
IgG N-Glycosylation Is Altered in Coronary Artery Disease


 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Methods
2.2.1. Isolation of IgG from Human Plasma
2.2.2. Deglycosylation, Labelling, and Purification of IgG N-Glycans
2.2.3. HILIC-UHPLC-FLR Analysis of 2-AB Labelled IgG N-Glycans
2.2.4. Statistical Analysis
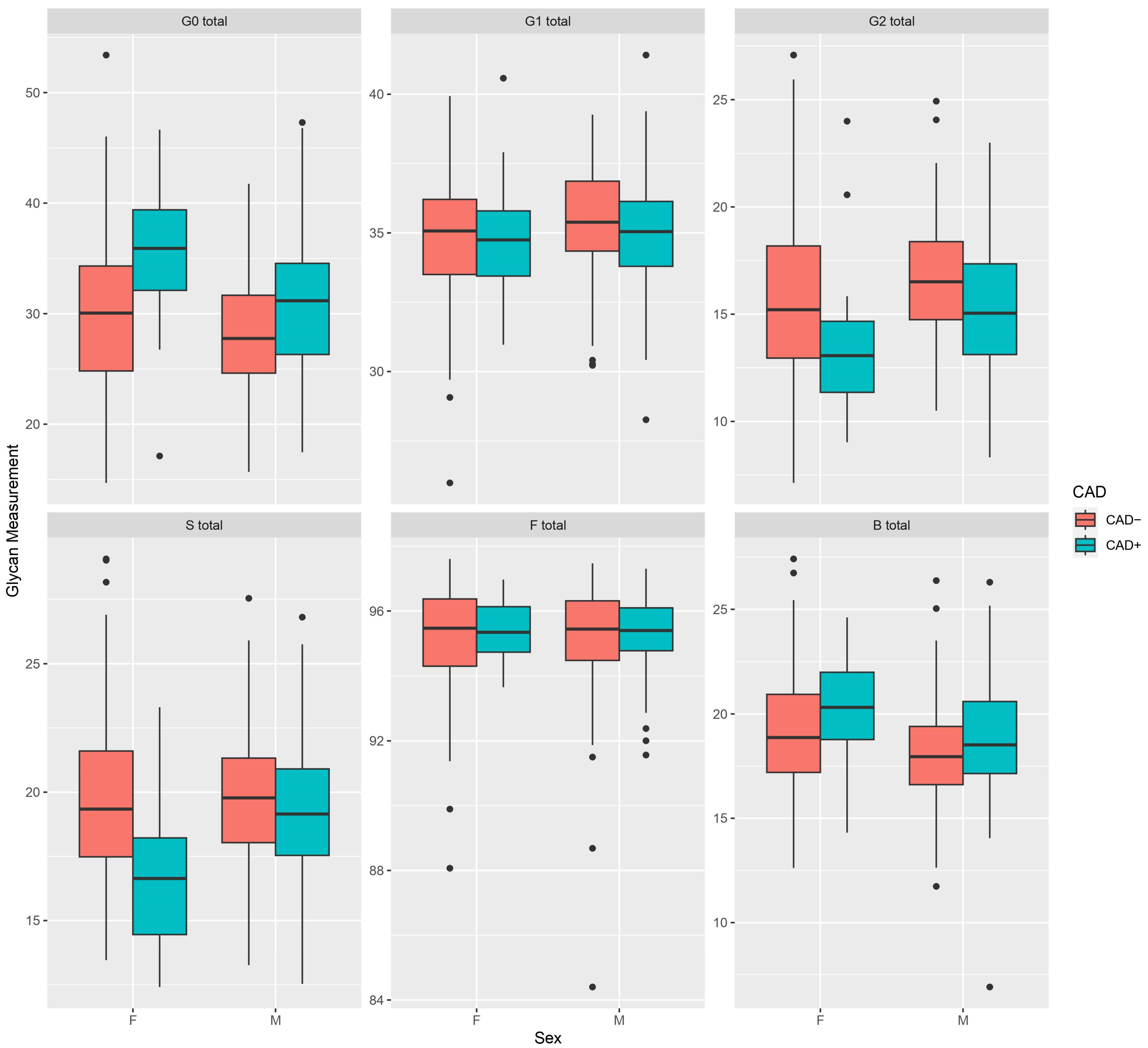
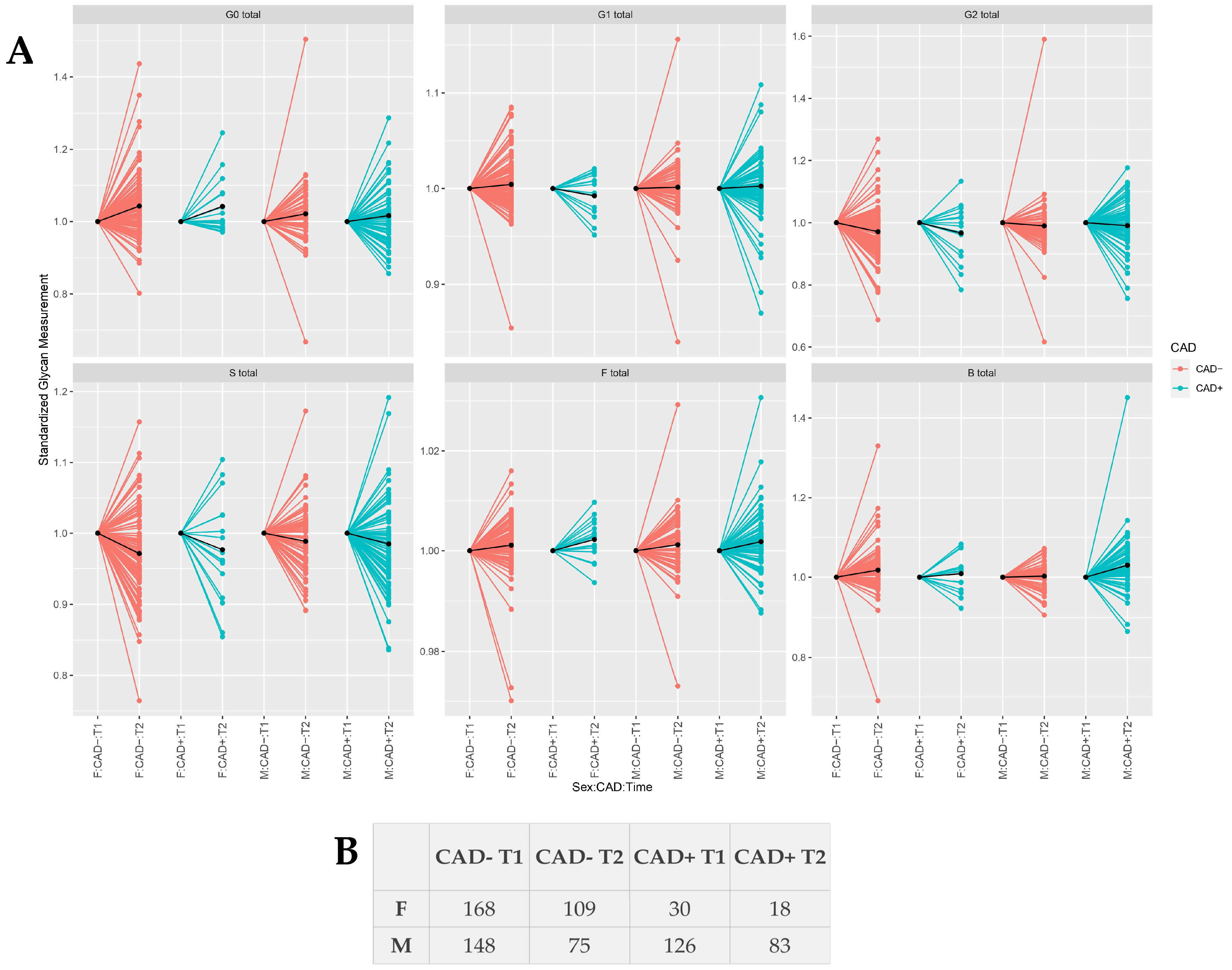
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.; Benjamin, E.J.; Benziger, C.P.; et al. Global burden of cardiovascular diseases and risk factors, 1990–2019: Update from the GBD 2019 study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Hajar, R. Risk factors for coronary artery disease: Historical perspectives. Heart Views Off. J. Gulf Heart Assoc. 2017, 18, 109. [Google Scholar] [CrossRef] [PubMed]
- Greenland, P.; Knoll, M.D.; Stamler, J.; Neaton, J.D.; Dyer, A.R.; Garside, D.B.; Wilson, P.W. Major risk factors as antecedents of fatal and nonfatal coronary heart disease events. JAMA 2003, 290, 891–897. [Google Scholar] [CrossRef]
- Ridker, P.M.; Buring, J.E.; Rifai, N.; Cook, N.R. Development and validation of improved algorithms for the assessment of global cardiovascular risk in women: The reynolds risk score. JAMA 2007, 297, 611–619. [Google Scholar] [CrossRef]
- Wilson, P.W.F.; Pencina, M.; Jacques, P.; Selhub, J.; D’Agostino, R.; O’Donnell, C.J. C-reactive protein and reclassification of cardiovascular risk in the framingham heart study. Circ. Cardiovasc. Qual. Outcomes 2008, 1, 92–97. [Google Scholar] [CrossRef]
- Naghavi, M.; Falk, E.; Hecht, H.S.; Jamieson, M.J.; Kaul, S.; Berman, D.; Fayad, Z.; Budoff, M.J.; Rumberger, J.; Naqvi, T.Z.; et al. From vulnerable plaque to vulnerable patient-part III: Executive summary of the screening for heart attack prevention and education (SHAPE) task force report. Am. J. Cardiol. 2006, 98, 2–15. [Google Scholar] [CrossRef]
- Deidda, M.; Noto, A.; Dessalvi, C.C.; Andreini, D.; Andreotti, F.; Ferrannini, E.; Latini, R.; Maggioni, A.P.; Magnoni, M.; Mercuro, G. Why do high-risk patients develop or not develop coronary artery disease? Metabolic insights from the CAPIRE study. Metabolites 2022, 12, 123. [Google Scholar] [CrossRef]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef]
