
Untargeted Lipidomic Approach for Studying Different Nervous System Tissues of the Murine Model of Krabbe Disease

 ,
,  , ,
, ,  ,
,  , , ,
, , , 
 , ,
, ,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Preparation and Storage
2.3. Chemicals and Reagents
2.4. Sample Preparation for Lipidomics
2.5. LC/MS Analysis
2.6. Raw Data Processing
3. Results and Discussion
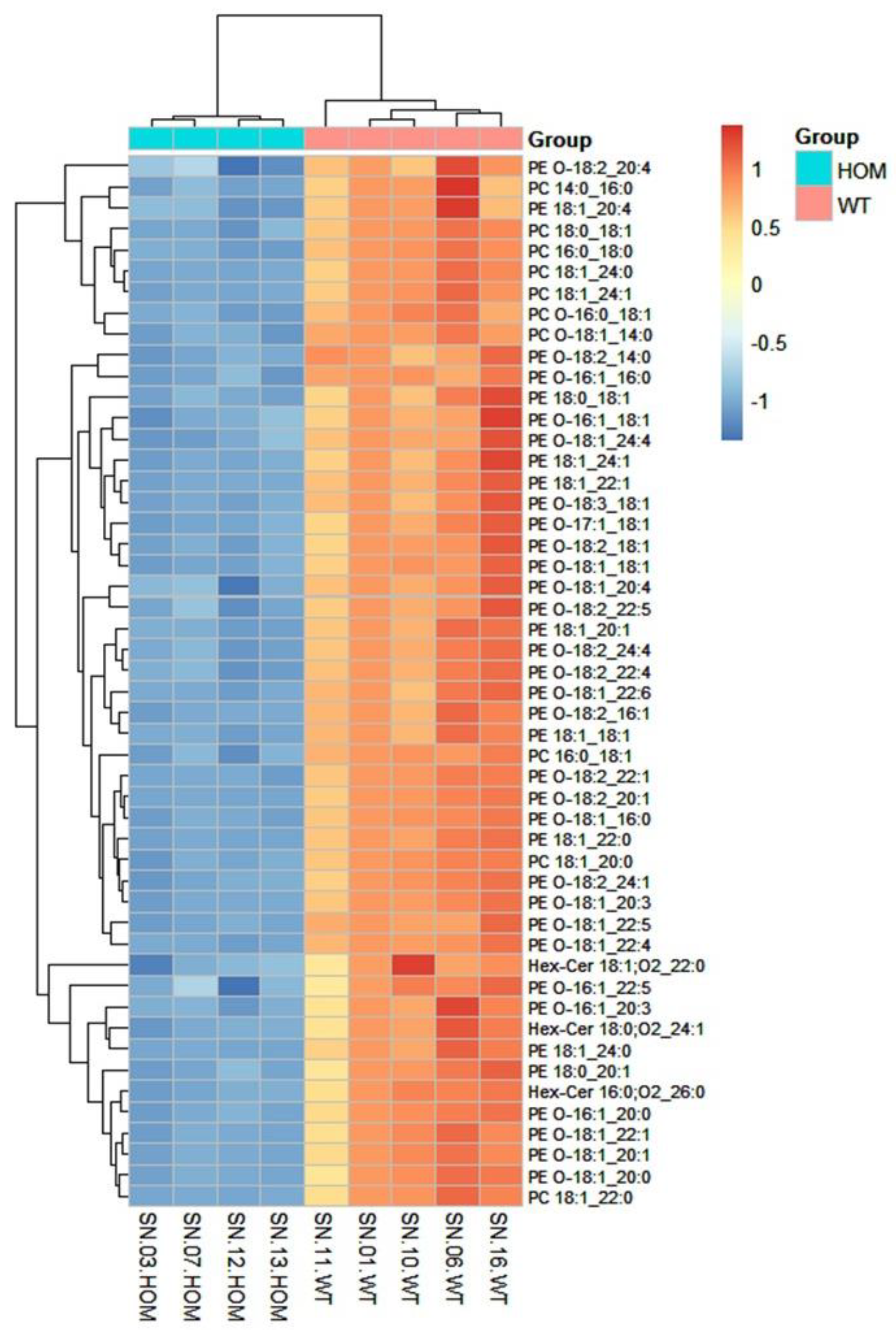
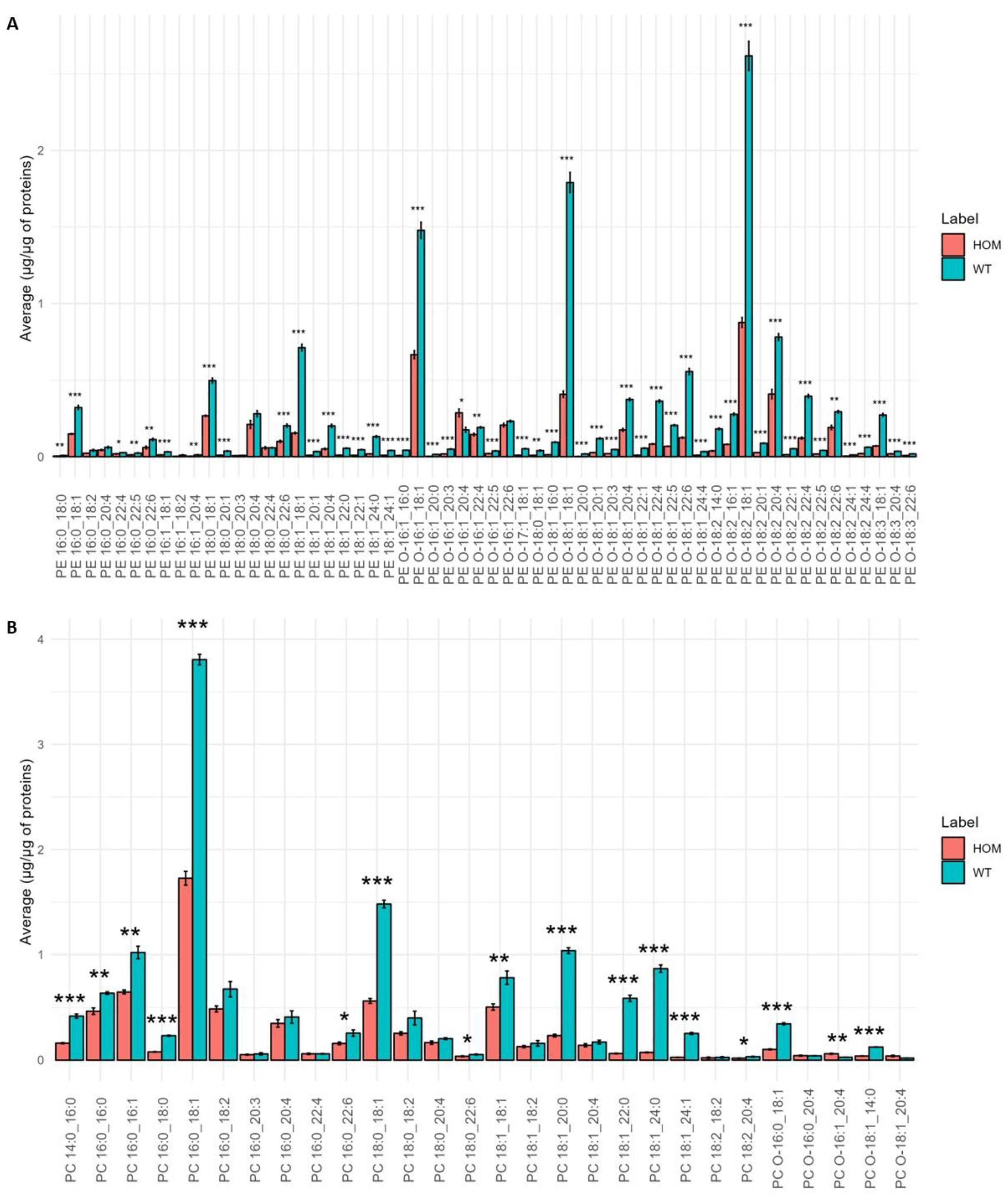
3.1. Within-Class Comparison of Lipid Profiles of Diseased and Healthy Mouse Models
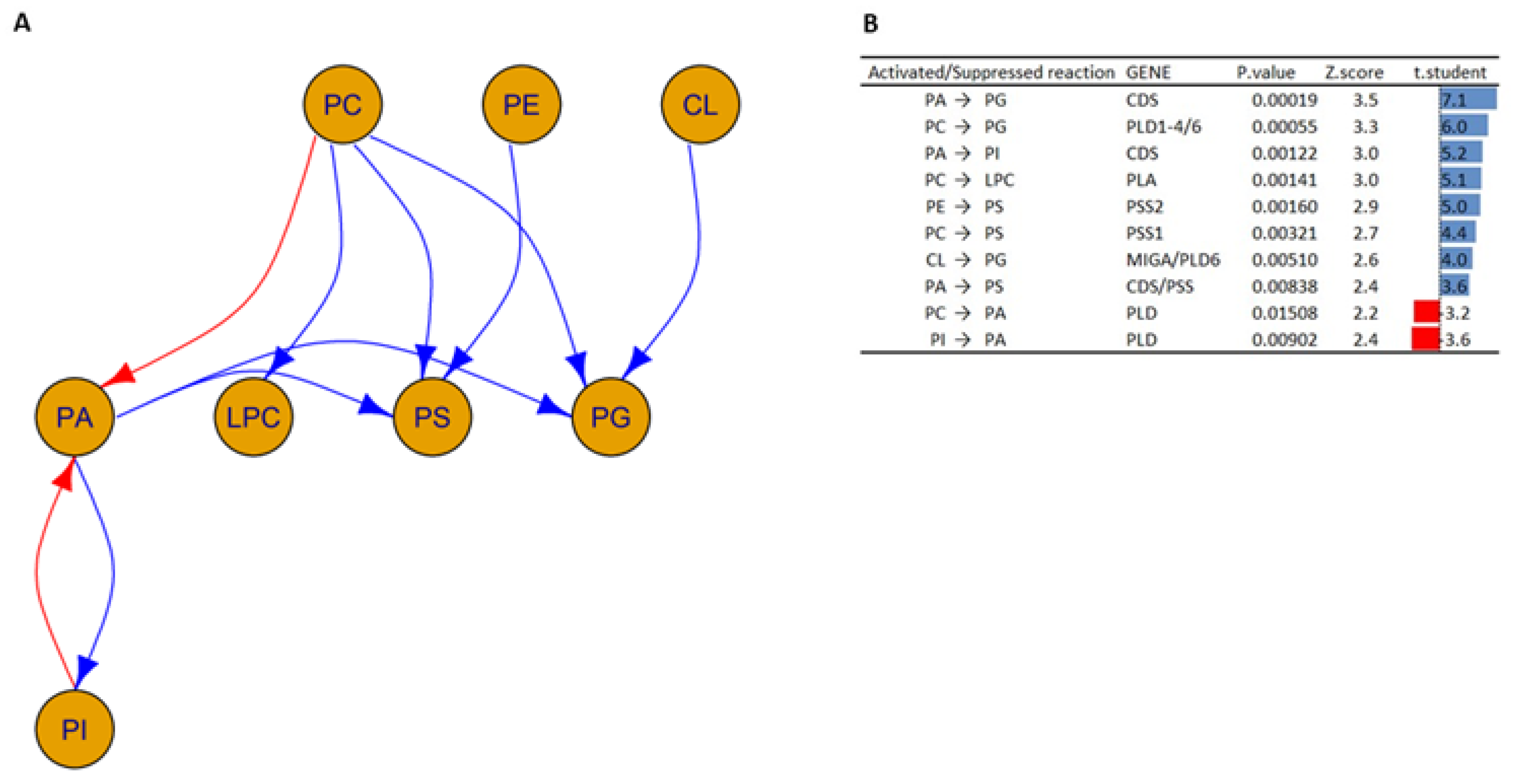
3.2. Interactions between Lipid Classes
- Increased activity of the enzyme phosphatidate cytidylyltransferase (also known as CDP-diacylglycerol synthase), which is involved in the transformation of PA to PG and PA to PI [39].
- Decreased enzyme activity of phospholipases D (encoded by PLD), which is responsible for the transformation of PI into PA and PC into PA [40].
- Increased activation in the phosphatidylserine synthase 1 and 2 enzymes (encoded by PSS1 and PSS2, respectively), which are responsible for the transformation of PC and PE into PS [41]. In addition, the concomitant activation of the PSS together with the CDS results in the transformation of PA into PS.
- Increased activation of the enzymes phospholipase D1 (encoded by PLD1), 5′-3′ exonuclease PLD4 (encoded by PLD4), and mitochondrial cardiolipin hydrolase (encoded by PLD6), which underlies PG synthesis from PCs [40].
- Increased activation of the enzymes mitoguardin 1 (encoded by MIGA) and mitochondrial cardiolipin hydrolase (encoded by PLD6), which are involved in the transformation of CL into PG.
- Increased activity of the phospholipase A enzyme (encoded by PLA), which is responsible for the conversion of PCs into LPCs [40]. This is in agreement with what has been reported in the literature; in fact, a similar trend has also been found in oligodendrocytes and astrocytes, where psychosine influences the regulation of cell death through the formation of LPCs [42,43].
