
Biotechnology, Bioengineering and Applications of Bacillus Nattokinase


Abstract
1. Nattokinase Is a New Type of Thrombolytic Drug with Great Potential
2. Bacillus Is the Main Strain for Synthesizing Nattokinase

{kind=link}
{kind=link}
{kind=link}
| Strain | Source | References |
|---|---|---|
| Bacillus subtilis Natto B-12 | Natto | [17] |
| Bacillus sp. strain CK 11-4 | Chungkook-Jang | [23] |
| Bacillus sp. strain DJ-4 | Doen-Jang | [26] |
| Bacillus amyloliquefaciens DC-4 | Douchi | [19] |
| Bacillus subtilis DC33 | Douchi | [21] |
| Bacillus subtilis QK02 | Fermented soybeans | [27] |
| Bacillus subtilis LD-8547 | Douchi | [20] |
| Bacillus subtilis RJAS19 | Fermented soy products | [28] |
| Bacillus subtilis YJ1 | Fermented soy products | [29] |
| Bacillus subtilis TKU007 | Soil | [30] |
| Bacillus sublitis | Thailand | [31] |
| Bacillus subtilis LSSE-22 | Chinese soybean paste | [32] |
| Pseudomonas sp. TKU015 | Soil | [16] |
| Pseudomonas aeruginosa CMSS | Milk | [15] |
| Bacilluscereus VITSDVM3 | Rust | [25] |
| Bacillus subtilis WRL101 | Doen-jang | [24] |
| Bacillus velezensis KMU01 | Pickle | [33] |
| Bacillus subtilis VITMS 2 | Fermented milk of Vigna unguiculata. | [34] |
| Bacillussubtilis JNFE0126 | Natto | [18] |
| Bacillus subtilis K2 | Moromi | [35] |
| Bacillussubtilis LSSE-62 | Chinese soybean paste | [22] |
| Bacillus subtilis ICTF-1 | Ocean | [36] |
3. Structure and Catalytic Mechanism of Nattokinase
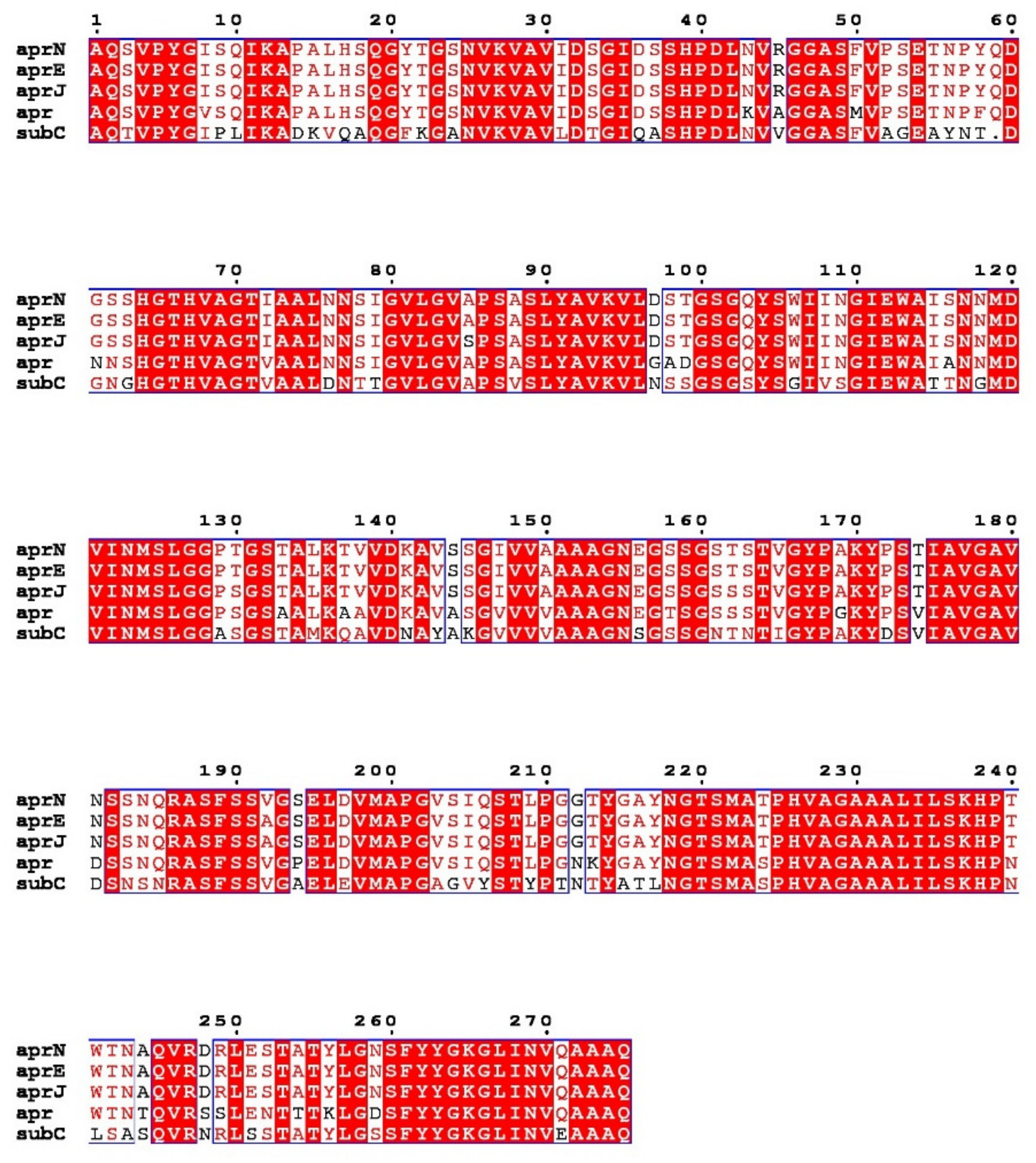
3.1. Nattokinase: The Only Member of the Alkaline Serine Protease Family with Thrombolytic Activity
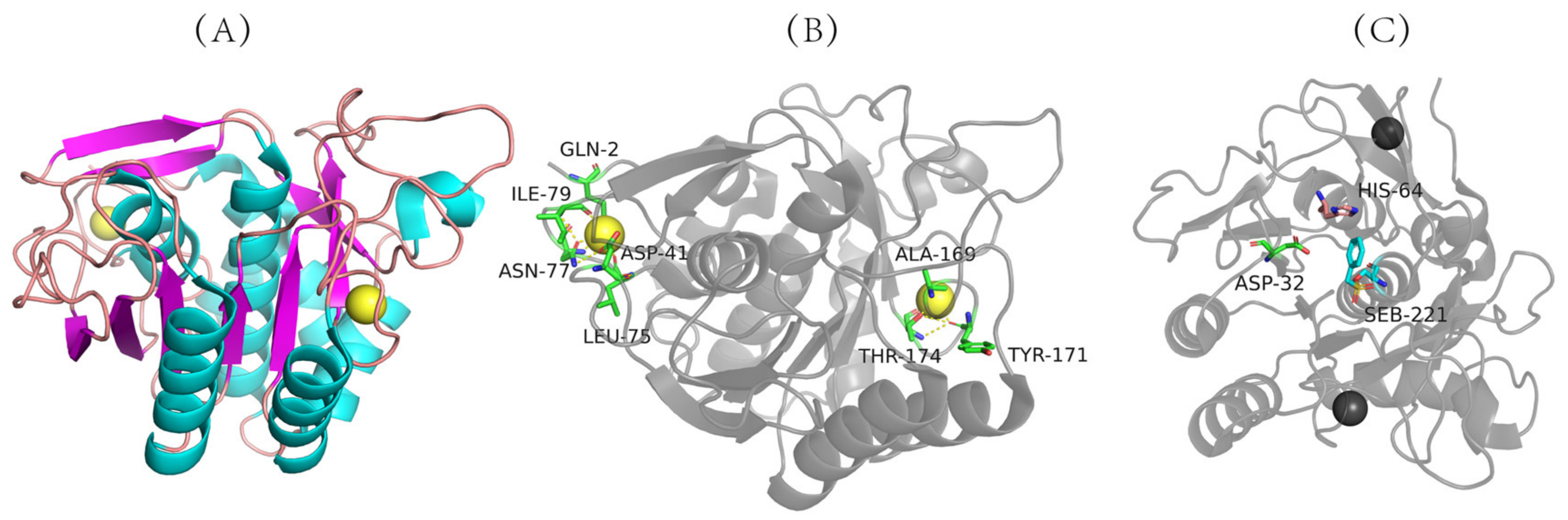
3.2. Structure and Reaction Mechanism of Nattokinase
3.3. The Propeptide of Nattokinase Is Involved in the Correct Folding of Nattokinase as an Intramolecular Chaperone
4. Study on Fermentation Process of Nattokinase
4.1. Production of Nattokinase by Solid Fermentation
4.2. Production of Nattokinase by Liquid Fermentation
| Strain | Carbon source | Nitrogen Source | Fermentation Time (h) | Fermentation pH | Mineral Salt | Fermentation Temperature (℃) | Enzyme Activity | References |
|---|---|---|---|---|---|---|---|---|
| Pseudomonas sp. TKU015 | 1% Shrimp shell waste | 1% Shrimp shell wastes | 48 | 7 | 0.1% K2HPO4 0.05% MgSO4·7H2O | 30 | 2.3 FU/mL (fibrin as a substrate) | [16] |
| Bacillus subtilis Natto B-12 | 2% Maltose | 2% Wheat | 60 | 7 | 0.5% NaCl, 0.1% KH2PO4, 0.4% K2HPO4, 0.05% MgSO4·7H2O, | 30 | 903 IU/mL (fibrin as a substrate) | [17] |
| Bacillus subtilis LD-8547 | 5% Rice powder | 4% Soybean powder 0.5% NH4NO3 | 72 | 7 | 0.5% NH4NO3, 0.01% CaC12, 0.7% MgSO4, 0.4% K2HPO4, 0.2% KH2PO4 | 35 | 3980 U/mL (fibrin as a substrate) | [20] |
| Bacillus subtilis TKU007 | 1.5% Shrimp shell powder | - | 48 | 7 | 0.1% K2HPO4, 0.05% MgSO4 | 30 | 0.35 U/mL (casein as a substrate) | [30] |
| Bacillus subtilis RJAS19 | 1% Lactose | 0.5% Peptone | 72 | 9 | - | 50 | - | [28] |
| Bacillus subtilis MX-6 | 1% Peptone | 0.3% Beef extract | 72 | 7.2–7.4 | 0.04% CaCl2, 0.04% MgSO4, 0.5% NaCl | 37 | - | [50] |
| Bacillus subtilis ICTF-1 | 3% Maltose | 0.5% Soy peptone, 1.5% Yeast extract | 48 | - | 0.15% K2HPO4·3H2O, 0.125% MgSO4·7H2O, 0.025% CaCl2·2H2O, 1% NaCl | 37 | 8645 U/mg (fibrin as a substrate) | [36] |
| Bacillus amyloliquefaciens DC-4 | 2% Dextrin | 0.15% Yeast extract 2% fibrin, 0.5% Peptone | 72 | 7.2 | 0.4% K2HPO4, 0.04% NaH2PO4, 0.3% CaCO3 | 37 | 407 U/mg (fibrin as a substrate) | [19] |
| Bacillus subtilis DC33 | 0.2% Galactose | 1.4% Tryptone | 72 | 7.2 | 0.1% KH2PO4, 0.2% Na2HPO4, 0.02% MgSO4, 0.3% CaCO3, 0.001% FeCl3, 0.001% LiSO4 | 37 | 447.4 U/mg (fibrin as a substrate) | [21] |
5. Enzymatic Activity Determination and Enzymatic Properties of Nattokinase
5.1. Determination of Fibrinolytic Activity of Nattokinase
| Method | Principle | Feature | Reference |
|---|---|---|---|
| Fibrin plate method | Nattokinase hydrolyzes fibrin on the plate to form a hydrolysis circle | The accuracy is not high, only used for preliminary characterization of enzyme activity | [55] |
