
PAGln, an Atrial Fibrillation-Linked Gut Microbial Metabolite, Acts as a Promoter of Atrial Myocyte Injury


Abstract
:1. Introduction
2. Materials and Methods
2.1. Relative Abundance of Gut Microbial PorA Enzyme
2.2. Study Population
2.3. Clinical Characteristics
2.4. Plasma PAGln Measurement
2.5. Cell Culture and Intervention
2.6. Apoptosis Assays
2.7. Reactive Oxygen Species Assay
2.8. MTT Assay
2.9. Measurement of Superoxide Dismutase (SOD) and NADPH Oxidase (NOX) Activities
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
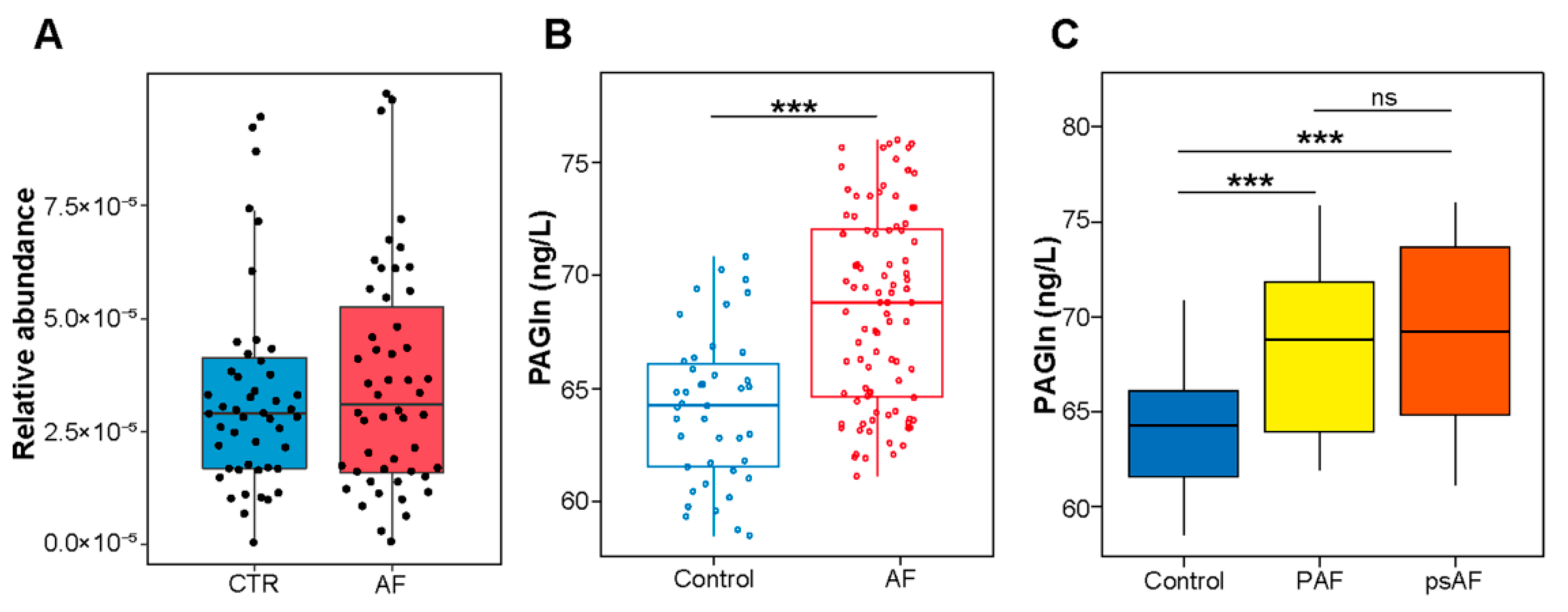
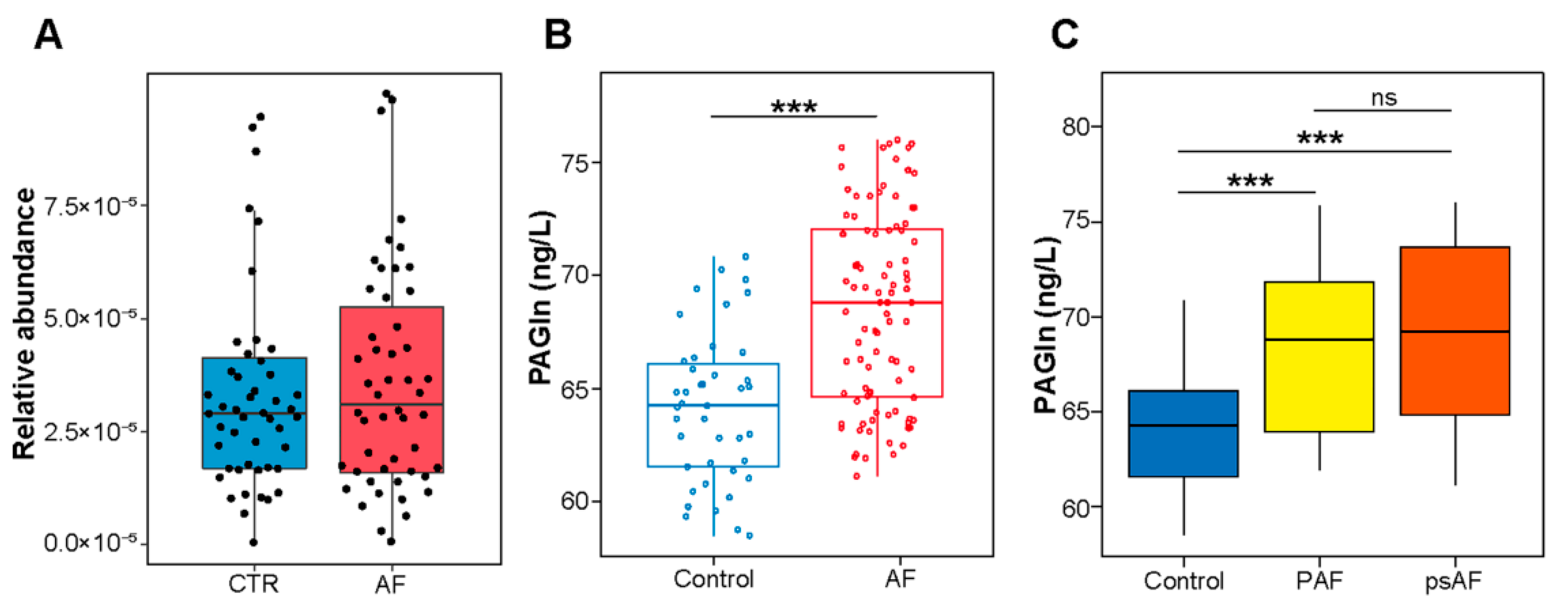
3.1. Association between PAGln and AF
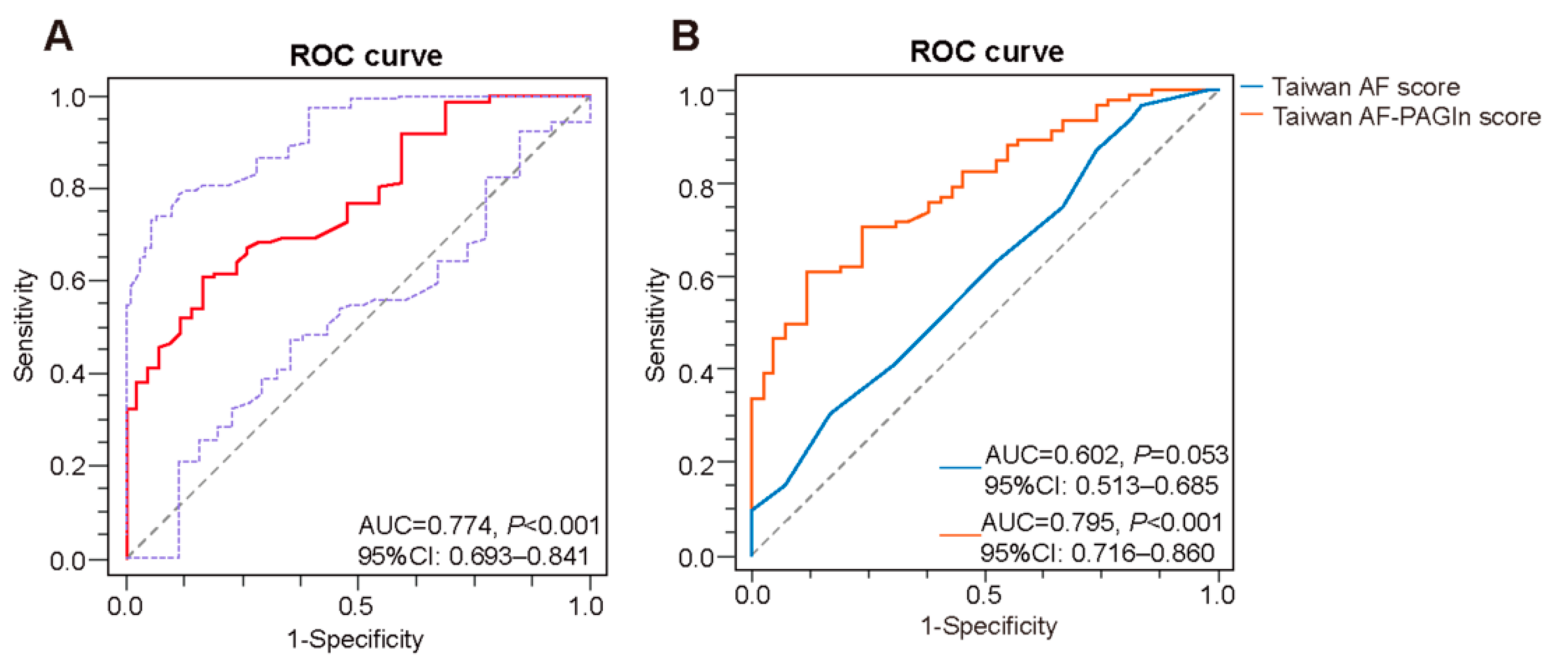
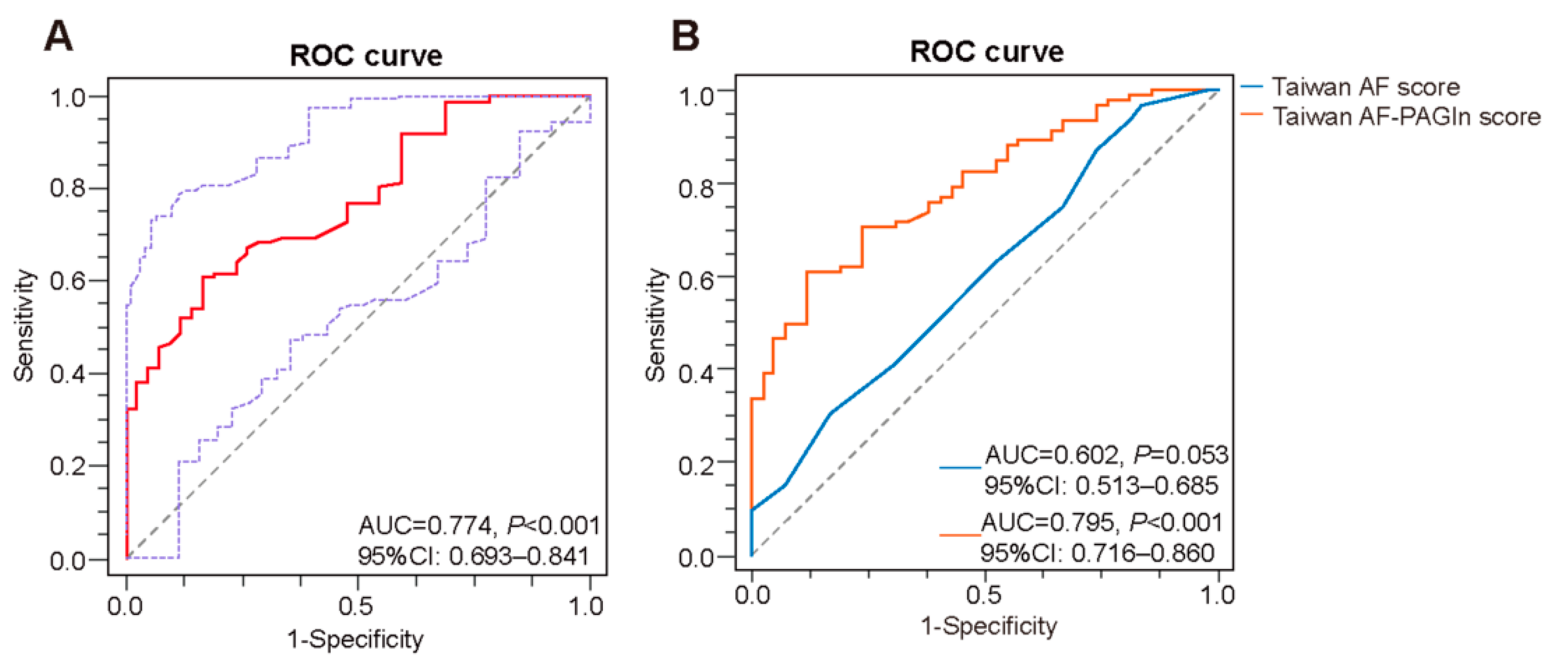
3.2. Plasma PAGln in the Prediction of AF
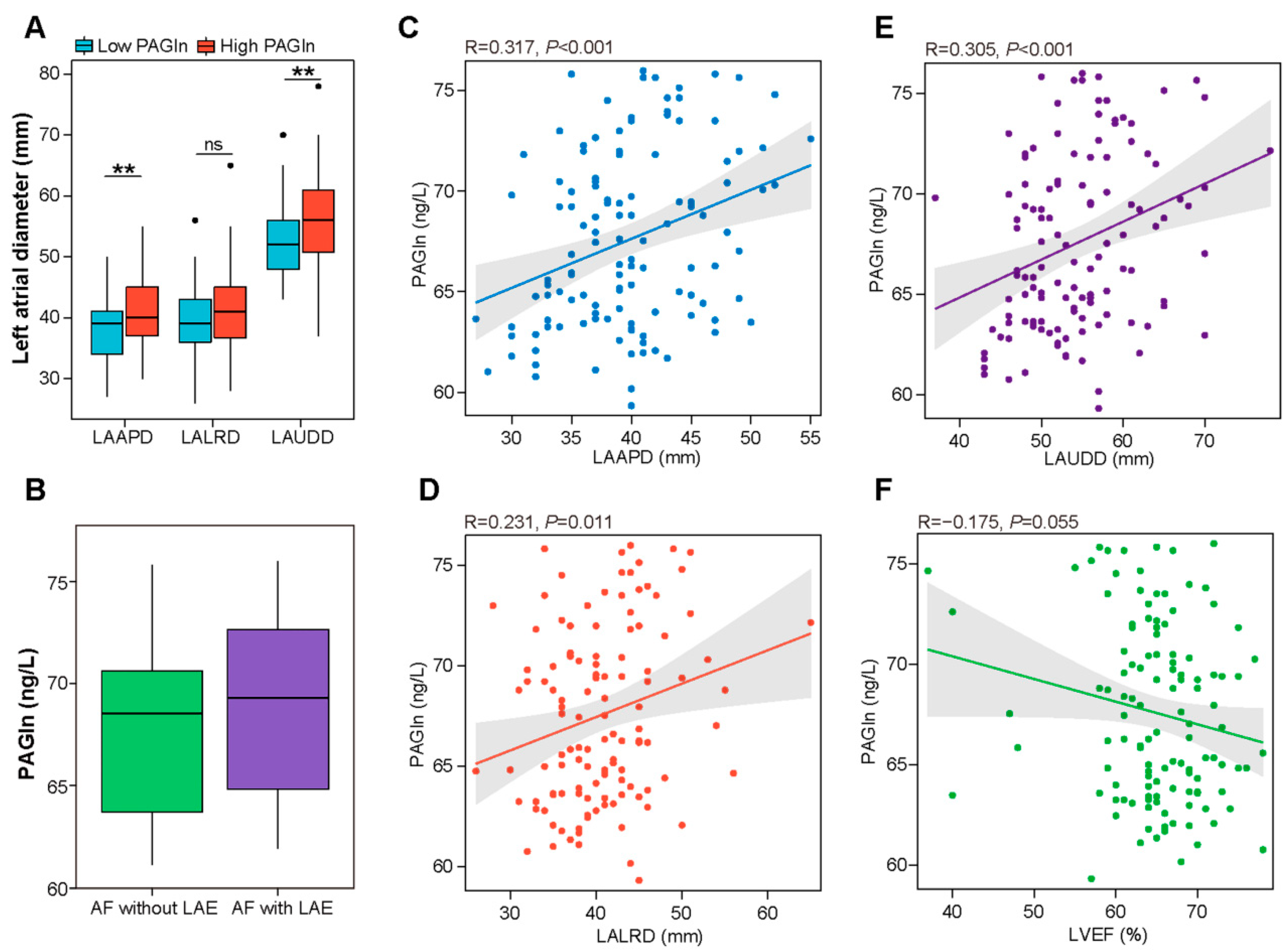
3.3. Correlation between Plasma PAGln and Structural Parameters of the Left Atrium
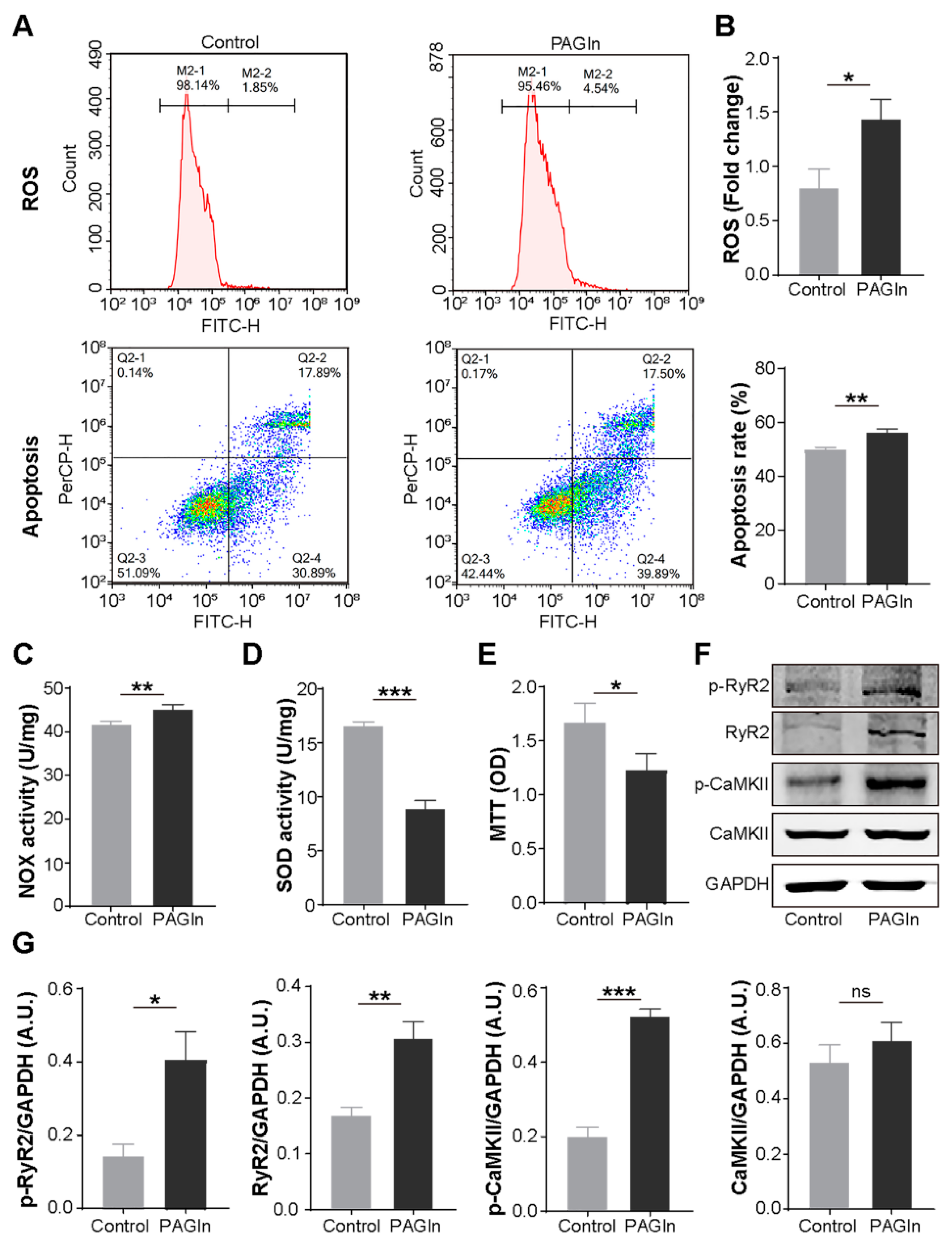
3.4. PAGln Aggravates Atrial Myocyte Oxidative Stress and Apoptosis
3.5. PAGln Induces Atrial Myocyte Activation of CaMKII and RyR2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mehta, N.K.; Strickling, J.; Mark, E.; Swinehart, S.; Puthumana, J.; Lavie, C.J.; Haines, D.E.; Franklin, B.A. Beyond cardioversion, ablation and pharmacotherapies: Risk factors, lifestyle change and behavioral counseling strategies in the prevention and treatment of atrial fibrillation. Prog. Cardiovasc. Dis. 2021, 66, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Wang, Z. Cross-Talk between Gut Microbiota and Heart via the Routes of Metabolite and Immunity. Gastroenterol. Res. Pract. 2018, 2018, 6458094. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, M.; Weeks, T.L.; Hazen, S.L. Gut Microbiota and Cardiovascular Disease. Circ. Res. 2020, 127, 553–570. [Google Scholar] [CrossRef]