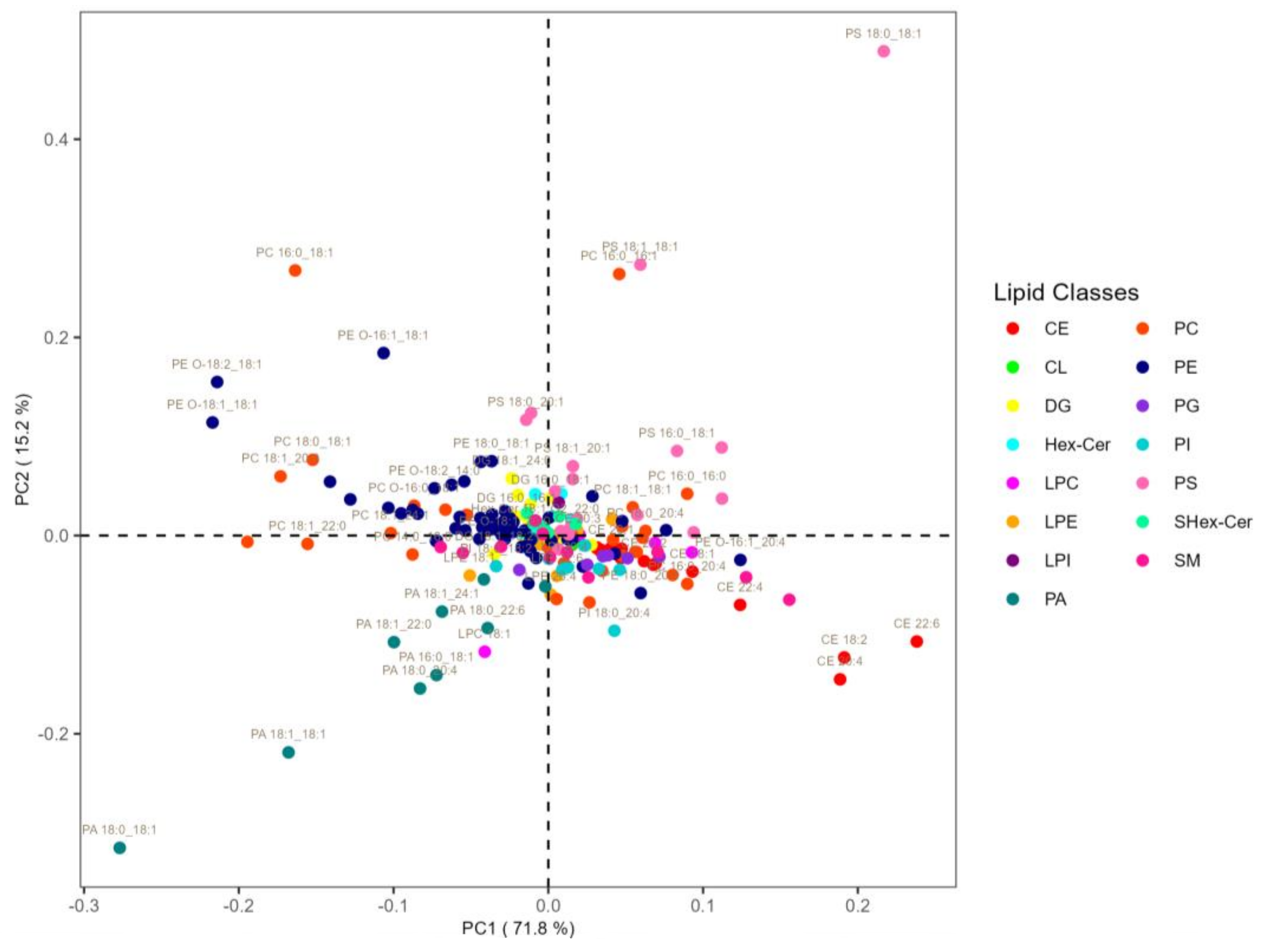
3.3. Analysis at the Level of Lipid Molecular Species
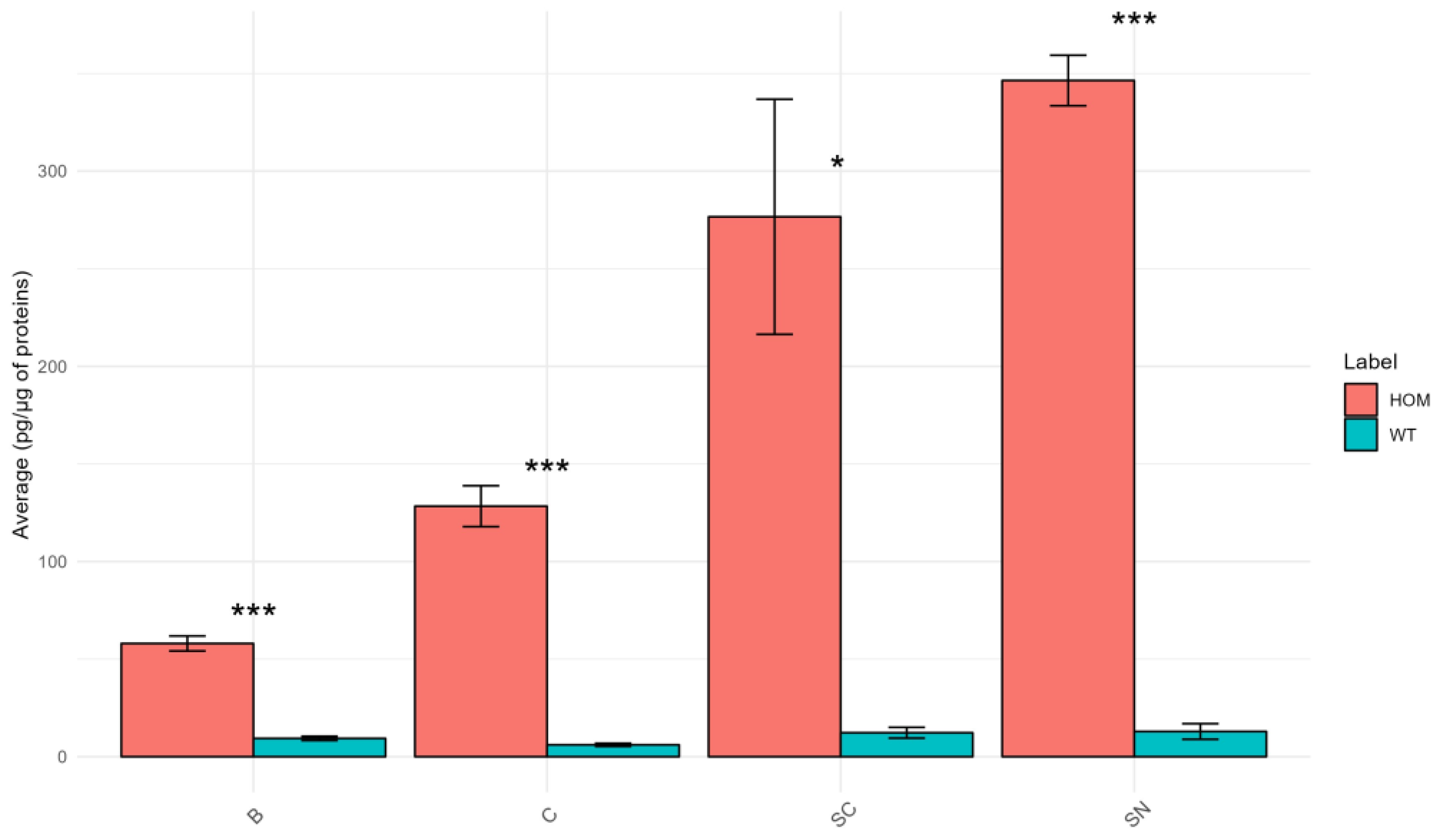
Psychosine Concentration
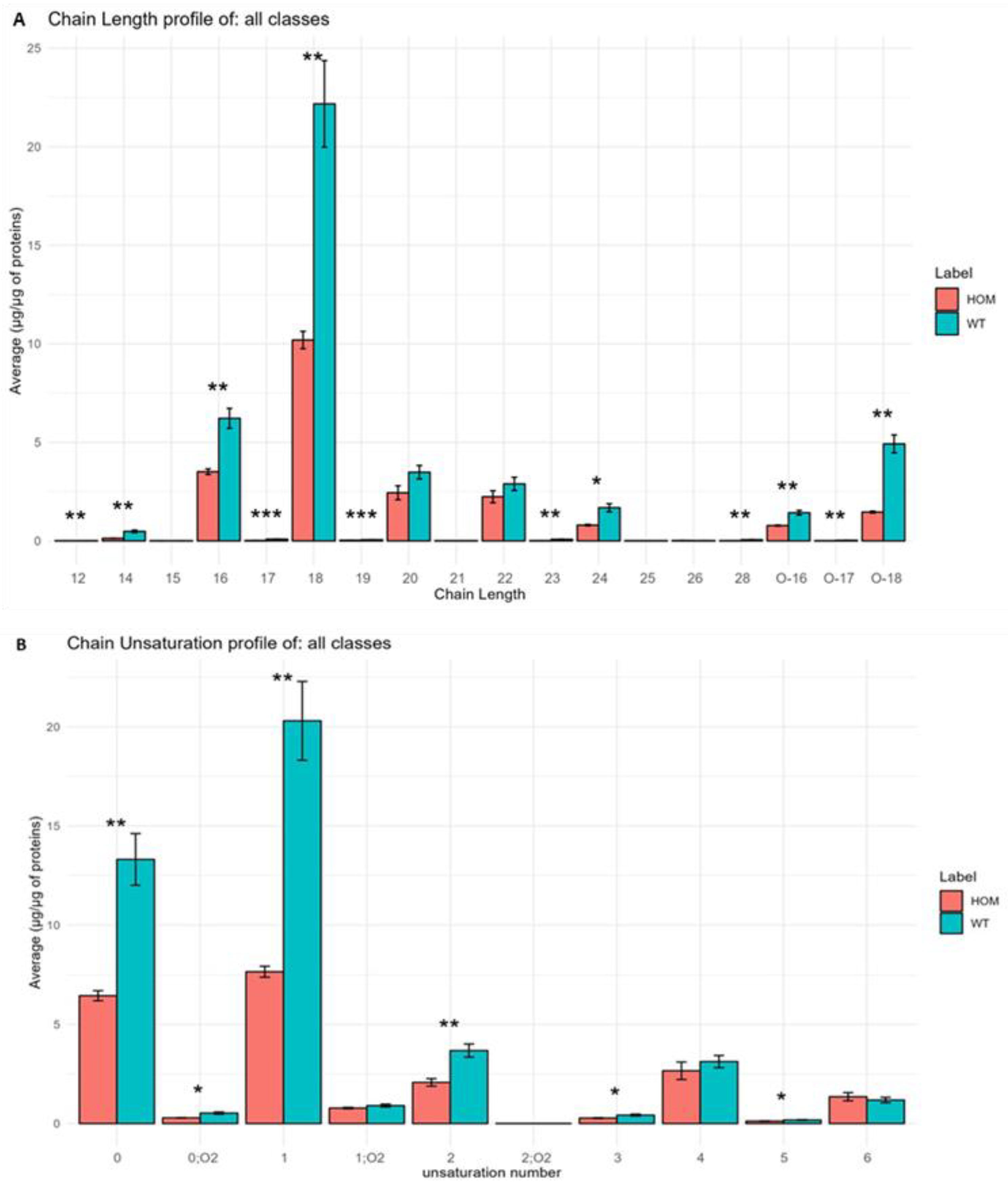
3.4. Analysis of Lipid Building Blocks
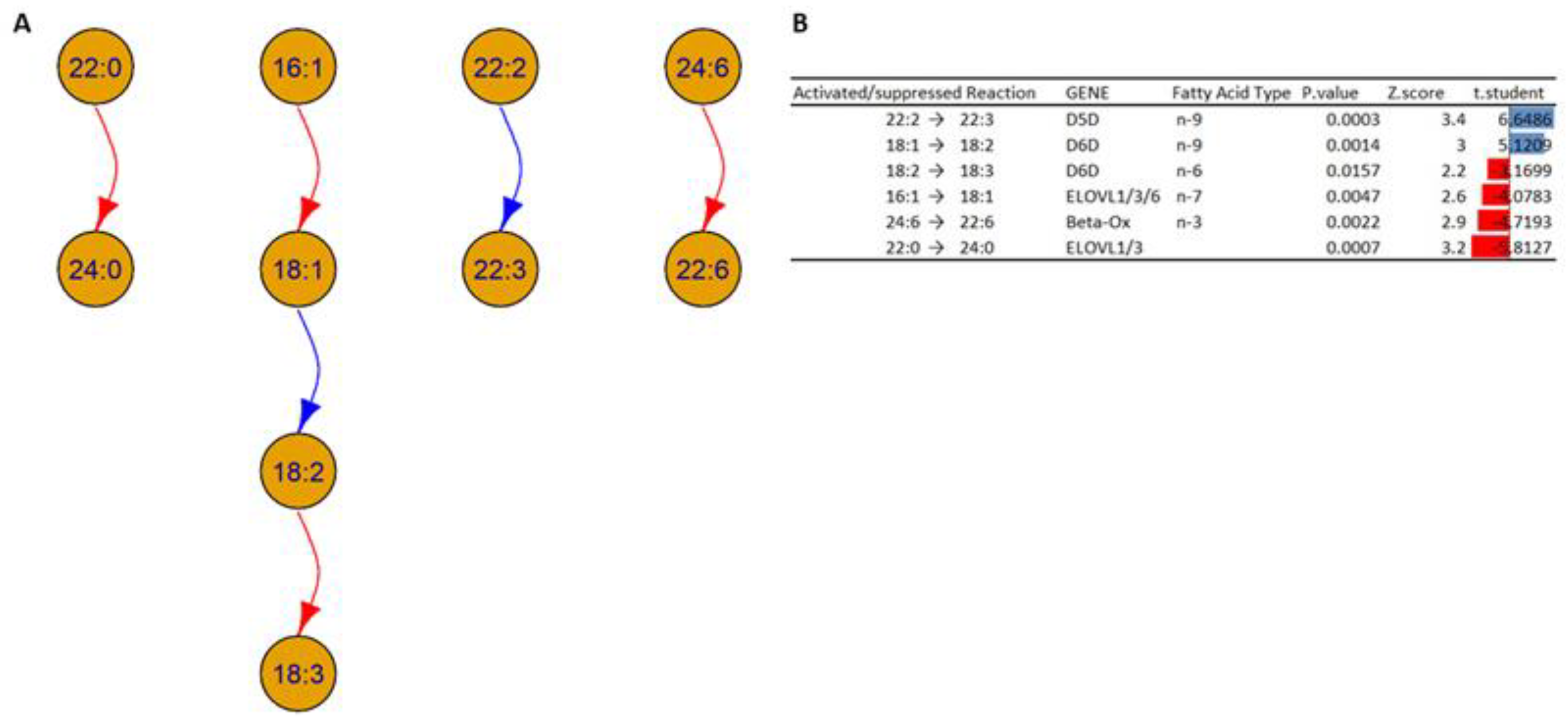
- Desaturation of the fatty acids: Based on the analysis, it is possible to note an increase in the concentrations of the chain 18:2 in HOMs compared to WT; this could be due to an alteration in the activity of Δ6 desaturase (D6D), which could indicate an increase in the transformation of fatty acid 18:1 (18 carbon atoms with one unsaturation) to fatty acid 18:2, and/or a decrease in the transformation of fatty acid 18:2 to 18:3. In addition, the analysis shows an increase in fatty acid 22:3 in HOM mice, which could indicate an increased desaturation of fatty acid 22:2, which could indicate an alteration in the activity of Δ5 desaturase (D5D).