| Fibrinolysis method | A standard curve was established using the time taken for urokinase to dissolve a certain amount of fibrin with different activity units. | Simple with limited precision | [56] |
| Tetrapeptide substrate method | Hydrolysis of Suc-Ala-Ala-Pro-Phe-pNA by nattokinase to produce chromogenic nitroaniline | Simple and fast, high sensitivity; only suitable for pure enzymes | [57] |
| HPLC method | The product of nattokinase hydrolysis of peptide A (LKRLKRFLKRLK) was detected by a diode array detector and an Eclipse XDB-C18 column. | The operation process is complicated | [58] |
| Milk plate method | Nattokinase hydrolyzes casein on the plate to form a hydrolysis circle | Simple and fast, used for preliminary characterization of nattokinase activity | [59] |
| Serum plate method | Nattokinase hydrolyzes fibrin in serum to form hydrolysis circle | Low cost, convenient and quick; easily affected by the quality of the prepared plate. | [60] |
| Enzyme-linked immunosorbent method | Determination of nattokinase activity by specific enzyme-linked adsorption | High sensitivity and specificity; complicated operation and high cost | [61] |
| UV spectrophotometer method | Determination of the fibrinolytic activity of nattokinase by measuring the change in absorbance of nattokinase fibrin hydrolysate at 275 nm | The operation process is complicated | [62] |
5.2. Biochemical Properties of Nattokinase
| Strain | MW (kDa) | Optimum pH | Optimum Temperature (°C) | Substrate Specificity | Specific Inhibitor | Metal Agonist | Metal Inhibitor | References |
|---|---|---|---|---|---|---|---|---|
| Bacillus subtilis Natto B-12 | 29 | 8 | 40 | - | - | Zn2+ | Al3+, Fe3+ | [17] |
| Bacillus subtilis BK-17 | 31 | - | - | H-D-Val-Leu-Lys-pNA | Chymostatin | - | Zn2+ | [67] |
| Bacillus sp. strain CK 11-4 | 28.2 | 10.5 | 70 | H-D-Val-Leu-Lys-rNA | PMSF | - | - | [23] |
| Bacillus sp. strain DJ-4 | 29 | 10 | 40 | - | PMSF | - | Cu2+, Zn2+ | [26] |
| Bacillus subtilis IMR-NK1 | 31.5 | 7.8 | 55 | N-succinyl-Ala-Ala-Pro-Phe-pNA | PMSF, NBS | - | - | [68] |
| Bacillus amyloliquefaciens DC-4 | 28 | 9 | 48 | N-succinyl-Ala-Ala-Pro-Phe-pNA | PMSF | - | - | [19] |
| Bacillus subtilis DC33 | 30 | 8 | 55 | N-Succinyl-Ala-Ala-Pro-Phe-pNA | PMSF, SBTI | - | Cu2+, Fe2+, Sn2+, Ag+, Ti2+ | [21] |
| Bacillus subtilis QK02 | 28 | 8.5 | 55 | - | PMSF | - | - | [27] |
| Bacillus subtilis LD-8547 | 30 | 8 | 50 | - | PMSF | Al3+, Zn2+ | Mn2+, Ba2+ | [20] |
| Bacillus subtilis TKU007 | 28–30 | 7 | 50 | - | - | - | - | [30] |
| Bacillus subtilis RJAS19 | 35 | 9.5 | 65 | - | - | - | - | [28] |
| Bacillus subtilis YJ1 | 27.5 | 8.5 | 50 | - | PMSF | Co2+, Ba2+ | Fe3+, Hg2+, Cu2+, Zn2+ | [29] |
| Bacillus subtilis TKU007 | 28–30 | 8 | 50 | - | PMSF | - | Ca2+, Cu2+ | [30] |
| Pseudomonas sp. TKU015 | 21–24 | 7 | 50 | N-benzoyl-Val-Gly-Arg-pNA and D-Val-Leu-Lys-pNA | PMSF | Fe2+ | Fe2+ | [16] |
| Pseudomonas aeruginosa CMSS | 27 | 5 | 45 | - | PMSF | Mn2+ | Ba2+, Mg2+ | [15] |
| Bacillus ereus VITSDVM3 | - | - | - | - | - | - | Cu2+, Ca2+ | [25] |
| Bacillus subtilis WRL101 | 29 | 11 | 47 | Met-Suc-Arg-Pro-Tyr-pNA(S-2586) | PMSF | Mg2+ | Zn2+, Cu2+ | [24] |
| Bacillus subtilis VTCC-DVN-12-01 | 27.7 | 9 | 65 | - | SDS | Mg2+ | - | [65] |
| Bacillus subtilis JNFE0126 | - | 7 | 40 | - | - | - | - | [18] |
| Bacillus subtilis VITMS 2 | 29 | 7 | 30 | N-Succinyl-Ala-Ala-Pro-Phe-pNA (S7338) | PMSF, EDTA | Ca2+, Zn2+, Co3+ | Hg2+, Cu2+ | [34] |
| Bacillus subtilis K2 | 26 | - | - | - | - | - | - | [35] |
| Bacillus subtilis ICTF-1 | 28 | 9 | 50 | - | PMSF | Zn2+, Fe2+, Hg2+ | - | [36] |
6. Molecular Modification and Expression of Nattokinase
6.1. Modification of Fibrinolytic Activity and Stability of Nattokinase
| Strains | Molecular Modification Strategy | Enzyme Activity Assay Method | Effect | References |
|---|---|---|---|---|
| Bacillus subtilis var. natto | Mutation based on the literature | Tetrapeptide substrate method | The I31L mutant increased catalytic efficiency | [57] |