- Marth, J.D.; Grewal, P.K. Mammalian glycosylation in immunity. Nat. Rev. Immunol. 2008, 8, 874–887. [Google Scholar] [CrossRef]
- Gudelj, I.; Lauc, G.; Pezer, M. Immunoglobulin G glycosylation in aging and diseases. Cell. Immunol. 2018, 333, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Clerc, F.; Reiding, K.R.; Jansen, B.C.; Kammeijer, G.S.M.; Bondt, A.; Wuhrer, M. Human plasma protein N-glycosylation. Glycoconj. J. 2016, 33, 309–343. [Google Scholar] [CrossRef]
- Gudelj, I.; Lauc, G. Protein N-glycosylation in cardiovascular diseases and related risk factors. Curr. Cardiovasc. Risk Rep. 2018, 12, 16. [Google Scholar] [CrossRef]
- Scott, D.W.; Patel, R.P. Endothelial heterogeneity and adhesion molecules N-glycosylation: Implications in leukocyte trafficking in inflammation. Glycobiology 2013, 23, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.W.; Chen, J.; Chacko, B.K.; Traylor, J.G.; Orr, A.W.; Patel, R.P. Role of endothelial N-glycan mannose residues in monocyte recruitment during atherogenesis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, e51–e59. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Xia, L. P-selectin glycoprotein ligand-1 plays a crucial role in the selective recruitment of leukocytes into the atherosclerotic arterial wall. Trends Cardiovasc. Med. 2009, 19, 140–145. [Google Scholar] [CrossRef]
- Krishnan, S.; Huang, J.; Lee, H.; Guerrero, A.; Berglund, L.; Anuurad, E.; Lebrilla, C.B.; Zivkovic, A.M. Combined high-density lipoprotein proteomic and glycomic profiles in patients at risk for coronary artery disease. J. Proteome Res. 2015, 14, 5109–5118. [Google Scholar] [CrossRef]
- Sukhorukov, V.; Gudelj, I.; Pučić-Baković, M.; Zakiev, E.; Orekhov, A.; Kontush, A.; Lauc, G. Glycosylation of human plasma lipoproteins reveals a high level of diversity, which directly impacts their functional properties. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 643–653. [Google Scholar] [CrossRef]