- Tang, W.H.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef]
- Kazemian, N.; Mahmoudi, M.; Halperin, F.; Wu, J.C.; Pakpour, S. Gut microbiota and cardiovascular disease: Opportunities and challenges. Microbiome 2020, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Gawałko, M.; Agbaedeng, T.A.; Saljic, A.; Müller, D.N.; Wilck, N.; Schnabel, R.; Penders, J.; Rienstra, M.; van Gelder, I.; Jespersen, T.; et al. Gut microbiota, dysbiosis and atrial fibrillation. Arrhythmogenic mechanisms and potential clinical implications. Cardiovasc. Res. 2021, cvab292. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Y. Gut microbiota derived metabolites in cardiovascular health and disease. Protein. Cell. 2018, 9, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Wu, J.; Ye, B.C.; Qi, N. Gut Microbiota-Derived Metabolites in the Development of Diseases. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 6658674. [Google Scholar] [CrossRef]
- Nemet, I.; Saha, P.P.; Gupta, N.; Zhu, W.; Romano, K.A.; Skye, S.M.; Cajka, T.; Mohan, M.L.; Li, L.; Wu, Y.; et al. A Cardiovascular Disease-Linked Gut Microbial Metabolite Acts via Adrenergic Receptors. Cell 2020, 180, 862–877.e22. [Google Scholar] [CrossRef]
- Kilts, J.D.; Gerhardt, M.A.; Richardson, M.D.; Sreeram, G.; Mackensen, G.B.; Grocott, H.P.; White, W.D.; Davis, R.D.; Newman, M.F.; Reves, J.G.; et al. Beta(2)-adrenergic and several other G protein-coupled receptors in human atrial membranes activate both G(s) and G(i). Circ. Res. 2000, 87, 705–709. [Google Scholar] [CrossRef]
- Zefirov, T.L.; Khisamieva, L.I.; Ziyatdinova, N.I.; Zefirov, A.L. Selective Blockade of α2-Adrenoceptor Subtypes Modulates Contractility of Rat Myocardium. Bull. Exp. Biol. Med. 2016, 162, 177–179. [Google Scholar] [CrossRef]