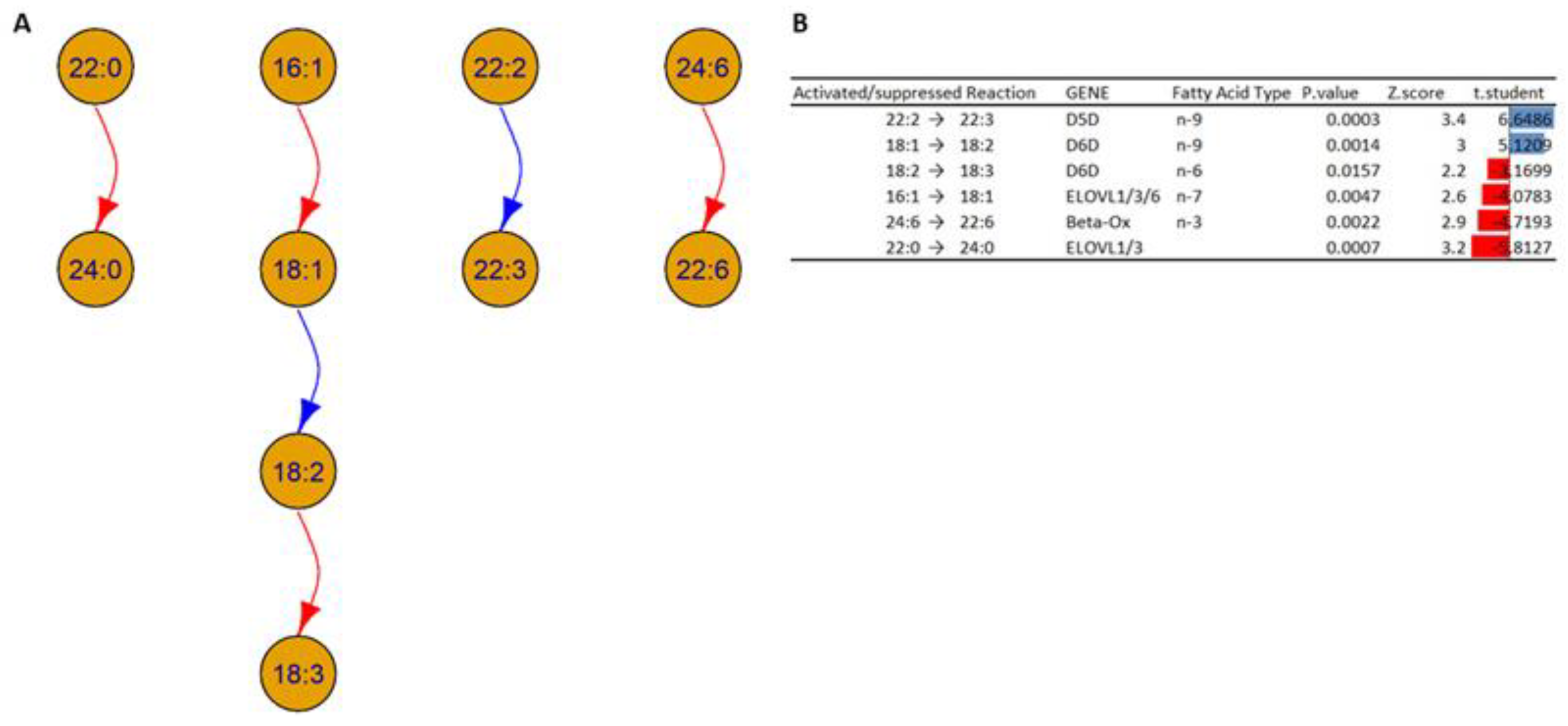
- Elongations and beta oxidation of the fatty acids: The analysis suggests a decreased activation in the activity of specific elongases (ELOVL) such as ELOVL1, ELOVL3, and ELOVL6, which can explain the decrease in the transformation of fatty acids 16:1 to 18:1 and 22:0 to 24:0 inside HOMs compared to WTs. The analysis also suggests decreased activation of the beta oxidation of fatty acid 24:6 in the HOMs.
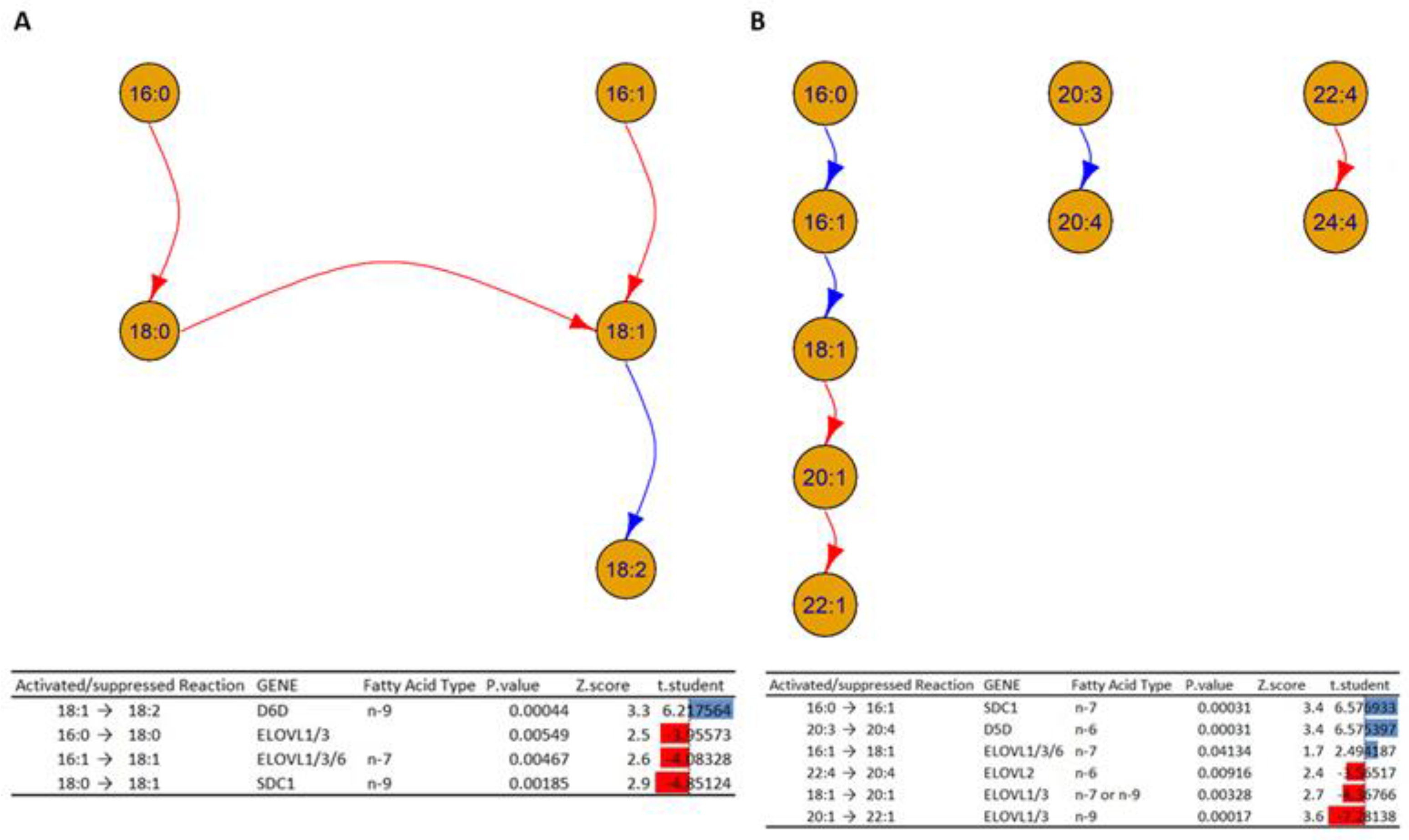
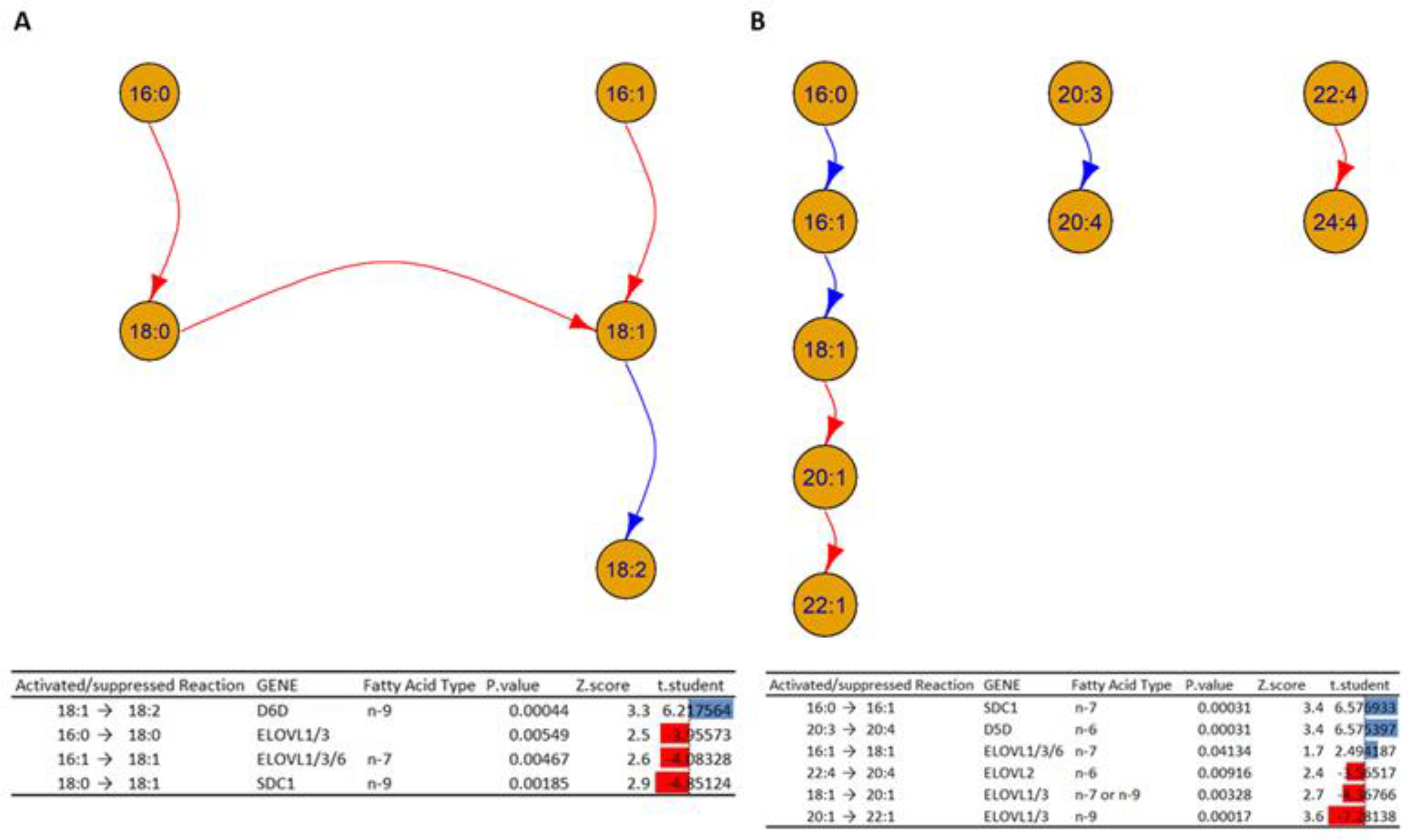
- The analysis in Figure 10A shows a decrease in activity in the elongation of the fatty acids 16:0 to 18:0 and 16:1 to 18:1, which could indicate alterations in the enzymes ELOVL1, ELOVL3, and ELOVL6 inside HOMs. On the other hand, the desaturation of 18:1 to 18:2 is increased in HOMs, which could indicate an increased activity of D6D. In addition, the analysis suggests an alteration in the activity of the enzyme Stearoyl-CoA desaturase (SCD1), which can explain the decrease transformation of the fatty acid 18:0 to 18:1 in the HOMs.
- Figure 10B shows a cascade of reactions that favor the formation of fatty acid 18:1 from the unsaturation of fatty acid 16:0, which could indicate an increased activity of SDC1 and a subsequent elongation to fatty acid 18:0, mediated by the ELOVL 1/3/6 genes. At the same time, the elongation of fatty acids 18:1 to 20:1 and 20:1 to 22:1 is inhibited, which could suggest alterations in the enzymes ELOVL 1/3. In addition, the desaturation of fatty acids 20:3 to 20:4 could suggest increased activation.
4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Del Grosso, A.; Parlanti, G.; Angella, L.; Giordano, N.; Tonazzini, I.; Ottalagana, E.; Carpi, S.; Pellegrino, R.M.; Alabed, H.B.R.; Emiliani, C.; et al. Chronic Lithium Administration in a Mouse Model for Krabbe Disease. JIMD Rep. 2022, 63, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Iacono, S.; Del Giudice, E.; Leon, A.; La Bella, V.; Spataro, R. A Novel Compound Heterozygous Mutation in GALC Associated with Adult-Onset Krabbe Disease: Case Report and Literature Review. Neurogenetics 2022, 23, 157–165. [Google Scholar] [CrossRef]
- White, A.B.; Galbiati, F.; Givogri, M.I.; Lopez Rosas, A.; Qiu, X.; van Breemen, R.; Bongarzone, E.R. Persistence of Psychosine in Brain Lipid Rafts Is a Limiting Factor in the Therapeutic Recovery of a Mouse Model for Krabbe Disease. J. Neurosci. Res. 2011, 89, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Mezzena, R.; Del Grosso, A.; Pellegrino, R.M.; Alabed, H.B.R.; Emiliani, C.; Tonazzini, I.; Cecchini, M. Mechanotransduction Impairment in Primary Fibroblast Model of Krabbe Disease. Biomedicines 2023, 11, 927. [Google Scholar] [CrossRef]
- Pellegrino, R.M.; Di Veroli, A.; Valeri, A.; Goracci, L.; Cruciani, G. LC/MS Lipid Profiling from Human Serum: A New Method for Global Lipid Extraction. Anal. Bioanal. Chem. 2014, 406, 7937–7948. [Google Scholar] [CrossRef] [PubMed]
- Géhin, C.; Fowler, S.J.; Trivedi, D.K. Chewing the Fat: How Lipidomics Is Changing Our Understanding of Human Health and Disease in 2022. Anal. Sci. Adv. 2023, 4, 104–131. [Google Scholar] [CrossRef]
- Han, X.; Gross, R.W. Global Analyses of Cellular Lipidomes Directly from Crude Extracts of Biological Samples by ESI Mass Spectrometry: A Bridge to Lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef]
- Pellegrino, R.M.; Giulietti, M.; Alabed, H.B.R.; Buratta, S.; Urbanelli, L.; Piva, F.; Emiliani, C. LipidOne: User-Friendly Lipidomic Data Analysis Tool for a Deeper Interpretation in a Systems Biology Scenario. Bioinformatics 2022, 38, 1767–1769. [Google Scholar] [CrossRef]
- Gaud, C.; Sousa, B.C.; Nguyen, A.; Fedorova, M.; Ni, Z.; O’Donnell, V.B.; Wakelam, M.J.O.; Andrews, S.; Lopez-Clavijo, A.F. BioPAN: A Web-Based Tool to Explore Mammalian Lipidome Metabolic Pathways on LIPID MAPS. F1000Research 2021, 10, 4. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Zhou, G.; Pang, Z.; Lu, Y.; Ewald, J.; Xia, J. OmicsNet 2.0: A Web-Based Platform for Multi-Omics Integration and Network Visual Analytics. Nucleic Acids Res. 2022, 50, W527–W533. [Google Scholar] [CrossRef] [PubMed]