| Bacillus subtilis var. natto | Mutation based on the literature | Tetrapeptide substrate method | The M222A/I31L mutant increased oxidative stability | [57] |
| Bacillus subtilis QK02 | Surface charge engineering, sequence alignment, and mutation based on the literature | Tetrapeptide substrate method | The Q59E mutant increased specific activity | [72] |
| Bacillus subtilis QK02 | Surface charge engineering, sequence alignment, and mutation based on the literature | Tetrapeptide substrate method | The S78T mutant improved acid stability | [72] |
| Bacillus subtilis QK02 | Surface charge engineering, sequence alignment, and mutation based on the literature | Tetrapeptide substrate method | The Y217K mutant enhanced acid and thermal stability | [72] |
| Bacillus subtilis QK02 | Surface charge engineering, sequence alignment, and mutation based on the literature | Tetrapeptide substrate method | The N218D mutant improved thermal stability | [72] |
| Bacillus subtilis QK02 | Surface charge engineering, sequence alignment, and mutation based on the literature | Tetrapeptide substrate method | The S78T/Y217K mutant improved acid stability | [72] |
| Bacillus subtilis var. natto strain AS 1.107 | Non-oxidative mutation of amino acid residues surrounding the catalytic residue Ser221 | Fibrin plate method and tetrapeptide substrate method | The T220S mutant increased oxidative stability | [43] |
| Bacillus subtilis var. natto strain AS 1.107 | Non-oxidative mutation of amino acid residues surrounding the catalytic residue Ser221 | Fibrin plate method and tetrapeptide substrate method | The M222A mutant increased oxidative stability | [43] |
| Bacillus licheniformis WX-02 | Deletion of protease genes and construction of high-efficiency expression system of nattokinase (PHY300PLK + P43 + SsacC + aprN + TamyL) | UV spectrophotometer method | Improved the synthesis efficiency of nattokinase in Bacillus licheniformis WX-02 | [73] |
| Bacillus licheniformis DW-02 | Manipulation of signal peptides and signal peptidases (signal peptide of AprE and signal peptidase SipV) | UV spectrophotometer method | Enhanced secretion efficiency of nattokinase in Bacillus licheniformis | [74] |
| Bacillus subtilis WB800 | Tandem promoter (PHpaII-PHpaII-PP43) | Fibrin plate method and UV spectrophotometer method | Improved the synthesis efficiency of nattokinase in Bacillus subtilis WB800 | [75] |
| Bacillus subtilis 168 | Sequence trimming and nucleotide optimization of the conserved region of the promoter PsrfA | HPLC method | High-efficiency self-inducible expression of nattokinase in Bacillus subtilis | [58] |
| strain Bacillus licheniformis DW2 | An optimized single-stranded Shine–Dalgarno (SD) sequence was inserted into the hairpin loop for better ribosome recognition and recruitment. | Milk plate method | Increased the fermentation yield of nattokinase in Bacillus licheniformis | [59] |
| Bacillus subtilis WB 800 | Deletion of protease genes | Tetrapeptide substrate method | Increased the fermentation yield of nattokinase in Bacillus subtilis WB 800 | [65] |
6.2. Construction of Nattokinase Expression System
7. Clinical Application of Nattokinase
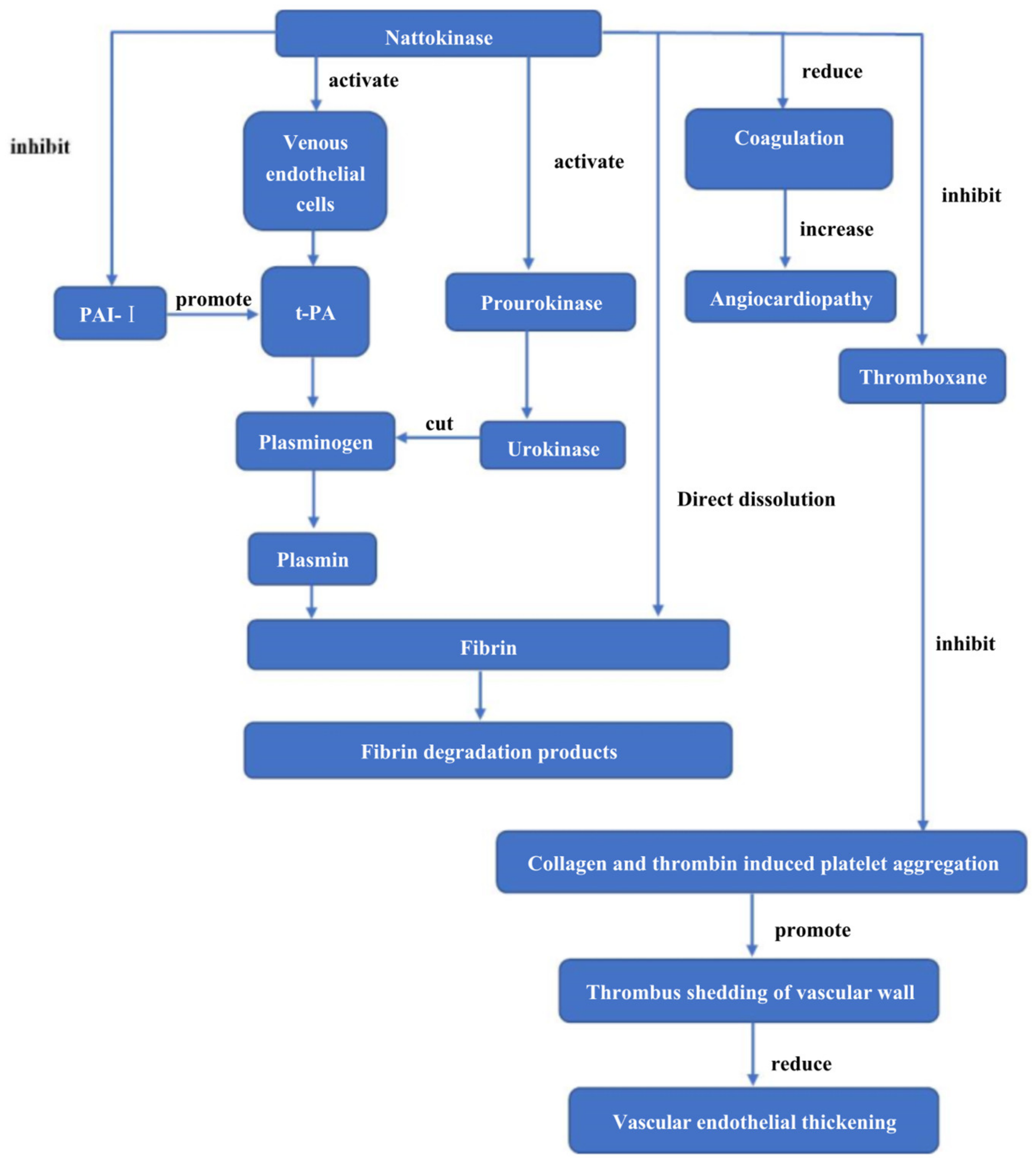
7.1. Study on Thrombolytic Effect of Nattokinase
7.2. Antihypertensive Effect of Nattokinase
7.3. Potential Applications of Nattokinase in Other Diseases
8. Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gianforcaro, A.; Kurz, M.; Guyette, F.; Callaway, C.W.; Rittenberger, J.C.; Elmer, J. Association of antiplatelet therapy with patient outcomes after out-of-hospital cardiac arrest. Resuscitation 2017, 121, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Furie, B.; Furie, B.C. Mechanisms of thrombus formation. N. Engl. J. Med. 2008, 359, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Nagata, C.; Wada, K.; Tamura, T.; Konishi, K.; Goto, Y.; Koda, S.; Kawachi, T.; Tsuji, M.; Nakamura, K. Dietary soy and natto intake and cardiovascular disease mortality in Japanese adults: The Takayama study. Am. J. Clin. Nutr. 2017, 105, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Sumi, H.; Hamada, H.; Tsushima, H.; Mihara, H.; Muraki, H. A novel fibrinolytic enzyme (nattokinase) in the vegetable cheese Natto; a typical and popular soybean food in the Japanese diet. Experientia 1987, 43, 1110–1111. [Google Scholar] [CrossRef]