- Sharifov, O.F.; Fedorov, V.V.; Beloshapko, G.G.; Glukhov, A.V.; Yushmanova, A.V.; Rosenshtraukh, L.V. Roles of adrenergic and cholinergic stimulation in spontaneous atrial fibrillation in dogs. J. Am. Coll. Cardiol. 2004, 43, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Scherlag, B.J.; Kem, D.C.; Zillner, C.; Male, S.; Thirunavukkarasu, S.; Shen, X.; Pitha, J.V.; Cunningham, M.W.; Lazzara, R.; et al. Atrial tachycardia provoked in the presence of activating autoantibodies to β2-adrenergic receptor in the rabbit. Heart Rhythm 2013, 10, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Altschuld, R.A.; Billman, G.E. beta(2)-Adrenoceptors and ventricular fibrillation. Pharmacol. Ther. 2000, 88, 1–14. [Google Scholar] [CrossRef]
- Casabella-Ramón, S.; Jiménez-Sábado, V.; Tarifa, C.; Casellas, S.; Lu, T.T.; Izquierdo-Castro, P.; Gich, I.; Jiménez, M.; Ginel, A.; Guerra, J.M.; et al. Impact of R-Carvedilol on β2-Adrenergic Receptor-Mediated Spontaneous Calcium Release in Human Atrial Myocytes. Biomedicines 2022, 10, 1759. [Google Scholar] [CrossRef]
- Lang, D.; Holzem, K.; Kang, C.; Xiao, M.; Hwang, H.J.; Ewald, G.A.; Yamada, K.A.; Efimov, I.R. Arrhythmogenic remodeling of β2 versus β1 adrenergic signaling in the human failing heart. Circ. Arrhythm. Electrophysiol. 2015, 8, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Desantiago, J.; Ai, X.; Islam, M.; Acuna, G.; Ziolo, M.T.; Bers, D.M.; Pogwizd, S.M. Arrhythmogenic effects of beta2-adrenergic stimulation in the failing heart are attributable to enhanced sarcoplasmic reticulum Ca load. Circ. Res. 2008, 102, 1389–1397. [Google Scholar] [CrossRef]
- Zuo, K.; Li, J.; Li, K.; Hu, C.; Gao, Y.; Chen, M.; Hu, R.; Liu, Y.; Chi, H.; Wang, H.; et al. Disordered gut microbiota and alterations in metabolic patterns are associated with atrial fibrillation. Gigascience 2019, 8, giz058. [Google Scholar] [CrossRef] [PubMed]