- Alabed, H.B.R.; Gorello, P.; Pellegrino, R.M.; Lancioni, H.; La Starza, R.; Taddei, A.A.; Urbanelli, L.; Buratta, S.; Fernandez, A.G.L.; Matteucci, C.; et al. Comparison between Sickle Cell Disease Patients and Healthy Donors: Untargeted Lipidomic Study of Erythrocytes. Int. J. Mol. Sci. 2023, 24, 2529. [Google Scholar] [CrossRef] [PubMed]
- Abinaya, B.; Waseem, M.; Kashif, M.; Srinivasan, H. Lipidomics: An Excellent Tool for Chronic Disease Detection. Curr. Res. Transl. Med. 2022, 70, 103346. [Google Scholar] [CrossRef]
- Fernandez, C.; Sandin, M.; Sampaio, J.L.; Almgren, P.; Narkiewicz, K.; Hoffmann, M.; Hedner, T.; Wahlstrand, B.; Simons, K.; Shevchenko, A.; et al. Plasma Lipid Composition and Risk of Developing Cardiovascular Disease. PLoS ONE 2013, 8, e71846. [Google Scholar] [CrossRef] [PubMed]
- Graessler, J.; Schwudke, D.; Schwarz, P.E.H.; Herzog, R.; Shevchenko, A.; Bornstein, S.R. Top-Down Lipidomics Reveals Ether Lipid Deficiency in Blood Plasma of Hypertensive Patients. PLoS ONE 2009, 4, e6261. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Holtzman, D.M.; McKeel, D.W.; Kelley, J.; Morris, J.C. Substantial Sulfatide Deficiency and Ceramide Elevation in Very Early Alzheimer’s Disease: Potential Role in Disease Pathogenesis. J. Neurochem. 2002, 82, 809–818. [Google Scholar] [CrossRef]
- Mills, G.B.; Moolenaar, W.H. The Emerging Role of Lysophosphatidic Acid in Cancer. Nat. Rev. Cancer 2003, 3, 582–591. [Google Scholar] [CrossRef]
- Won, J.-S.; Kim, J.; Paintlia, M.K.; Singh, I.; Singh, A.K. Role of Endogenous Psychosine Accumulation in Oligodendrocyte Differentiation and Survival: Implication for Krabbe Disease. Brain Res. 2013, 1508, 44–52. [Google Scholar] [CrossRef]
- Guenzel, A.J.; Turgeon, C.T.; Nickander, K.K.; White, A.L.; Peck, D.S.; Pino, G.B.; Studinski, A.L.; Prasad, V.K.; Kurtzberg, J.; Escolar, M.L.; et al. The Critical Role of Psychosine in Screening, Diagnosis, and Monitoring of Krabbe Disease. Genet. Med. 2020, 22, 1108–1118. [Google Scholar] [CrossRef]
- Reiter, C.R.; Rebiai, R.; Kwak, A.; Marshall, J.; Wozniak, D.; Scesa, G.; Nguyen, D.; Rue, E.; Pathmasiri, K.C.; Pijewski, R.; et al. The Pathogenic Sphingolipid Psychosine Is Secreted in Extracellular Vesicles in the Brain of a Mouse Model of Krabbe Disease. ASN Neuro 2022, 14, 175909142210878. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Y.; Benitez, B.A.; Nagree, M.S.; Dearborn, J.T.; Jiang, X.; Guzman, M.A.; Woloszynek, J.C.; Giaramita, A.; Yip, B.K.; et al. Genetic Ablation of Acid Ceramidase in Krabbe Disease Confirms the Psychosine Hypothesis and Identifies a New Therapeutic Target. Proc. Natl. Acad. Sci. USA 2019, 116, 20097–20103. [Google Scholar] [CrossRef] [PubMed]
- Spassieva, S.; Bieberich, E. Lysosphingolipids and Sphingolipidoses: Psychosine in Krabbe’s Disease. J. Neurosci. Res. 2016, 94, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Carpi, S.; Del Grosso, A.; De Sarlo, M.; Colagiorgio, L.; Scaccini, L.; Tonazzini, I.; Parlanti, G.; Cecchini, M. Reliable and Fast Genotyping Protocol for Galactosylceramidase (Galc) in the Twitcher (Twi) Mouse. Biomedicines 2022, 10, 3146. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Suzuki, K. The Twitcher Mouse. A Model of Human Globoid Cell Leukodystrophy (Krabbe’s Disease). Am. J. Pathol. 1983, 111, 394–397. [Google Scholar]
- Tsugawa, H.; Ikeda, K.; Takahashi, M.; Satoh, A.; Mori, Y.; Uchino, H.; Okahashi, N.; Yamada, Y.; Tada, I.; Bonini, P.; et al. A Lipidome Atlas in MS-DIAL 4. Nat. Biotechnol. 2020, 38, 1159–1163. [Google Scholar] [CrossRef]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Köfeler, H.; et al. Update on LIPID MAPS Classification, Nomenclature, and Shorthand Notation for MS-Derived Lipid Structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef]
- Aschner, M.; Toews, A.D. Myelin and Myelination. In Comprehensive Toxicology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 181–198. [Google Scholar]
- Poitelon, Y.; Kopec, A.M.; Belin, S. Myelin Fat Facts: An Overview of Lipids and Fatty Acid Metabolism. Cells 2020, 9, 812. [Google Scholar] [CrossRef]
- Dupree, J.L.; Pomicter, A.D. Myelin, DIGs, and Membrane Rafts in the Central Nervous System. Prostaglandins Other Lipid Mediat. 2010, 91, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, Y.; Todo, M.; Bizen, H.; Kimura, D.; Suzuki, T. Precision Pinch Force Control via Brain and Spinal Motor Neuron Excitability during Motor Imagery. Neurosci. Lett. 2021, 754, 135843. [Google Scholar] [CrossRef]
