
Role of Cytokine-Inducible SH2 Domain-Containing (CISH) Protein in the Regulation of Erythropoiesis

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Mouse Studies
2.2. Embryo Analysis
2.3. Blood Analysis
2.4. Bone Marrow and Spleen Harvest
2.5. FACS
2.6. Colony-Forming Assays
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
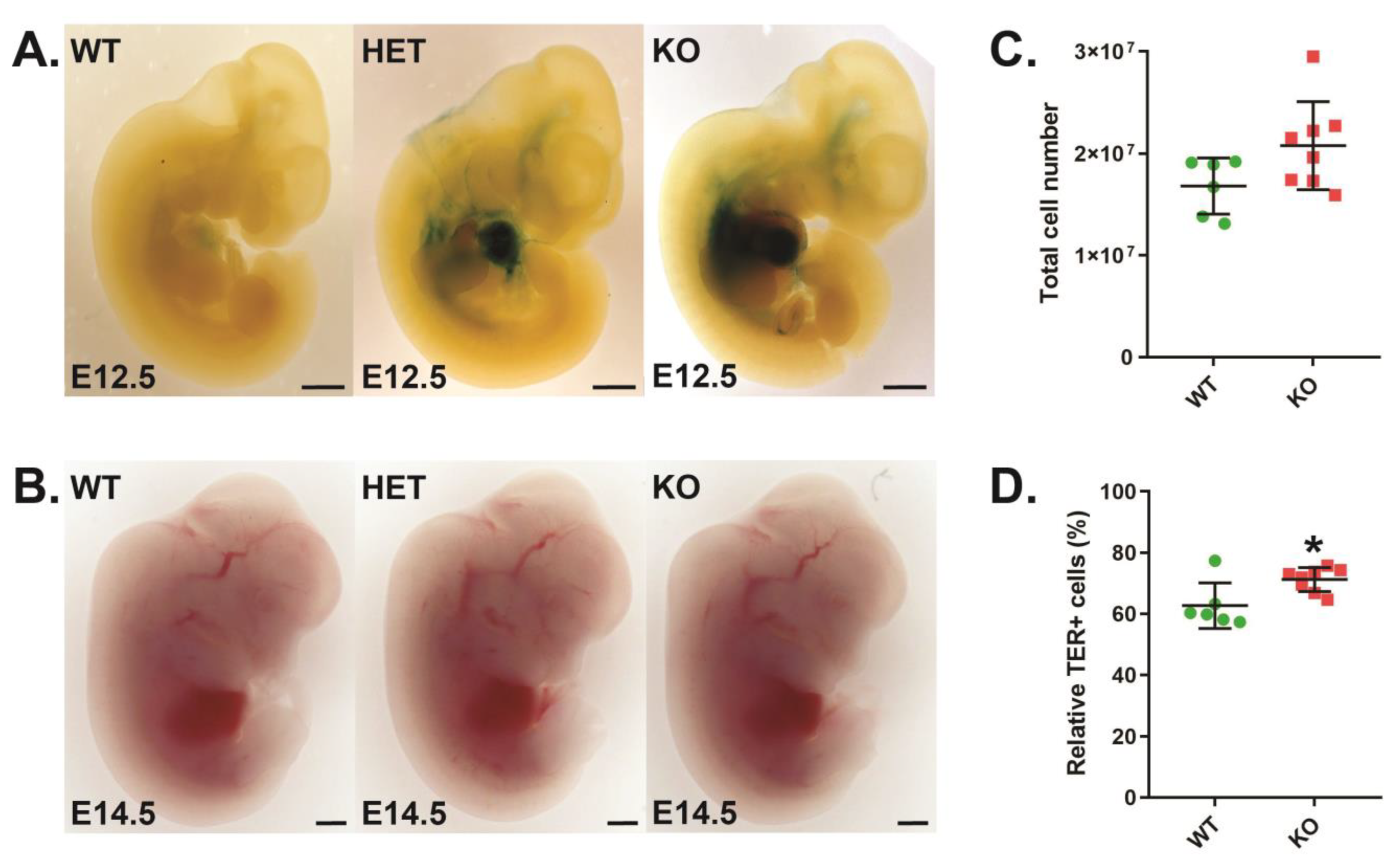
3.1. Role of CISH in Developmental Erythropoiesis
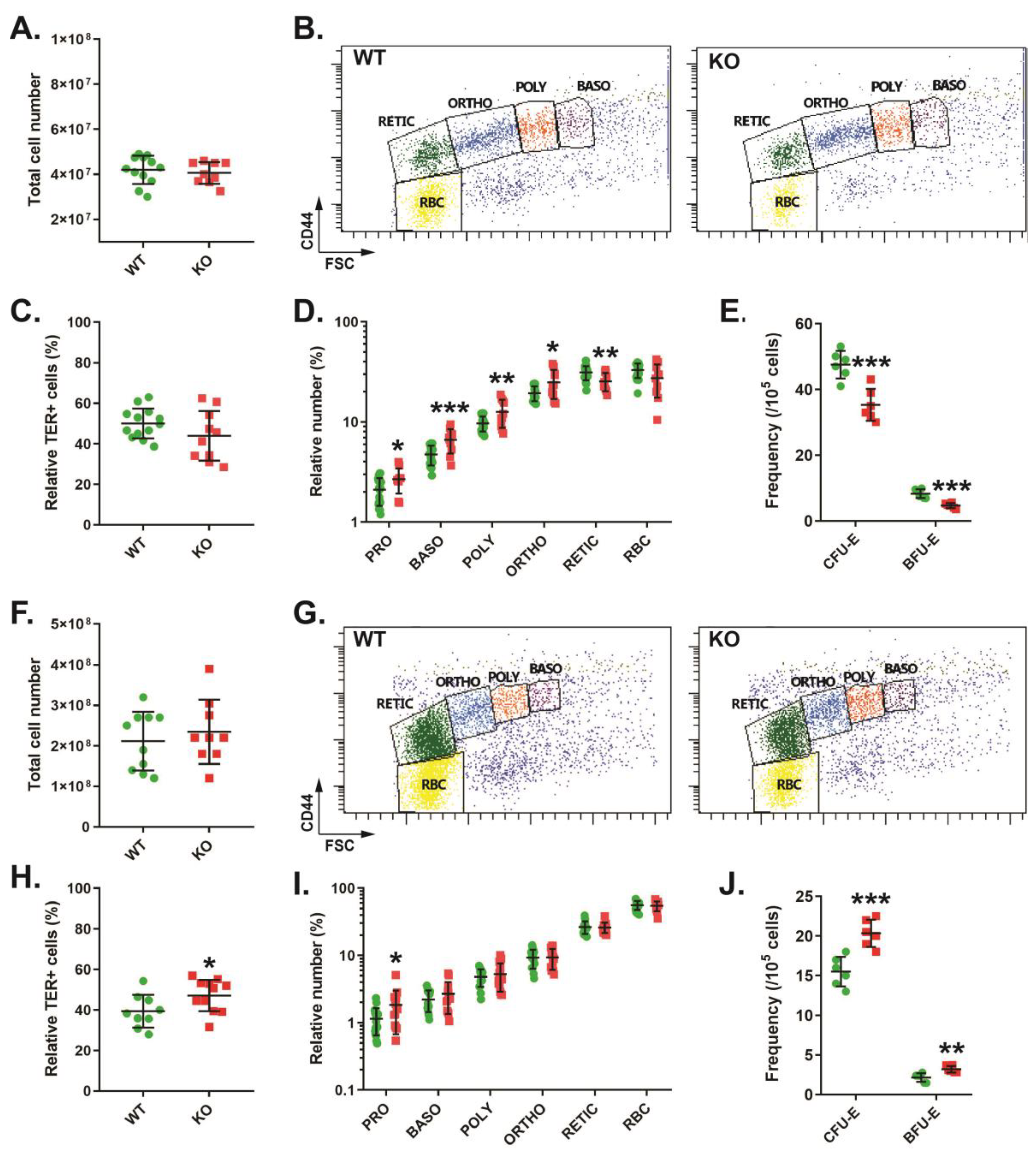
3.2. Role of CISH in Steady-State Erythropoiesis
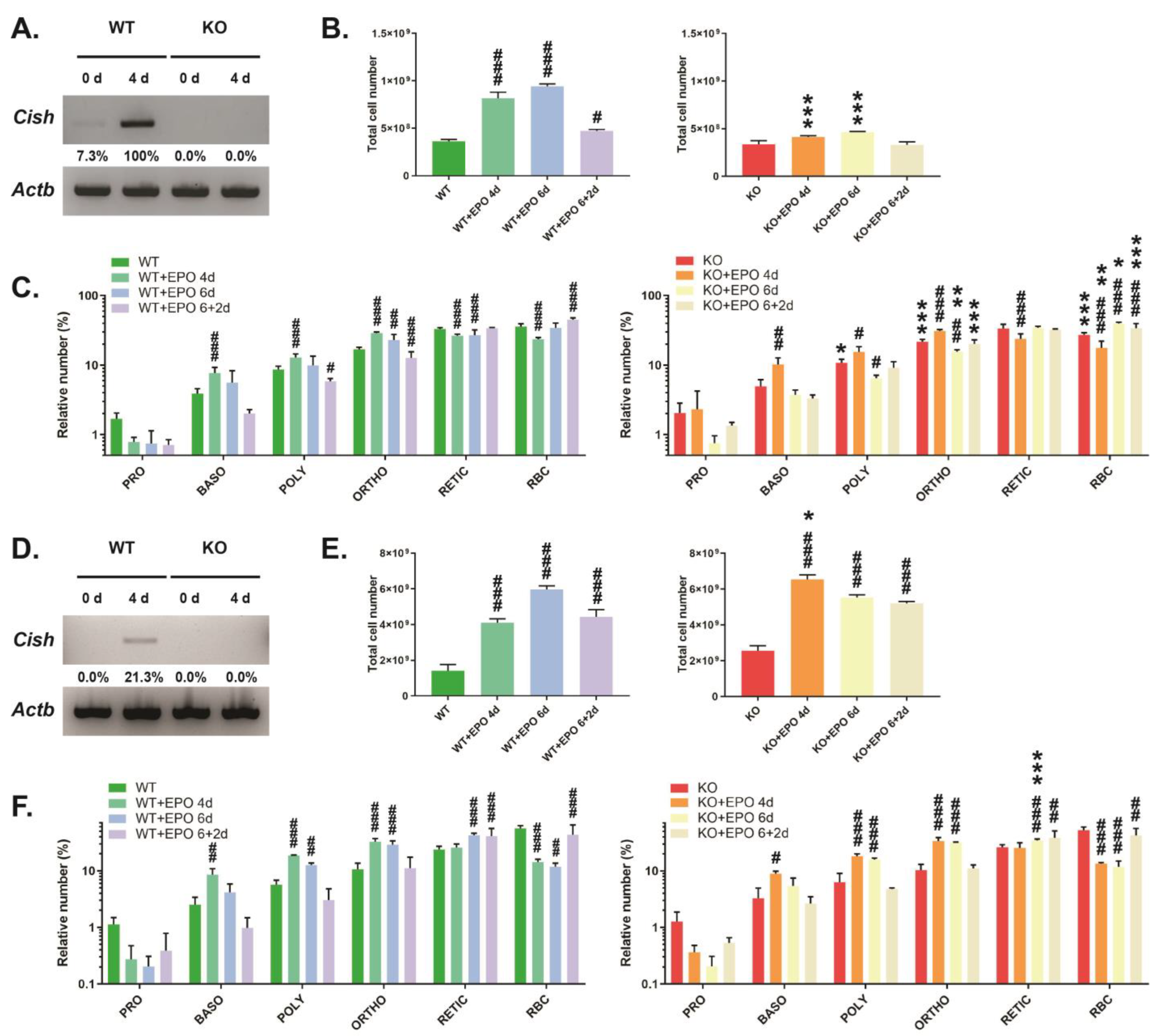
3.3. Role of CISH in EPO-Induced Erythropoiesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wojchowski, D.M.; Menon, M.P.; Sathyanarayana, P.; Fang, J.; Karur, V.; Houde, E.; Kapelle, W.; Bogachev, O. Erythropoietin-dependent erythropoiesis: New insights and questions. Blood Cells Mol. Dis. 2006, 36, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Kuhrt, D.; Wojchowski, D.M. Emerging EPO and EPO receptor regulators and signal transducers. Blood 2015, 125, 3536–3541. [Google Scholar] [CrossRef] [PubMed]
- Trengove, M.C.; Ward, A.C. SOCS proteins in development and disease. Am. J. Exp. Clin. Immunol. 2013, 2, 1–29. [Google Scholar]
- Yoshimura, A.; Ohkubo, T.; Kiguchi, T.; Jenkins, N.A.; Gilbert, D.J.; Copeland, N.G.; Hara, T.; Miyajima, A. A novel cytokine-inducible gene CIS encodes an SH2-containing protein that binds to tyrosine-phosphorylated interleukin 3 and erythropoietin receptors. EMBO J. 1995, 14, 2816–2826. [Google Scholar] [CrossRef] [PubMed]
- Sobah, M.L.; Liongue, C.; Ward, A.C. SOCS proteins in immunity, inflammatory diseases and immune-related cancer. Front. Med. 2021, 8, 727987. [Google Scholar] [CrossRef] [PubMed]