- Hindricks, G.; Potpara, T.; Dagres, N.; Arbelo, E.; Bax, J.J.; Blomström-Lundqvist, C.; Boriani, G.; Castella, M.; Dan, G.A.; Dilaveris, P.E.; et al. ESC Scientific Document Group. 2020 ESC Guidelines for the diagnosis and management of atrial fibrillation developed in collaboration with the European Association for Cardio-Thoracic Surgery (EACTS): The Task Force for the diagnosis and management of atrial fibrillation of the European Society of Cardiology (ESC) Developed with the special contribution of the European Heart Rhythm Association (EHRA) of the ESC. Eur. Heart J. 2021, 42, 373–498. [Google Scholar] [CrossRef]
- Jiang, Q.; Dong, X.; Hu, D.; Chen, L.; Luo, Y. Aquaporin 4 inhibition alleviates myocardial ischemia-reperfusion injury by restraining cardiomyocyte pyroptosis. Bioengineered 2021, 12, 9021–9030. [Google Scholar] [CrossRef]
- Li, Y.; Xia, J.; Jiang, N.; Xian, Y.; Ju, H.; Wei, Y.; Zhang, X. Corin protects H2O2-induced apoptosis through PI3K/AKT and NF-κB pathway in cardiomyocytes. Biomed. Pharmacother. 2018, 97, 594–599. [Google Scholar] [CrossRef]
- Chao, T.F.; Chiang, C.E.; Chen, T.J.; Liao, J.N.; Tuan, T.C.; Chen, S.A. Clinical Risk Score for the Prediction of Incident Atrial Fibrillation: Derivation in 7 220 654 Taiwan Patients With 438 930 Incident Atrial Fibrillations During a 16-Year Follow-Up. J. Am. Heart Assoc. 2021, 10, e020194. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Peng, L.; Ghista, D.N.; Wong, K.K.L. Left Atrial Remodeling Mechanisms Associated with Atrial Fibrillation. Cardiovasc. Eng. Technol. 2021, 12, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Dai, D.; Hu, J.; Zhu, J.; Lu, L.; Tao, G.; Zhang, R. Dexmedetomidine improves cardiac function and protects against maladaptive remodeling following myocardial infarction. Mol. Med. Rep. 2019, 20, 5183–5189. [Google Scholar] [CrossRef]