- Guo, H.; Ban, Y.-H.; Cha, Y.; An, E.S.; Choi, J.; Seo, D.W.; Park, D.; Choi, E.-K.; Kim, Y.-B. Comparative anti-thrombotic activity and haemorrhagic adverse effect of nattokinase and tissue-type plasminogen activator. Food Sci. Biotechnol. 2019, 28, 1535–1542. [Google Scholar] [CrossRef]
- Wu, H.; Wang, H.; Xu, F.; Chen, J.; Duan, L.; Zhang, F. Acute toxicity and genotoxicity evaluations of Nattokinase, a promising agent for cardiovascular diseases prevention. Regul. Toxicol. Pharmacol. 2019, 103, 205–209. [Google Scholar] [CrossRef]
- Hsia, C.-H.; Shen, M.-C.; Lin, J.-S.; Wen, Y.-K.; Hwang, K.-L.; Cham, T.-M.; Yang, N.-C. Nattokinase decreases plasma levels of fibrinogen, factor VII, and factor VIII in human subjects. Nutr. Res. 2009, 29, 190–196. [Google Scholar] [CrossRef]
- Jensen, G.S.; Lenninger, M.; Ero, M.P.; Benson, K.F. Consumption of nattokinase is associated with reduced blood pressure and von Willebrand factor, a cardiovascular risk marker: Results from a randomized, double-blind, placebo-controlled, multicenter North American clinical trial. Integr. Blood Press. Control. 2016, 9, 95–104. [Google Scholar] [CrossRef]
- Chang, C.-H.; Chen, K.-T.; Lee, T.-H.; Wang, C.-H.; Kuo, Y.-W.; Chiu, Y.-H.; Hsieh, C.-L.; Wu, C.-J.; Chang, Y.-L. Effects of natto extract on endothelial injury in a rat model. Acta Med. Okayama 2010, 64, 399–406. [Google Scholar]
- Huang, Z.; Ng, T.K.; Chen, W.; Sun, X.; Huang, D.; Zheng, D.; Yi, J.; Xu, Y.; Zhuang, X.; Chen, S. Nattokinase Attenuates Retinal Neovascularization via Modulation of Nrf2/HO-1 and Glial Activation. Investig. Ophthalmol. Vis. Sci. 2021, 62, 25. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, Y.; Qian, J.; Wu, S.; Ji, Y.; Liu, Y.; Zeng, J.; Gong, A. Nattokinase Crude Extract Inhibits Hepatocellular Carcinoma Growth in Mice. J. Microbiol. Biotechnol. 2019, 29, 1281–1287. [Google Scholar] [CrossRef]
- Wu, H.; Wang, Y.; Zhang, Y.; Xu, F.; Chen, J.; Duan, L.; Zhang, T.; Wang, J.; Zhang, F. Breaking the vicious loop between inflammation, oxidative stress and coagulation, a novel anti-thrombus insight of nattokinase by inhibiting LPS-induced inflammation and oxidative stress. Redox Biol. 2020, 32, 101500. [Google Scholar] [CrossRef]
- Fujita, M.; Hong, K.; Ito, Y.; Fujii, R.; Kariya, K.; Nishimuro, S. Thrombolytic Effect of Nattokinase on a Chemically Induced Thrombosis Model in Rat. Biol. Pharm. Bull. 1995, 18, 1387–1391. [Google Scholar] [CrossRef]
- Pan, S.; Chen, G.; Zeng, J.; Cao, X.; Zheng, X.; Zeng, W.; Liang, Z. Fibrinolytic enzyme production from low-cost substrates by marine Bacillus subtilis: Process optimization and kinetic modeling. Biochem. Eng. J. 2019, 141, 268–277. [Google Scholar] [CrossRef]
- Devi, C.S.; Mohanasrinivasan, V.; Sharma, P.; Das, D.; Vaishnavi, B.; Naine, S.J. Production, Purification and Stability Studies on Nattokinase: A Therapeutic Protein Extracted from Mutant Pseudomonas aeruginosa CMSS Isolated from Bovine Milk. Int. J. Pept. Res. Ther. 2015, 22, 263–269. [Google Scholar] [CrossRef]
- Wang, S.-L.; Chen, H.-J.; Liang, T.-W.; Lin, Y.-D. A novel nattokinase produced by Pseudomonas sp. TKU015 using shrimp shells as substrate. Process Biochem. 2009, 44, 70–76. [Google Scholar] [CrossRef]
- Wang, C.; Du, M.; Zheng, D.; Kong, F.; Zu, G.; Feng, Y. Purification and Characterization of Nattokinase from Bacillus subtilis Natto B-12. J. Agric. Food Chem. 2009, 57, 9722–9729. [Google Scholar] [CrossRef]
- Zhang, X.; Tong, Y.; Wang, J.; Lyu, X.; Yang, R. Screening of a Bacillus subtilis strain producing both nattokinase and milk-clotting enzyme and its application in fermented milk with thrombolytic activity. J. Dairy Sci. 2021, 104, 9437–9449. [Google Scholar] [CrossRef]