- Harry, G.J.; Toews, A.D. Myelination, Dysmyelination, and Demyelination. In Handbook of Developmental Neurotoxicology; Elsevier: Amsterdam, The Netherlands, 1998; pp. 87–115. [Google Scholar]
- Dong, Y.; Yong, V.W. Oxidized Phospholipids as Novel Mediators of Neurodegeneration. Trends Neurosci. 2022, 45, 419–429. [Google Scholar] [CrossRef]
- Marcus, J.; Popko, B. Galactolipids Are Molecular Determinants of Myelin Development and Axo–Glial Organization. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2002, 1573, 406–413. [Google Scholar] [CrossRef]
- Gómez-Fernández, J.C.; Corbalán-García, S. Diacylglycerols, Multivalent Membrane Modulators. Chem. Phys. Lipids 2007, 148, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Campomanes, P.; Zoni, V.; Vanni, S. Local Accumulation of Diacylglycerol Alters Membrane Properties Nonlinearly Due to Its Transbilayer Activity. Commun. Chem. 2019, 2, 72. [Google Scholar] [CrossRef]
- Dorninger, F.; Forss-Petter, S.; Berger, J. From Peroxisomal Disorders to Common Neurodegenerative Diseases—The Role of Ether Phospholipids in the Nervous System. FEBS Lett. 2017, 591, 2761–2788. [Google Scholar] [CrossRef]
- Phillips, G.R.; Hancock, S.E.; Brown, S.H.J.; Jenner, A.M.; Kreilaus, F.; Newell, K.A.; Mitchell, T.W. Cholesteryl Ester Levels Are Elevated in the Caudate and Putamen of Huntington’s Disease Patients. Sci. Rep. 2020, 10, 20314. [Google Scholar] [CrossRef]
- Nguyen, A.; Rudge, S.A.; Zhang, Q.; Wakelam, M.J. Using Lipidomics Analysis to Determine Signalling and Metabolic Changes in Cells. Curr. Opin. Biotechnol. 2017, 43, 96–103. [Google Scholar] [CrossRef]
- Mishra, N.N.; Tran, T.T.; Seepersaud, R.; Garcia-de-la-Maria, C.; Faull, K.; Yoon, A.; Proctor, R.; Miro, J.M.; Rybak, M.J.; Bayer, A.S.; et al. Perturbations of Phosphatidate Cytidylyltransferase (CdsA) Mediate Daptomycin Resistance in Streptococcus Mitis/Oralis by a Novel Mechanism. Antimicrob. Agents Chemother. 2017, 61, e02435-16. [Google Scholar] [CrossRef]
- Fisher, A.B.; Jain, M. Phospholipases: Degradation of Membrane Phospholipids. In eLS; Wiley: Hoboken, NJ, USA, 2001. [Google Scholar]
- Kuge, O.; Nishijima, M. Phosphatidylserine Synthase I and II of Mammalian Cells. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1997, 1348, 151–156. [Google Scholar] [CrossRef]
- Giri, S.; Khan, M.; Rattan, R.; Singh, I.; Singh, A.K. Krabbe Disease: Psychosine-Mediated Activation of Phospholipase A2 in Oligodendrocyte Cell Death. J. Lipid Res. 2006, 47, 1478–1492. [Google Scholar] [CrossRef]
- Misslin, C.; Velasco-Estevez, M.; Albert, M.; O’Sullivan, S.A.; Dev, K.K. Phospholipase A2 Is Involved in Galactosylsphingosine-Induced Astrocyte Toxicity, Neuronal Damage and Demyelination. PLoS ONE 2017, 12, e0187217. [Google Scholar] [CrossRef]
- Chrast, R.; Saher, G.; Nave, K.-A.; Verheijen, M.H.G. Lipid Metabolism in Myelinating Glial Cells: Lessons from Human Inherited Disorders and Mouse Models. J. Lipid Res. 2011, 52, 419–434. [Google Scholar] [CrossRef]
- Olmstead, C.E. Neurological and Neurobehavioral Development of the Mutant ‘Twitcher’ Mouse. Behav. Brain Res. 1987, 25, 143–153. [Google Scholar] [CrossRef]
- Ichioka, T.; Kishimoto, Y.; Brennan, S.; Santos, G.W.; Yeager, A.M. Hematopoietic Cell Transplantation in Murine Globoid Cell Leukodystrophy (the Twitcher Mouse): Effects on Levels of Galactosylceramidase, Psychosine, and Galactocerebrosides. Proc. Natl. Acad. Sci. USA 1987, 84, 4259–4263. [Google Scholar] [CrossRef] [PubMed]
- Ricca, A.; Rufo, N.; Ungari, S.; Morena, F.; Martino, S.; Kulik, W.; Alberizzi, V.; Bolino, A.; Bianchi, F.; Del Carro, U.; et al. Combined Gene/Cell Therapies Provide Long-Term and Pervasive Rescue of Multiple Pathological Symptoms in a Murine Model of Globoid Cell Leukodystrophy. Hum. Mol. Genet. 2015, 24, 3372–3389. [Google Scholar] [CrossRef]
- D’Auria, L.; Reiter, C.; Ward, E.; Moyano, A.L.; Marshall, M.S.; Nguyen, D.; Scesa, G.; Hauck, Z.; van Breemen, R.; Givogri, M.I.; et al. Psychosine Enhances the Shedding of Membrane Microvesicles: Implications in Demyelination in Krabbe’s Disease. PLoS ONE 2017, 12, e0178103. [Google Scholar] [CrossRef] [PubMed]