- Dobrev, D.; Wehrens, X.H. Calmodulin kinase II, sarcoplasmic reticulum Ca2+ leak, and atrial fibrillation. Trends. Cardiovasc. Med. 2010, 20, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Zuo, K.; Yin, X.; Li, K.; Zhang, J.; Wang, P.; Jiao, J.; Liu, Z.; Liu, X.; Liu, J.; Li, J.; et al. Different Types of Atrial Fibrillation Share Patterns of Gut Microbiota Dysbiosis. mSphere 2020, 5, e00071-20. [Google Scholar] [CrossRef] [PubMed]
- Zuo, K.; Li, J.; Wang, P.; Liu, Y.; Liu, Z.; Yin, X.; Liu, X.; Yang, X. Duration of Persistent Atrial Fibrillation Is Associated with Alterations in Human Gut Microbiota and Metabolic Phenotypes. mSystems 2019, 4, e00422-19. [Google Scholar] [CrossRef]
- Li, J.; Zuo, K.; Zhang, J.; Hu, C.; Wang, P.; Jiao, J.; Liu, Z.; Yin, X.; Liu, X.; Li, K.; et al. Shifts in gut microbiome and metabolome are associated with risk of recurrent atrial fibrillation. J. Cell. Mol. Med. 2020, 24, 13356–13369. [Google Scholar] [CrossRef]
- Fang, C.; Zuo, K.; Fu, Y.; Li, J.; Wang, H.; Xu, L.; Yang, X. Dysbiosis of Gut Microbiota and Metabolite Phenylacetylglutamine in Coronary Artery Disease Patients with Stent Stenosis. Front. Cardiovasc. Med. 2022, 9, 832092. [Google Scholar] [CrossRef]
- Yu, F.; Li, X.; Feng, X.; Wei, M.; Luo, Y.; Zhao, T.; Xiao, B.; Xia, J. Phenylacetylglutamine, a Novel Biomarker in Acute Ischemic Stroke. Front. Cardiovasc. Med. 2021, 8, 798765. [Google Scholar] [CrossRef]
- Ren, X.; Wang, X.; Yuan, M.; Tian, C.; Li, H.; Yang, X.; Li, X.; Li, Y.; Yang, Y.; Liu, N.; et al. Mechanisms and Treatments of Oxidative Stress in Atrial Fibrillation. Curr. Pharm. Des. 2018, 24, 3062–3071. [Google Scholar] [CrossRef] [PubMed]
