 (halogen), Tyr118, Tyr132 (pi–pi), Leu121, Phe233, Leu376, Pro230, Ile304, Ile131, Lys143 (alkyl, pi–alkyl) |
| 5UIV (thymidylate kinase) | −9.4 | Gly155, Asp91, Arg39 (H-bond), Asp13, Arg39, Glu159 (pi–cation/pi–anion/salt bridge), Glu159, Ser18 (C-H bond), Lys17 (pi–alkyl) | −8.2 | Ser18, Asp13, Asp91, Arg92, Lys17, Lys35, Arg39, Gly157 Gly155 (H-bond), Glu162, Glu159 (salt bridge, attractive charge), Asp13, Asp91, Lys35, Gly155 (C-H bond), Ile196, Arg153, Lys17, Arg39, Val199 (alkyl, pi–alkyl) | −7.7 | Arg92, Lys35, SerA (H-bond), Glu162, Gln159 (salt bridge, attractive charge), Pro37 (alkyl), Asp13 (C-H bond), Ser18 (unfavorable donor–donor) | −8.9 | Arg92, Lys17, Arg14, Ser18, Gly16 (H-bond), Glu159, Asp91, Asp13 (attractive charge, pi–anion), Lys35 (unfavorable donor–donor), Tyr100 (pi–pi), Leu51 (pi–alkyl) |
| 4LEB (Als3 adhesin) | −10.6 | Thr168 (H-bond), Asp169 (attractive charge), Asp169, Tyr166 (C-H bond), Val161 (pi–sigma), Val161, Leu167 (alkyl) | −6.5 | Thr168, Tyr226, Thr20, Asn22 (H-bond), Trp295 (pi–cation), Pro29, Arg171, Tyr21 (alkyl, pi–alkyl), Asn22 (pi–donor H-bond) | −7.7 | Asn22 (H-bond), Tyr226 (pi–alkyl), Arg294 (unfavorable positive–positive) | - | - |
| 2Y7L (Als9-2) | −8.1 | Thr293 (C-H bond), Trp294 (pi–cation), Tyr21, Pro160, Val161 (pi–alkyl) | −6.4 | Thr168 (H-bond), Arg171, Val22, Pro160, Val161, Ile167, Tyr23, Phe225, Pro29 (alkyl, pi–alkyl) | −7.1 | Glu86, Ser210, Asn213 (H-bond), Asn211 (C-H bond), Tyr261 (pi–alkyl) | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manzano, J.A.H.; Brogi, S.; Calderone, V.; Macabeo, A.P.G.; Austriaco, N. Globospiramine Exhibits Inhibitory and Fungicidal Effects against Candida albicans via Apoptotic Mechanisms. Biomolecules 2024, 14, 610. https://doi.org/10.3390/biom14060610
Manzano JAH, Brogi S, Calderone V, Macabeo APG, Austriaco N. Globospiramine Exhibits Inhibitory and Fungicidal Effects against Candida albicans via Apoptotic Mechanisms. Biomolecules. 2024; 14(6):610. https://doi.org/10.3390/biom14060610
Chicago/Turabian StyleManzano, Joe Anthony H., Simone Brogi, Vincenzo Calderone, Allan Patrick G. Macabeo, and Nicanor Austriaco. 2024. "Globospiramine Exhibits Inhibitory and Fungicidal Effects against Candida albicans via Apoptotic Mechanisms" Biomolecules 14, no. 6: 610. https://doi.org/10.3390/biom14060610
APA StyleManzano, J. A. H., Brogi, S., Calderone, V., Macabeo, A. P. G., & Austriaco, N. (2024). Globospiramine Exhibits Inhibitory and Fungicidal Effects against Candida albicans via Apoptotic Mechanisms. Biomolecules, 14(6), 610. https://doi.org/10.3390/biom14060610