- Peng, Y.; Huang, Q.; Zhang, R.-H.; Zhang, Y.-Z. Purification and characterization of a fibrinolytic enzyme produced by Bacillus amyloliquefaciens DC-4 screened from douchi, a traditional Chinese soybean food. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 134, 45–52. [Google Scholar] [CrossRef]
- Wang, S.H.; Zhang, C.; Yang, Y.L.; Diao, M.; Bai, M.F. Screening of a high fibrinolytic enzyme producing strain and characterization of the fibrinolytic enzyme produced from Bacillus subtilis LD-8547. World J. Microbiol. Biotechnol. 2007, 24, 475–482. [Google Scholar] [CrossRef]
- Wang, C.T.; Ji, B.P.; Li, B.; Nout, R.; Li, P.L.; Ji, H.; Chen, L.F. Purification and characterization of a fibrinolytic enzyme of Bacillus subtilis DC33, isolated from Chinese traditional Douchi. J. Ind. Microbiol. Biotechnol. 2006, 33, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Luo, M.; Xu, L.; Zhang, Y.; Lin, X.; Kong, P.; Liu, H. Production of Fibrinolytic Enzyme from Bacillus amyloliquefaciens by Fermentation of Chickpeas, with the Evaluation of the Anticoagulant and Antioxidant Properties of Chickpeas. J. Agric. Food Chem. 2011, 59, 3957–3963. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Choi, K.; Kim, Y.; Park, H.; Choi, J.; Lee, Y.; Oh, H.; Kwon, I.; Lee, S. Purification and characterization of a fibrinolytic enzyme produced from Bacillus sp. strain CK 11-4 screened from Chungkook-Jang. Appl. Environ. Microbiol. 1996, 62, 2482–2488. [Google Scholar] [CrossRef] [PubMed]
- Chang-Su, P.; Dong, H.K.; Woo-Yiel, L.; Dae-Ook, K.; Jae, J.S.; Nack-Shick, C. Identification of fibrinogen-induced nattokinase WRL101 from Bacillus subtilis WRL101 isolated from Doen-jang. Afr. J. Microbiol. Res. 2013, 7, 1983–1992. [Google Scholar] [CrossRef]
- Vaithilingam, M.; Chandrasekaran, S.D.; Gupta, S.; Paul, D.; Sahu, P.; Selvaraj, J.N.; Babu, V. Extraction of Nattokinase Enzyme from Bacillus cereus Isolated from Rust. Natl. Acad. Sci. Lett. 2016, 39, 263–267. [Google Scholar] [CrossRef]
- Kim, S.H.; Choi, N.S. Purification and Characterization of Subtilisin DJ-4 Secreted by Bacillus sp. Strain DJ-4 Screened from Doen-Jang. J. Agric. Chem. Soc. Jpn. 2000, 64, 1722–1725. [Google Scholar]
- Ko, J.H.; Yan, J.P.; Zhu, L.; Qi, Y.P. Identification of two novel fibrinolytic enzymes from Bacillus subtilis QK02. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 137, 65–74. [Google Scholar] [CrossRef]
- Kumar, D.M.; Rakshitha, R.; Vidhya, M.A.; Jennifer, P.S.; Prasad, S.; Kumar, M.R.; Kalaichelv, P. Production, Optimization and Characterization of Fibrinolytic Enzyme by Bacillus subtilis RJAS19. Pak. J. Biol. Sci. 2014, 17, 529–534. [Google Scholar] [CrossRef]
- Yin, L.-J.; Lin, H.-H.; Jiang, S.-T. Bioproperties of Potent Nattokinase from Bacillus subtilis YJ1. J. Agric. Food Chem. 2010, 58, 5737–5742. [Google Scholar] [CrossRef]
- Wang, S.-L.; Yeh, P.-Y. Production of a surfactant- and solvent-stable alkaliphilic protease by bioconversion of shrimp shell wastes fermented by Bacillus subtilis TKU007. Process Biochem. 2006, 41, 1545–1552. [Google Scholar] [CrossRef]
- Inatsu, Y.; Nakamura, N.; Yuriko, Y.; Fushimi, T.; Watanasiritum, L.; Kawamoto, S. Characterization of Bacillus subtilis strains in Thua nao, a traditional fermented soybean food in northern Thailand. Lett. Appl. Microbiol. 2006, 43, 237–242. [Google Scholar] [CrossRef]
- Wei, X.; Luo, M.; Xie, Y.; Yang, L.; Li, H.; Xu, L.; Liu, H. Strain Screening, Fermentation, Separation, and Encapsulation for Production of Nattokinase Functional Food. Appl. Biochem. Biotechnol. 2012, 168, 1753–1764. [Google Scholar] [CrossRef]
- Heo, S.; Kim, J.-H.; Kwak, M.-S.; Sung, M.-H.; Jeong, D.-W. Functional Annotation Genome Unravels Potential Probiotic Bacillus velezensis Strain KMU01 from Traditional Korean Fermented Kimchi. Foods 2021, 10, 563. [Google Scholar] [CrossRef]
- Keziah, S.M.; Devi, C.S. Fibrinolytic and ACE Inhibitory Activity of Nattokinase Extracted from Bacillus subtilis VITMS 2: A Strain Isolated from Fermented Milk of Vigna unguiculata. J. Protein Chem. 2021, 40, 876–890. [Google Scholar] [CrossRef]