- Burashnikov, A.; Antzelevitch, C. Late-phase 3 EAD. A unique mechanism contributing to initiation of atrial fibrillation. Pacing Clin. Electrophysiol. 2006, 29, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Mesubi, O.O.; Anderson, M.E. Atrial remodelling in atrial fibrillation: CaMKII as a nodal proarrhythmic signal. Cardiovasc. Res. 2016, 109, 542–557. [Google Scholar] [CrossRef]
- Jalife, J.; Kaur, K. Atrial remodeling, fibrosis, and atrial fibrillation. Trends Cardiovasc. Med. 2015, 25, 475–484. [Google Scholar] [CrossRef]
- Dridi, H.; Kushnir, A.; Zalk, R.; Yuan, Q.; Melville, Z.; Marks, A.R. Intracellular calcium leak in heart failure and atrial fibrillation: A unifying mechanism and therapeutic target. Nat. Rev. Cardiol. 2020, 17, 732–747. [Google Scholar] [CrossRef]
- Trappe, K.; Thomas, D.; Bikou, O.; Kelemen, K.; Lugenbiel, P.; Voss, F.; Becker, R.; Katus, H.A.; Bauer, A. Suppression of persistent atrial fibrillation by genetic knockdown of caspase 3: A pre-clinical pilot study. Eur. Heart J. 2013, 34, 147–157. [Google Scholar] [CrossRef]
- Aimé-Sempé, C.; Folliguet, T.; Rücker-Martin, C.; Krajewska, M.; Krajewska, S.; Heimburger, M.; Aubier, M.; Mercadier, J.J.; Reed, J.C.; Hatem, S.N. Myocardial cell death in fibrillating and dilated human right atria. J. Am. Coll. Cardiol. 1999, 34, 1577–1586. [Google Scholar] [CrossRef]
- Hu, Y.F.; Chen, Y.J.; Lin, Y.J.; Chen, S.A. Inflammation and the pathogenesis of atrial fibrillation. Nat. Rev. Cardiol. 2015, 12, 230–243. [Google Scholar] [CrossRef]
- Mayyas, F.; Alzoubi, K.H.; Van Wagoner, D.R. Impact of aldosterone antagonists on the substrate for atrial fibrillation: Aldosterone promotes oxidative stress and atrial structural/electrical remodeling. Int. J. Cardiol. 2013, 168, 5135–5142. [Google Scholar] [CrossRef]