- Gabay, C.; Kushner, I. Acute-phase proteins and other systemic responses to inflammation. N. Engl. J. Med. 1999, 340, 448–454. [Google Scholar] [CrossRef]
- Arnold, J.N.; Saldova, R.; Abd Hamid, U.M.; Rudd, P.M. Evaluation of the serum N-linked glycome for the diagnosis of cancer and chronic inflammation. Proteomics 2008, 8, 3284–3293. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef]
- Radovani, B.; Gudelj, I. N-Glycosylation and Inflammation; The not-so-sweet relation. Front. Immunol. 2022, 13, 3235. [Google Scholar] [CrossRef] [PubMed]
- Menni, C.; Gudelj, I.; MacDonald-Dunlop, E.; Mangino, M.; Zierer, J.; Bešić, E.; Joshi, P.K.; Trbojević-Akmačić, I.; Chowienczyk, P.J.; Spector, T.D.; et al. Glycosylation profile of immunoglobulin G Is cross-sectionally associated with cardiovascular disease risk score and subclinical atherosclerosis in two independent cohorts. Circ. Res. 2018, 122, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Birukov, A.; Plavša, B.; Eichelmann, F.; Kuxhaus, O.; Hoshi, R.A.; Rudman, N.; Štambuk, T.; Trbojević-Akmačić, I.; Schiborn, C.; Morze, J.; et al. Immunoglobulin G N-glycosylation signatures in incident type 2 diabetes and cardiovascular disease. Diabetes Care 2022, 45, 2729–2736. [Google Scholar] [CrossRef]
- Wu, Z.; Guo, Z.; Zheng, Y.; Wang, Y.; Zhang, H.; Pan, H.; Li, Z.; Balmer, L.; Li, X.; Tao, L.; et al. IgG N-glycosylation cardiovascular age tracks cardiovascular risk beyond calendar age. Engineering 2023. [Google Scholar] [CrossRef]
- Magnoni, M.; Andreini, D.; Gorini, M.; Moccetti, T.; Modena, M.G.; Canestrari, M.; Berti, S.; Casolo, G.; Gabrielli, D.; Marraccini, P.; et al. Coronary atherosclerosis in outlier subjects at the opposite extremes of traditional risk factors: Rationale and preliminary results of the coronary atherosclerosis in outlier subjects: Protective and novel individual risk factors evaluation (CAPIRE) stud. Am. Heart J. 2016, 173, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Austen, W.G.; Edwards, J.E.; Frye, R.L.; Gensini, G.G.; Gott, V.L.; Griffith, L.S.; McGoon, D.C.; Murphy, M.L.; Roe, B.B. A reporting system on patients evaluated for coronary artery disease. Report of the ad hoc committee for grading of coronary artery disease, council on cardiovascular surgery, American heart association. Circulation 1975, 51, 5–40. [Google Scholar] [CrossRef]