- Syahbanu, F.; Giriwono, P.E.; Tjandrawinata, R.R.; Suhartono, M.T. Molecular analysis of a fibrin-degrading enzyme from Bacillus subtilis K2 isolated from the Indonesian soy-bean-based fermented food moromi. Mol. Biol. Rep. 2020, 47, 8553–8563. [Google Scholar] [CrossRef]
- Mahajan, P.M.; Nayak, S.; Lele, S.S. Fibrinolytic enzyme from newly isolated marine bacterium Bacillus subtilis ICTF-1: Media optimization, purification and characterization. J. Biosci. Bioeng. 2012, 113, 307–314. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamagata, Y.; Ichishima, E. Nucleotide Sequence of the Subtilisin NAT Gene, aprN, of Bacillus subtilis(natto). Biosci. Biotechnol. Biochem. 1992, 56, 1869–1871. [Google Scholar] [CrossRef]
- Kobayashi, T.; Hakamada, Y.; Adachi, S.; Hitomi, J.; Yoshimatsu, T.; Koike, K.; Kawai, S.; Ito, S. Purification and properties of an alkaline protease from alkalophilic Bacillus sp. KSM-K16. Appl. Micro-Biol. Biotechnol. 1995, 43, 473–481. [Google Scholar] [CrossRef]
- Song, X.; Yang, Y.; Liang, N.; Yang, F.; Chen, S.; Zhou, L.; Zhou, K.; Wang, Y. Quantitative pharmacokinetic evaluation of Subtilisin QK-2 after a bolus IV injection in a rat model using a novel sandwich enzyme-linked immunosorbent assay. J. Pharm. Biomed. Anal. 2020, 186, 113264. [Google Scholar] [CrossRef]
- Carter, P.; Wells, J.A. Dissecting the catalytic triad of a serine protease. Nature 1988, 332, 564–568. [Google Scholar] [CrossRef]
- Yanagisawa, Y.; Chatake, T.; Chiba-Kamoshida, K.; Naito, S.; Ohsugi, T.; Sumi, H.; Yasuda, I.; Morimoto, Y. Purification, crystallization and preliminary X-ray diffraction experiment of nattokinase from Bacillus subtilis natto. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66 Pt 12, 1670–1673. [Google Scholar] [CrossRef]
- Carter, P.; Wells, J.A. Functional interaction among catalytic residues in subtilisin BPN′. Proteins Struct. Funct. Bioinform. 1990, 7, 335–342. [Google Scholar] [CrossRef]
- Weng, M.; Zheng, Z.; Bao, W.; Cai, Y.; Yin, Y.; Zou, G. Enhancement of oxidative stability of the subtilisin nattokinase by site-directed mutagenesis expressed in Escherichia coli. Biochim. Biophys. Acta 2009, 1794, 1566–1572. [Google Scholar] [CrossRef]
- Yang, M.; Wu, J.; Huang, Q.; Jia, Y. Probing the Role of Catalytic Triad on the Cleavage between Intramolecular Chaperone and NK Mature Peptide. J. Agric. Food Chem. 2021, 69, 2348–2353. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, Z.; Li, S.; Tian, Z.; Ma, X. Study on the mechanism of production of gamma-PGA and nattokinase in Bacillus subtilis natto based on RNA-seq analysis. Microb. Cell Fact. 2021, 20, 83. [Google Scholar] [CrossRef] [PubMed]
- Pagnoncelli, M. Nattokinases, in Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 509–526. [Google Scholar]
- Tuan, N.A.; Thuan, D.T.H.; Tam, T.T.M.; Huong, N.T. Determination the optimum fermentation in obtaining nattokinase by Bacillus subtilis natto. Int. J. Innov. Appl. Stud. 2015, 13, 663. [Google Scholar]
- Berenjian, A.; Mahanama, R.; Kavanagh, J.; Dehghani, F.; Ghasemi, Y. Nattokinase production: Medium components and feeding strategy studies. Chem. Ind. Chem. Eng. Q. 2014, 20, 541–547. [Google Scholar] [CrossRef]
- Amin, K.; Zeng, X.; You, Y.; Hu, Y.; Sun, H.; Lyu, B.; Piao, C.; Yu, H. Enhanced thermostability and antioxidant activity of Nattokinase by biogenic enrichment of selenium. J. Food Meas. Charact. 2020, 14, 2145–2154. [Google Scholar] [CrossRef]
- Man, L.L.; Xiang, D.J.; Zhang, C.L. Strain Screening from Traditional Fermented Soybean Foods and Induction of Nattokinase Production in Bacillus subtilis MX-6. Probiotics Antimicrob. Proteins 2018, 11, 283–294. [Google Scholar] [CrossRef]
- Wu, R.; Chen, G.; Pan, S.; Zeng, J.; Liang, Z. Cost-effective fibrinolytic enzyme production by Bacillus subtilis WR350 using medium supplemented with corn steep powder and sucrose. Sci. Rep. 2019, 9, 6824. [Google Scholar] [CrossRef]