- Karam, B.S.; Chavez-Moreno, A.; Koh, W.; Akar, J.G.; Akar, F.G. Oxidative stress and inflammation as central mediators of atrial fibrillation in obesity and diabetes. Cardiovasc. Diabetol. 2017, 16, 120. [Google Scholar] [CrossRef]
- Nattel, S.; Burstein, B.; Dobrev, D. Atrial remodeling and atrial fibrillation: Mechanisms and implications. Circ. Arrhythmia Electrophysiol. 2008, 1, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.; Abhayaratna, W.P. Left Atrial Reverse Remodeling: Mechanisms, Evaluation, and Clinical Significance. JACC Cardiovasc. Imaging. 2017, 10, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Nolla-Colomer, C.; Casabella-Ramon, S.; Jimenez-Sabado, V.; Vallmitjana, A.; Tarifa, C.; Herraiz-Martínez, A.; Llach, A.; Tauron, M.; Montiel, J.; Cinca, J.; et al. β2-adrenergic stimulation potentiates spontaneous calcium release by increasing signal mass and co-activation of ryanodine receptor clusters. Acta Physiol. 2022, 234, e13736. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | AF | p Value | |
|---|---|---|---|
| Number | 42 | 92 | |
| Male, % | 15 (35.71) | 50 (54.35) | 0.062 |
| HTN, % | 24 (57.14) | 49 (53.26) | 0.712 |
| DM, % | 5 (11.90) | 25 (27.17) | 0.073 |
| CAD, % | 0 (0.00) | 7 (7.61) | 0.098 |
| Smoking, % | 9 (21.43) | 15 (16.30) | 0.476 |
| Drinking, % | 10 (23.81) | 14 (15.22) | 0.235 |
| Age, years | 61.52 ± 9.83 | 65.00 ± 9.88 | 0.061 |
| BMI, kg/m2 | 25.99 ± 3.40 | 26.07 ± 3.60 | 0.918 |
| WBC, ×109/L | 5.90 ± 1.17 | 5.95 ± 1.49 | 0.883 |
| HGB, g/L | 129.28 ± 9.44 | 138.87 ± 17.74 | 0.002 ** |
| PLT, ×109/L | 224.56 ± 40.10 | 204.10 ± 60.54 | 0.173 |
| TC, mmol/L | 4.36 ± 1.06 | 4.06 ± 0.88 | 0.101 |
| TG, mmol/L | 0.95 (0.77, 1.89) | 1.19 (0.92, 1.49) | 0.257 |