- Pučić, M.; Knežević, A.; Vidič, J.; Adamczyk, B.; Novokmet, M.; Polašek, O.; Gornik, O.; Šupraha-Goreta, S.; Wormald, M.R.; Redžic, I.; et al. High throughput isolation and glycosylation analysis of IgG-variability and heritability of the IgG glycome in three isolated human populations. Mol. Cell. Proteom. 2011, 10, M111.010090. [Google Scholar] [CrossRef]
- Colafella, K.M.M.; Denton, K.M. Sex-specific differences in hypertension and associated cardiovascular disease. Nat. Rev. Nephrol. 2018, 14, 185–201. [Google Scholar] [CrossRef]
- Wittenbecher, C.; Štambuk, T.; Kuxhaus, O.; Rudman, N.; Vučković, F.; Štambuk, J.; Schiborn, C.; Rahelić, D.; Dietrich, S.; Gornik, O.; et al. Plasma N-glycans as emerging biomarkers of cardiometabolic risk: A prospective investigation intheepic-potsdamcohortstudy. Diabetes Care 2020, 43, 661–668. [Google Scholar] [CrossRef]
- Ercan, A.; Kohrt, W.M.; Cui, J.; Deane, K.D.; Pezer, M.; Yu, E.W.; Hausmann, J.S.; Campbell, H.; Kaiser, U.B.; Rudd, P.M.; et al. Estrogens regulate glycosylation of IgG in women and men. JCI Insight 2017, 2, e89703. [Google Scholar] [CrossRef]
- Deriš, H.; Kifer, D.; Cindrić, A.; Petrović, T.; Cvetko, A.; Trbojević-Akmačić, I.; Kolčić, I.; Polašek, O.; Newson, L.; Spector, T.; et al. Immunoglobulin G glycome composition in transition from premenopause to postmenopause. iScience 2022, 25, 103897. [Google Scholar] [CrossRef] [PubMed]
- McKinlay, S.M.; Brambilla, D.J.; Posner, J.G. The normal menopause transition. Maturitas 1992, 14, 103–115. [Google Scholar] [CrossRef]
- Jackson, R.; Lawes, C.M.; Bennett, D.A.; Milne, R.J.; Rodgers, A. Treatment with drugs to lower blood pressure and blood cholesterol based on an individual’s absolute cardiovascular risk. Lancet 2005, 365, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, M.; Chiodini, P.; Chambless, L.E.; Cesana, G.; Vanuzzo, D.; Panico, S.; Sega, R.; Pilotto, L.; Palmieri, L.; Giampaoli, S.; et al. Prediction of coronary events in a low incidence population. assessing accuracy of the CUORE cohort study prediction equation. Int. J. Epidemiol. 