- Verdier, F.; Chretien, S.; Muller, O.; Varlet, P.; Yoshimura, A.; Gisselbrecht, S.; Lacombe, C.; Mayeux, P. Proteasomes regulate erythropoietin receptor and STAT5 activation: Possible involvement of the ubiquitinated CIS protein. J. Biol. Chem. 1998, 273, 28185–28190. [Google Scholar] [CrossRef] [PubMed]
- Piessevaux, J.; De Ceuninck, L.; Catteeuw, D.; Peelman, F.; Tavernier, J. Elongin B/C recruitment regulates substrate binding by CIS. J. Biol. Chem. 2008, 283, 21334–21346. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Masuhara, M.; Mitsui, K.; Yokouchi, M.; Ohtsubo, M.; Misawa, H.; Miyajima, A.; Yoshimura, A. CIS, a cytokine inducible SH2 protein, is a target of the JAK-STAT5 pathway and modulates STAT5 activation. Blood 1997, 89, 3148–3154. [Google Scholar] [CrossRef]
- Ketteler, R.; Moghraby, C.S.; Hsiao, J.G.; Sandra, O.; Lodish, H.F.; Klingmuller, U. The cytokine-inducible Src homology domain-containing protein negatively regulates signaling by promoting apoptosis in erythroid progenitor cells. J. Biol. Chem. 2003, 278, 2654–2660. [Google Scholar] [CrossRef]
- Matsumoto, A.; Seki, Y.; Kubo, M.; Ohtsuka, S.; Suzuki, A.; Hayashi, I.; Tsuji, K.; Nakahata, T.; Okabe, M.; Yamada, S.; et al. Suppression of STAT5 functions in liver, mammary glands, and T cells in cytokine-inducible SH2 protein-1 (CIS1) transgenic mice. Mol. Cell. Biol. 1999, 19, 6396–6407. [Google Scholar] [CrossRef]
- Yang, X.O.; Zhang, H.; Kim, B.S.; Niu, X.; Peng, J.; Chen, Y.; Kerketta, R.; Lee, Y.H.; Chang, S.H.; Corry, D.B.; et al. The signaling suppressor CIS controls proallergic T cell development and allergic airway inflammation. Nat. Immunol. 2013, 14, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Palmer, D.C.; Guittard, G.C.; Franco, Z.; Crompton, J.G.; Eil, R.L.; Patel, S.J.; Ji, Y.; Van Panhuys, N.; Klebanoff, C.A.; Sukumar, M.; et al. Cish actively silences TCR signaling in CD8+ T cells to maintain tumor tolerance. J. Exp. Med. 2015, 212, 2095–2113. [Google Scholar] [CrossRef] [PubMed]