- Xiao, Z.; Shen, J.; Li, Y.; Wang, Z.; Zhao, Y.; Chen, Y.; Zhao, J.-Y. High and Economical Nattokinase Production with Acetoin as a Useful Byproduct from Soybean Milk and Glucose. Probiotics Antimicrob. Proteins. 2021. [Google Scholar] [CrossRef]
- Sahoo, A.; Mahanty, B.; Daverey, A.; Dutta, K. Nattokinase production from Bacillus subtilis using cheese whey: Effect of nitrogen supplementation and dynamic modelling. J. Water Process Eng. 2020, 38, 101533. [Google Scholar] [CrossRef]
- Li, T.; Zhan, C.; Guo, G.; Liu, Z.; Hao, N.; Ouyang, P. Tofu processing wastewater as a low-cost substrate for high activity nattokinase production using Bacillus subtilis. BMC Biotechnol. 2021, 21, 57. [Google Scholar] [CrossRef]
- Astrup, T.; Müllertz, S. The fibrin plate method for estimating fibrinolytic activity. Arch. Biochem. Biophys. 1952, 40, 346–351. [Google Scholar] [CrossRef]
- Sumi, H.; Nakajima, N.; Taya, N. The Method of Determination of the Thrombolytic Enzyme Nattokinase. J. Brew. Soc. Jpn. 1993, 88, 482–486. [Google Scholar] [CrossRef]
- Weng, M.; Deng, X.; Bao, W.; Zhu, L.; Wu, J.; Cai, Y.; Jia, Y.; Zheng, Z.; Zou, G. Improving the activity of the subtilisin nattokinase by site-directed mutagenesis and molecular dynamics simulation. Biochem. Biophys. Res. Commun. 2015, 465, 580–586. [Google Scholar] [CrossRef]
- Cheng, J.; Guan, C.; Cui, W.; Zhou, L.; Liu, Z.; Li, W.; Zhou, Z. Enhancement of a high efficient autoinducible expression system in Bacillus subtilis by promoter engineering. Protein Expr. Purif. 2016, 127, 81–87. [Google Scholar] [CrossRef]
- Xiao, J.; Peng, B.; Su, Z.; Liu, A.; Hu, Y.; Nomura, C.T.; Chen, S.; Wang, Q. Facilitating Protein Expression with Portable 5′-UTR Secondary Structures in Bacillus licheniformis. ACS Synth. Biol. 2020, 9, 1051–1058. [Google Scholar] [CrossRef]
- Hara, T.; Tadokoro, Y.; Satoyama, T. A Simple, Easy and Routine Assay of Thrombolytic Enzyme Activity. Nippon Shokuhin Kogyo Gakkai-Shi 1996, 43, 172–175. [Google Scholar] [CrossRef]
- Yuki, Y.; Nakagawa, T.; Fujita, M.; Asada, A.; Nakanishi, K.; Kato, K. A Sandwich Enzyme-Linked Immunosorbent Assay for Nattokinase. Biosci. Biotechnol. Biochem. 1994, 58, 366–370. [Google Scholar] [CrossRef]
- Wei, R.; Liu, Z.; Wen, J. Determination of extracellular fibrinolytic activity of nattokinase by ultraviolet spectrophotometry. China Brew. 2012, 31, 184–188. [Google Scholar]
- Zhang, X.; Lyu, X.; Tong, Y.; Wang, J.; Ye, J.; Yang, R. Chitosan/casein based microparticles with a bilayer shell–core structure for oral delivery of nattokinase. Food Funct. 2020, 11, 10799–10816. [Google Scholar] [CrossRef]
- Zhao, H. Modification of Thermal Stability and pH Stability of Nattokinase; Jiangnan University: Wuxi, China, 2018. [Google Scholar]
- Nguyen, T.T.; Quyen, T.D.; Le, H.T. Cloning and enhancing production of a detergent-and organic-solvent-resistant nat-tokinase from Bacillus subtilis VTCC-DVN-12-01 by using an eight-protease-gene-deficient Bacillus subtilis WB800. Microb. Cell Fact. 2013, 12, 79. [Google Scholar] [CrossRef]
- Yuan, J.; Yang, J.; Zhuang, Z.; Yang, Y.; Lin, L.; Wang, S. Thrombolytic effects of Douchi Fibrinolytic enzyme from Bacillus subtilis LD-8547 in vitro and in vivo. BMC Biotechnol. 2012, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.K.; Park, J.U.; Baek, H.; Park, S.H.; Kong, I.S.; Kim, D.W.; Joo, W.H. Purification and biochemical characterization of a fibrinolytic enzyme from Bacillus subtilis BK-17. World J. Microbiol. Biotechnol. 2001, 17, 89–92. [Google Scholar] [CrossRef]
- Chang, C.T.; Fan, M.H.; Kuo, F.C.; Sung, H.Y. Potent fibrinolytic enzyme from a mutant of Bacillus subtilis IMR-NK1. J. Agric. Food Chem. 2000, 48, 3210–3216. [Google Scholar] [CrossRef] [PubMed]