2005, 34, 413–421. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, R.B.; Vasan, R.S.; Pencina, M.J.; Wolf, P.A.; Cobain, M.; Massaro, J.M.; Kannel, W.B. General cardiovascular risk profile for use in primary care: The framingham heart study. Circulation 2008, 117, 743–753. [Google Scholar] [CrossRef]
- Wong, N.D.; Budoff, M.J.; Ferdinand, K.; Graham, I.M.; Michos, E.D.; Reddy, T.; Shapiro, M.D.; Toth, P.P. Atherosclerotic cardiovascular disease risk assessment: An American society for preventive cardiology clinical practice statement. Am. J. Prev. Cardiol. 2022, 10, 100335. [Google Scholar] [CrossRef]
- Goff, D.C.; Lloyd-Jones, D.M.; Bennett, G.; Coady, S.; D’Agostino, R.B.; Gibbons, R.; Greenland, P.; Lackland, D.T.; Levy, D.; O’Donnell, C.J.; et al. 2013 ACC/AHA guideline on the assessment of cardiovascular risk: A report of the american college of cardiology/American heart association task force on practice guidelines. Circulation 2014, 129, 49–73. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| All Population (n = 472) | CAD− (n = 316) | CAD+ (n = 156) | |
|---|---|---|---|
| Age (y), mean ± SD | 59.9 ± 8.4 | 58.1 ± 8.3 | 63.7 ± 7.4 |
| Women, % | 42 | 53 | 19 |
| BMI (kg/m2), mean ± SD | 26 ± 4.2 | 26 ± 3.9 | 27.7 ± 4.5 |
| Diabetes, % | 12 | 7 | 21 |
| Smoking, % | 26 | 21 | 37 |
| Glycan Traits | Model 1 ** | Model 2 *** | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Women | Men | Women | Men | |||||||||||||
| Effect | SE | p-Value | padj-Value * | Effect | SE | p-Value | padj-Value * | Effect | SE | p-Value | padj-Value * | Effect | SE | p-Value | padj-Value * | |
| S total | −0.82 | 0.1705 | 1.99 × 10−6 | 6.78 × 10−5 | 0.03 | 0.1296 | 8.05 × 10−1 | 9.58 × 10−1 | −0.63 | 0.1781 | 3.89 × 10−4 | 2.65 × 10−2 | 0.19 | 0.1306 | 1.40 × 10−1 | 4.92 × 10−1 |
| G0 total | 0.48 | 0.1599 | 2.51 × 10−3 | 2.26 × 10−2 | 0.1 | 0.1241 | 4.03 × 10−1 | 7.93 × 10−1 | 0.4 | 0.1708 | 1.80 × 10−2 | 1.53 × 10−1 | −0.05 | 0.1249 | 6.81 × 10−1 | 9.38 × 10−1 |