- Delconte, R.B.; Kolesnik, T.B.; Dagley, L.F.; Rautela, J.; Shi, W.; Putz, E.M.; Stannard, K.; Zhang, J.G.; Teh, C.; Firth, M.; et al. CIS is a potent checkpoint in NK cell-mediated tumor immunity. Nat. Immunol. 2016, 17, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Louis, C.; Souza-Fonseca-Guimaraes, F.; Yang, Y.; D’silva, D.; Kratina, T.; Dagley, L.; Hediyeh-Zadeh, S.; Rautela, J.; Masters, S.L.; Davis, M.J.; et al. NK cell-derived GM-CSF potentiates inflammatory arthritis and is negatively regulated by CIS. J. Exp. Med. 2020, 217, e20191421. [Google Scholar] [CrossRef]
- Naser, W.; Maymand, S.; Dlugolenski, D.; Basheer, F.; Ward, A.C. The role of cytokine-inducible SH2 domain-containing (CISH) protein in the regulation of basal and cytokine-mediated myelopoiesis. Int. J. Mol. Sci. 2023, 24, 12757. [Google Scholar] [CrossRef] [PubMed]
- Naser, W.; Maymand, S.; Rivera, L.R.; Connor, T.; Liongue, C.; Smith, C.M.; Aston-Mourney, K.; McCulloch, D.R.; McGee, S.L.; Ward, A.C. Cytokine-inducible SH2 domain containing protein contributes to regulation of adiposity, food intake, and glucose metabolism. FASEB J. 2022, 36, e22320. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.S.; Noor, S.M.; Fraser, F.W.; Sertori, R.; Liongue, C.; Ward, A.C. Regulation of embryonic hematopoiesis by a cytokine-inducible SH2 domain homolog in zebrafish. J. Immunol. 2014, 192, 5739–5748. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Gertsenstein, M.; Vintersten, K.; Behringer, R. Staining whole mouse embryos for beta-galactosidase (lacZ) activity. CSH Protoc. 2007, 2007, pdb-prot4725. [Google Scholar]