| AST, U/L | 17.40 ± 6.67 | 19.86 ± 6.29 | 0.053 |
| ALT, U/L | 13.90 (10.40, 19.30) | 17.00 (13.00, 23.00) | 0.015 * |
| sCr, μmol/L | 65.39 ± 15.41 | 70.57 ± 14.49 | 0.076 |
| cTNI, ng/mL | 0.00 (0.00, 0.00) | 0.00 (0.00,0.01) | 0.266 |
| Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|
| OR (95% CI) | p Value | OR (95% CI) | p Value | |
| Male | 0.467 (0.220–0.991) | 0.047 * | ||
| DM | 0.362 (0.128–1.025) | 0.056 | ||
| Age | 1.036 (0.998–1.075) | 0.063 | 1.103 (1.026–1.186) | 0.008 ** |
| HGB | 1.037 (1.003–1.072) | 0.032 * | ||
| AST | 1.081 (0.999–1.170) | 0.054 | ||
| sCr | 1.026 (0.997–1.055) | 0.079 | ||
| PAGln | 1.331 (1.183–1.502) | <0.001 *** | 1.437 (1.143–1.806) | 0.002 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, C.; Zuo, K.; Jiao, K.; Zhu, X.; Fu, Y.; Zhong, J.; Xu, L.; Yang, X. PAGln, an Atrial Fibrillation-Linked Gut Microbial Metabolite, Acts as a Promoter of Atrial Myocyte Injury. Biomolecules 2022, 12, 1120. https://doi.org/10.3390/biom12081120
Fang C, Zuo K, Jiao K, Zhu X, Fu Y, Zhong J, Xu L, Yang X. PAGln, an Atrial Fibrillation-Linked Gut Microbial Metabolite, Acts as a Promoter of Atrial Myocyte Injury. Biomolecules. 2022; 12(8):1120. https://doi.org/10.3390/biom12081120
Chicago/Turabian StyleFang, Chen, Kun Zuo, Kaicheng Jiao, Xiaoming Zhu, Yuan Fu, Jiuchang Zhong, Li Xu, and Xinchun Yang. 2022. "PAGln, an Atrial Fibrillation-Linked Gut Microbial Metabolite, Acts as a Promoter of Atrial Myocyte Injury" Biomolecules 12, no. 8: 1120. https://doi.org/10.3390/biom12081120
APA StyleFang, C., Zuo, K., Jiao, K., Zhu, X., Fu, Y., Zhong, J., Xu, L., & Yang, X. (2022). PAGln, an Atrial Fibrillation-Linked Gut Microbial Metabolite, Acts as a Promoter of Atrial Myocyte Injury. Biomolecules, 12(8), 1120. https://doi.org/10.3390/biom12081120