| G1 total | −0.03 | 0.204 | 8.76 × 10−1 | 9.58 × 10−1 | −0.18 | 0.1293 | 1.56 × 10−1 | 4.25 × 10−1 | −0.12 | 0.2207 | 5.65 × 10−1 | 9.11 × 10−1 | −0.11 | 0.1335 | 3.90 × 10−1 | 8.21 × 10−1 |
| G2 total | −0.28 | 0.1584 | 7.29 × 10−2 | 2.70 × 10−1 | −0.17 | 0.1224 | 1.74 × 10−1 | 4.36 × 10−1 | −0.26 | 0.1707 | 1.18 × 10−1 | 4.92 × 10−1 | −0.05 | 0.1241 | 6.77 × 10−1 | 9.38 × 10−1 |
| F total | −0.02 | 0.2032 | 9.38 × 10−1 | 9.80 × 10−1 | 0 | 0.1327 | 9.80 × 10−1 | 9.80 × 10−1 | −0.01 | 0.2183 | 9.75 × 10−1 | 9.91 × 10−1 | 0.01 | 0.1386 | 9.54 × 10−1 | 9.91 × 10−1 |
| B total | 0.21 | 0.1915 | 2.58 × 10−1 | 5.65 × 10−1 | 0.19 | 0.1308 | 1.48 × 10−1 | 4.20 × 10−1 | −0.06 | 0.1993 | 7.75 × 10−1 | 9.49 × 10−1 | 0.06 | 0.131 | 6.39 × 10−1 | 9.38 × 10−1 |
| FGS/(F + FG + FGS) | −0.71 | 0.1702 | 3.44 × 10−5 | 7.81 × 10−4 | 0.01 | 0.1285 | 9.66 × 10−1 | 9.80 × 10−1 | −0.56 | 0.18 | 1.66 × 10−3 | 2.91 × 10−2 | 0.13 | 0.1312 | 3.09 × 10−1 | 8.09 × 10−1 |
| FBS1/(FS1 + FBS1) | −0.03 | 0.184 | 8.81 × 10−1 | 9.58 × 10−1 | 0.02 | 0.1301 | 8.86 × 10−1 | 9.58 × 10−1 | −0.06 | 0.1988 | 7.73 × 10−1 | 9.49 × 10−1 | 0.00 | 0.136 | 9.91 × 10−1 | 9.91 × 10−1 |
| FBS1/FS1 | −0.03 | 0.184 | 8.81 × 10−1 | 9.58 × 10−1 | 0.02 | 0.1301 | 8.86 × 10−1 | 9.58 × 10−1 | −0.06 | 0.1988 | 7.73 × 10−1 | 9.49 × 10−1 | 0.00 | 0.136 | 9.91 × 10−1 | 9.91 × 10−1 |
| Glycan Traits | Model 1 ** | Model 2 *** | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Women | Men | Women | Men | |||||||||||||
| Effect | SE | p-Value | padj-Value * | Effect | SE | p-Value | padj-Value * | Effect | SE | p-Value | padj-Value * | Effect | SE | p-Value | padj-Value * | |
| S total | 0 | 0.1025 | 9.98 × 10−1 | 9.98 × 10−1 | −0.02 | 0.0623 | 7.49 × 10−1 | 9.41 × 10−1 | 0.06 | 0.109 | 5.99 × 10−1 | 9.54 × 10−1 | −0.02 | 0.0667 | 7.18 × 10−1 | 9.90 × 10−1 |
| G0 total | 0.05 | 0.1027 | 6.09 × 10−1 | 9.41 × 10−1 | −0.01 | 0.0648 | 8.26 × 10−1 | 9.62 × 10−1 | 0.01 | 0.1107 | 9.42 × 10−1 | 9.90 × 10−1 | −0.02 | 0.068 | 7.84 × 10−1 | 9.90 × 10−1 |
| G1 total | −0.16 | 0.1154 | 1.73 × 10−1 | 6.76 × 10−1 | −0.01 | 0.0896 | 9.17 × 10−1 | 9.89 × 10−1 | −0.1 | 0.1256 | 4.23 × 10−1 | 8.74 × 10−1 | 0.03 | 0.0925 | 7.75 × 10−1 | 9.90 × 10−1 |