- Liu, J.; Zhang, J.; Ginzburg, Y.; Li, H.; Xue, F.; De Franceschi, L.; Chasis, J.A.; Mohandas, N.; An, X. Quantitative analysis of murine terminal erythroid differentiation in vivo: Novel method to study normal and disordered erythropoiesis. Blood 2013, 121, e43–e49. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Richmond, T.D.; Chohan, M.; Barber, D.L. Turning cells red: Signal transduction mediated by erythropoietin. Trends Cell. Biol. 2005, 15, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Van den Akker, E.; van Dijk, T.; Parren-van Amelsvoort, M.; Grossmann, K.S.; Schaeper, U.; Toney-Earley, K.; Waltz, S.E.; Löwenberg, B.; von Lindern, M. Tyrosine kinase receptor RON functions downstream of the erythropoietin receptor to induce expansion of erythroid progenitors. Blood 2004, 103, 4457–4465. [Google Scholar] [CrossRef][Green Version]
- Povinelli, B.J.; Nemeth, M.J. Wnt5a regulates hematopoietic stem cell proliferation and repopulation through the Ryk receptor. Stem Cells 2014, 32, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, M.D.; Allen, M.A.; Bensard, C.L.; Wang, X.; Schwinn, M.K.; Qin, B.; Long, H.W.; Daniels, D.L.; Hahn, W.C.; Dowell, R.D.; et al. HIF1A employs CDK8-mediator to stimulate RNAPII elongation in response to hypoxia. Cell 2013, 153, 1327–1339. [Google Scholar] [CrossRef] [PubMed]
- Rademacher, M.; Kuhn, H.; Borchert, A. Systemic deficiency of mouse arachidonate 15-lipoxygenase induces defective erythropoiesis and transgenic expression of the human enzyme rescues this phenotype. FASEB J. 2020, 34, 14318–14335. [Google Scholar] [CrossRef] [PubMed]