- Fernández, R.; Carriel, V.; Lage, S.; Garate, J.; Díez-García, J.; Ochoa, B.; Castro, B.; Alaminos, M.; Fernández, J.A. Deciphering the Lipid Architecture of the Rat Sciatic Nerve Using Imaging Mass Spectrometry. ACS Chem. Neurosci. 2016, 7, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Heller, G.J.; Marshall, M.S.; Issa, Y.; Marshall, J.N.; Nguyen, D.; Rue, E.; Pathmasiri, K.C.; Domowicz, M.S.; van Breemen, R.B.; Tai, L.M.; et al. Waning Efficacy in a Long-Term AAV-Mediated Gene Therapy Study in the Murine Model of Krabbe Disease. Mol. Ther. 2021, 29, 1883–1902. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B | C | SC | SN | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Explained Class Name | WT | HOM | p-Value | WT | HOM | p-Value | WT | HOM | p-Value | WT | HOM | p-Value |
| Cholesteryl ester | 0.087 | 0.115 | 0.131 | 0.182 | 0.137 | 0.730 | * | 0.169 | 2.351 | ** | ||
| Ceramide | 0.022 | 0.023 | 0.055 | 0.050 | 0.063 | 0.054 | 0.011 | 0.012 | ||||
| Cardiolipin | 0.330 | 0.327 | 0.182 | 0.203 | 0.143 | 0.217 | 0.142 | 0.246 | ||||
| Diacylglycerol | 0.115 | 0.101 | 0.210 | 0.137 | 0.445 | 0.251 | * | 0.466 | 0.174 | *** | ||
| Ether-linked phosphatidylcholine | 0.077 | 0.076 | 0.908 | 0.863 | 0.245 | 0.218 | 0.613 | 0.287 | ** | |||
| Ether-linked phosphatidylethanolamine | 3.670 | 3.483 | 5.619 | 4.557 | * | 10.399 | 8.169 | * | 12.154 | 4.209 | *** | |
| Ether-linked phosphatidylinositol | 0.198 | 0.176 | ||||||||||
| Hexosylceramide | 0.029 | 0.023 | 0.040 | 0.028 | * | 0.151 | 0.128 | 0.193 | 0.102 | *** | ||
| Lysophosphatidylcholine | 0.231 | 0.239 | 0.251 | 0.262 | 0.598 | 0.505 | 0.480 | 0.401 | ||||
| Lysophosphatidylethanolamine | 0.406 | 0.409 | 0.380 | 0.405 | 1.065 | 0.853 | 0.424 | 0.232 | * | |||
| Lysophosphatidylinositol | 0.029 | 0.024 | 0.042 | 0.033 | 0.054 | 0.040 | * | 0.035 | 0.023 | * | ||
| Phosphatidic acid | 0.052 | 0.046 | 0.158 | 0.092 | 2.891 | 1.065 | * | 4.295 | 0.342 | * | ||
| Phosphatidylcholine | 8.254 | 7.902 | 8.312 | 7.994 | 9.829 | 8.323 | * | 14.912 | 6.426 | *** | ||
| Phosphatidylethanolamine | 2.612 | 2.590 | 3.254 | 2.822 | 4.621 | 3.724 | 3.277 | 1.245 | ** | |||
| Phosphatidylglycerol | 0.082 | 0.089 | 0.096 | 0.097 | 0.184 | 0.227 | 0.059 | 0.164 | ** | |||
| Phosphatidylinositol | 0.438 | 0.364 | 0.925 | 0.817 | 1.421 | 1.194 | 1.146 | 0.684 | * | |||
| Phosphatidylserine | 3.071 | 3.009 | 4.182 | 3.832 | 6.569 | 5.589 | 2.740 | 2.980 | ||||
| Sulfatide | 0.021 | 0.018 | * | 0.032 | 0.023 | *** | 0.058 | 0.044 | 0.070 | 0.037 | ** | |
| Sphingomyelin | 0.614 | 0.617 | 0.186 | 0.194 | 0.893 | 1.056 | 2.468 | 1.771 | ||||
| Total | 20.139 | 19.456 | 24.962 | 22.591 | 39.964 | 32.564 | 43.654 | 21.686 | ** | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alabed, H.B.R.; Del Grosso, A.; Bellani, V.; Urbanelli, L.; Carpi, S.; De Sarlo, M.; Bertocci, L.; Colagiorgio, L.; Buratta, S.; Scaccini, L.; et al. Untargeted Lipidomic Approach for Studying Different Nervous System Tissues of the Murine Model of Krabbe Disease. Biomolecules 2023, 13, 1562. https://doi.org/10.3390/biom13101562
Alabed HBR, Del Grosso A, Bellani V, Urbanelli L, Carpi S, De Sarlo M, Bertocci L, Colagiorgio L, Buratta S, Scaccini L, et al. Untargeted Lipidomic Approach for Studying Different Nervous System Tissues of the Murine Model of Krabbe Disease. Biomolecules. 2023; 13(10):1562. https://doi.org/10.3390/biom13101562
Chicago/Turabian StyleAlabed, Husam B. R., Ambra Del Grosso, Valeria Bellani, Lorena Urbanelli, Sara Carpi, Miriam De Sarlo, Lorenzo Bertocci, Laura Colagiorgio, Sandra Buratta, Luca Scaccini, and et al. 2023. "Untargeted Lipidomic Approach for Studying Different Nervous System Tissues of the Murine Model of Krabbe Disease" Biomolecules 13, no. 10: 1562. https://doi.org/10.3390/biom13101562
APA StyleAlabed, H. B. R., Del Grosso, A., Bellani, V., Urbanelli, L., Carpi, S., De Sarlo, M., Bertocci, L., Colagiorgio, L., Buratta, S., Scaccini, L., Frongia Mancini, D., Tonazzini, I., Cecchini, M., Emiliani, C., & Pellegrino, R. M. (2023). Untargeted Lipidomic Approach for Studying Different Nervous System Tissues of the Murine Model of Krabbe Disease. Biomolecules, 13(10), 1562. https://doi.org/10.3390/biom13101562