| G2 total | −0.03 | 0.1011 | 7.75 × 10−1 | 9.41 × 10−1 | 0 | 0.0621 | 9.79 × 10−1 | 9.98 × 10−1 | −0.02 | 0.1098 | 8.42 × 10−1 | 9.90 × 10−1 | 0.02 | 0.0644 | 8.11 × 10−1 | 9.90 × 10−1 |
| F total | 0.1 | 0.1061 | 3.40 × 10−1 | 8.41 × 10−1 | 0.03 | 0.0811 | 6.74 × 10−1 | 9.41 × 10−1 | 0.15 | 0.1133 | 1.85 × 10−1 | 5.85 × 10−1 | 0.04 | 0.0838 | 6.09 × 10−1 | 9.54 × 10−1 |
| B total | 0.02 | 0.1092 | 8.73 × 10−1 | 9.62 × 10−1 | 0.16 | 0.0621 | 8.96 × 10−3 | 2.45 × 10−1 | 0.07 | 0.1204 | 5.83 × 10−1 | 9.54 × 10−1 | 0.2 | 0.0653 | 2.41 × 10−3 | 8.79 × 10−2 |
| FGS/(F + FG + FGS) | 0.05 | 0.1084 | 6.75 × 10−1 | 9.41 × 10−1 | −0.03 | 0.0601 | 6.61 × 10−1 | 9.41 × 10−1 | 0.09 | 0.1182 | 4.56 × 10−1 | 8.74 × 10−1 | −0.03 | 0.0635 | 6.13 × 10−1 | 9.54 × 10−1 |
| FBS1/(FS1 + FBS1) | −0.08 | 0.1061 | 4.80 × 10−1 | 8.93 × 10−1 | 0.11 | 0.0566 | 6.24 × 10−2 | 6.09 × 10−1 | −0.02 | 0.1163 | 8.60 × 10−1 | 9.90 × 10−1 | 0.12 | 0.0592 | 4.66 × 10−2 | 4.61 × 10−1 |
| FBS1/FS1 | −0.08 | 0.1061 | 4.80 × 10−1 | 8.93 × 10−1 | 0.11 | 0.0566 | 6.24 × 10−2 | 6.09 × 10−1 | −0.02 | 0.1163 | 8.60 × 10−1 | 9.90 × 10−1 | 0.12 | 0.0592 | 4.66 × 10−2 | 4.61 × 10−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radovani, B.; Vučković, F.; Maggioni, A.P.; Ferrannini, E.; Lauc, G.; Gudelj, I. IgG N-Glycosylation Is Altered in Coronary Artery Disease. Biomolecules 2023, 13, 375. https://doi.org/10.3390/biom13020375
Radovani B, Vučković F, Maggioni AP, Ferrannini E, Lauc G, Gudelj I. IgG N-Glycosylation Is Altered in Coronary Artery Disease. Biomolecules. 2023; 13(2):375. https://doi.org/10.3390/biom13020375
Chicago/Turabian StyleRadovani, Barbara, Frano Vučković, Aldo P. Maggioni, Ele Ferrannini, Gordan Lauc, and Ivan Gudelj. 2023. "IgG N-Glycosylation Is Altered in Coronary Artery Disease" Biomolecules 13, no. 2: 375. https://doi.org/10.3390/biom13020375
APA StyleRadovani, B., Vučković, F., Maggioni, A. P., Ferrannini, E., Lauc, G., & Gudelj, I. (2023). IgG N-Glycosylation Is Altered in Coronary Artery Disease. Biomolecules, 13(2), 375. https://doi.org/10.3390/biom13020375