- Da Cunha, A.F.; Brugnerotto, A.F.; Duarte, A.S.; Lanaro, C.; Costa, G.G.L.; Saad, S.T.O.; Costa, F.F. Global gene expression reveals a set of new genes involved in the modification of cells during erythroid differentiation. Cell Prolif. 2010, 43, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Moriggl, R.; Sexl, V.; Kenner, L.; Duntsch, C.; Stangl, K.; Gingras, S.; Hoffmeyer, A.; Bauer, A.; Piekorz, R.; Wang, D.; et al. Stat5 tetramer formation is associated with leukemogenesis. Cancer Cell 2005, 7, 87–99. [Google Scholar] [CrossRef]
- Wierenga, A.T.; Vellenga, E.; Schuringa, J.J. Maximal STAT5-induced proliferation and self-renewal at intermediate STAT5 activity levels. Mol. Cell. Biol. 2008, 28, 6668–6680. [Google Scholar] [CrossRef]
- Debierre-Grockiego, F. Anti-apoptotic role of STAT5 in haematopoietic cells and in the pathogenesis of malignancies. Apoptosis 2004, 9, 717–728. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RBC (×105/mm3) | Hb (g/dL) | HCT (%) | MCV (fL) | MCH (g/dL) | MCHC (%) | |
|---|---|---|---|---|---|---|
| WT | 11.3 ± 0.1 | 19.0 ± 0.2 | 55.7 ± 0.6 | 48.6 ± 0.1 | 16.8 ± 0.2 | 34.6 ± 0.4 |
| KO | 11.3 ± 0.2 | 18.0 ± 0.3 * | 51.9 ± 1.4 * | 49.7 ± 0.2 *** | 16.7 ± 0.2 | 33.2 ± 0.3 * |
| RBC (×105/mm3) | Hb (g/dL) | HCT (%) | MCV (fL) | MCH (g/dL) | MCHC (%) | |
|---|---|---|---|---|---|---|
| WT | 11.0 ± 0.1 | 18.4 ± 0.2 | 55.1 ± 0.5 | 48.8 ± 0.2 | 16.5 ± 0.2 | 34.4 ± 0.5 |
| WT + EPO 4d | 11.9 ± 0.4 | 20.1 ± 0.7 # | 59.5 ± 1.5 # | 51.3 ± 0.6 ## | 17.2 ± 0.2 # | 33.1 ± 0.4 |
| WT +EPO 6d | 12.8 ± 0.5 ## | 21.9 ± 2.2 ### | 68.0 ± 1.5 ## | 55.7 ± 0.2 ### | 17.7 ± 0.3 # | 31.7 ± 0.6 # |
| WT + EPO 6 + 2d | 12.9 ± 0.6 ## | 21.6 ± 0.8 ### | 66.2 ± 2.0 ## | 54.3 ± 1.2 ### | 17.4 ± 0.3 # | 31.7 ± 0.2 # |
| KO | 11.6 ± 0.6 | 16.7 ± 0.5 ** | 47.8 ± 2.4 * | 49.5 ± 0.2 * | 16.7 ± 0.1 | 33.0 ± 0.2 * |
| KO + EPO 4d | 10.5 ± 0.8 | 17.1 ± 0.8 * | 49.4 ± 2.4 * | 52.5 ± 1.0 ## | 17.4 ± 0.3 # | 33.6 ± 0.3 |
| KO + EPO 6d | 12.3 ± 0.9 | 19.7 ± 0.7 # | 60.8 ± 2.6 # | 55.0 ± 1.0 ### | 18.2 ± 0.7 # | 31.9 ± 0.2 # |
| KO + EPO 6 + 2d | 13.0 ± 0.9 | 20.4 ± 0.8 # | 66.0 ± 2.7 ## | 54.0 ± 0.6 ### | 17.2 ± 0.1 # | 31.9 ± 0.3 ## |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maymand, S.; Lakkavaram, A.L.; Naser, W.; Rasighaemi, P.; Dlugolenski, D.; Liongue, C.; Stambas, J.; de Koning-Ward, T.F.; Ward, A.C. Role of Cytokine-Inducible SH2 Domain-Containing (CISH) Protein in the Regulation of Erythropoiesis. Biomolecules 2023, 13, 1510. https://doi.org/10.3390/biom13101510
Maymand S, Lakkavaram AL, Naser W, Rasighaemi P, Dlugolenski D, Liongue C, Stambas J, de Koning-Ward TF, Ward AC. Role of Cytokine-Inducible SH2 Domain-Containing (CISH) Protein in the Regulation of Erythropoiesis. Biomolecules. 2023; 13(10):1510. https://doi.org/10.3390/biom13101510
Chicago/Turabian StyleMaymand, Saeed, Asha L. Lakkavaram, Wasan Naser, Parisa Rasighaemi, Daniel Dlugolenski, Clifford Liongue, John Stambas, Tania F. de Koning-Ward, and Alister C. Ward. 2023. "Role of Cytokine-Inducible SH2 Domain-Containing (CISH) Protein in the Regulation of Erythropoiesis" Biomolecules 13, no. 10: 1510. https://doi.org/10.3390/biom13101510
APA StyleMaymand, S., Lakkavaram, A. L., Naser, W., Rasighaemi, P., Dlugolenski, D., Liongue, C., Stambas, J., de Koning-Ward, T. F., & Ward, A. C. (2023). Role of Cytokine-Inducible SH2 Domain-Containing (CISH) Protein in the Regulation of Erythropoiesis. Biomolecules, 13(10), 1510. https://doi.org/10.3390/biom13101510