- Yongjun, C.; Wei, B.; Shujun, J.; Meizhi, W.; Yan, J.; Yan, Y.; Zhongliang, Z.; Goulin, Z. Directed evolution improves the fibrinolytic activity of nattokinase from Bacillus natto. FEMS Microbiol. Lett. 2011, 325, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Khowal, S.; Panda, B.P.; Wajid, S. Comparative genomic analyses of Bacillus subtilis strains to study the biochemical and molecular attributes of nattokinases. Biotechnol. Lett. 2022, 44, 485–502. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Feng, C.; Zhong, J.; Huan, L. Roles of S3 Site Residues of Nattokinase on its Activity and Substrate Specificity. J. Biochem. 2007, 142, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, H.; Han, L.; Cui, W.; Zhou, L.; Zhou, Z. Improvement of the acid resistance, catalytic efficiency, and thermostability of nattokinase by multisite-directed mutagenesis. Biotechnol. Bioeng. 2019, 116, 1833–1843. [Google Scholar] [CrossRef]
- Wei, X.; Zhou, Y.; Chen, J.; Cai, D.; Wang, D.; Qi, G.; Chen, S. Efficient expression of nattokinase in Bacillus licheniformis: Host strain construction and signal peptide optimization. J. Ind. Microbiol. Biotechnol. 2015, 42, 287–295. [Google Scholar] [CrossRef]
- Cai, D.; Wei, X.; Qiu, Y.; Chen, Y.; Chen, J.; Wen, Z.; Chen, S. High-level expression of nattokinase in Bacillus licheniformis by manipulating signal peptide and signal peptidase. J. Appl. Microbiol. 2016, 121, 704–712. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, W.; Ge, C.; Cui, W.; Zhou, L.; Zhou, Z. High-level extracellular production of recombinant nattokinase in Bacillus subtilis WB800 by multiple tandem promoters. BMC Microbiol. 2019, 19, 89. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, J.; Liu, C.; Li, J.; Li, Z.; Zhao, J.; Liu, H. Synthesis of sustained release/controlled release nanoparticles carrying nattokinase and their application in thrombolysis. Pharmazie 2021, 76, 145–149. [Google Scholar]
- Cockburn, D.W.; Vandenende, C.; Clarke, A.J. Modulating the pH− Activity Profile of Cellulase by Substitution: Replacing the General Base Catalyst Aspartate with Cysteinesulfinate in Cellulase A from Cellulomonas fimi. Biochemistry 2010, 49, 2042–2050. [Google Scholar] [CrossRef]
- Zhao, T.; Li, Y.; Yuan, S.; Ye, Y.; Peng, Z.; Zhou, R.; Liu, J. Structure-Based Design of Acetolactate Synthase from Bacillus licheniformis Improved Protein Stability under Acidic Conditions. Front. Microbiol. 2020, 11, 582909. [Google Scholar] [CrossRef]
- Ni, H.; Guo, P.-C.; Jiang, W.-L.; Fan, X.-M.; Luo, X.-Y.; Li, H.-H. Expression of nattokinase in Escherichia coli and renaturation of its inclusion body. J. Biotechnol. 2016, 231, 65–71. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Zhong, J.; Huan, L. Secretory expression of a heterologous nattokinase in Lactococcus lactis. Appl. Microbiol. Biotechnol. 2007, 75, 95–101. [Google Scholar] [CrossRef]
- Guangbo, Y.; Min, S.; Wei, S.; Lixin, M.; Chao, Z.; Yaping, W.; Zunxi, H. Heterologous expression of nattokinase from B. subtilis natto using Pichia pastoris GS115 and assessment of its thrombolytic activity. BMC Biotechnol. 2021, 21, 49. [Google Scholar] [CrossRef]
- Chiang, C.-J.; Chen, H.; Chao, Y.-P.; Tzen, J.T.C. Efficient System of Artificial Oil Bodies for Functional Expression and Purification of Recombinant Nattokinase in Escherichia coli. J. Agric. Food Chem. 2005, 53, 4799–4804. [Google Scholar] [CrossRef]
- Li, C.; Du, Z.; Qi, S.; Zhang, X.; Wang, M.; Zhou, Y.; Lu, H.; Gu, X.; Tian, H. Food-grade expression of nattokinase in Lactobacillus delbrueckii subsp. bulgaricus and its thrombolytic activity in vitro. Biotechnol. Lett. 2020, 42, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Sumi, H.; Hamada, H.; Nakanishi, K.; Hiratani, H. Enhancement of the Fibrinolytic Activity in Plasma by Oral Administration of Nattokinases. Acta Haematol. 1990, 84, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Yatagai, C.; Maruyama, M.; Kawahara, T.; Sumi, H. Nattokinase-Promoted Tissue Plasminogen Activator Release from Human Cells. Pathophysiol. Haemost. Thromb. 2008, 36, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Urano, T.; Ihara, H.; Umemura, K.; Suzuki, Y.; Oike, M.; Akita, S.; Tsukamoto, Y.; Suzuki, I.; Takada, A. The Profibrinolytic Enzyme Subtilisin NAT Purified from Bacillus subtilis Cleaves and Inactivates Plasminogen Activator Inhibitor Type 1. J. Biol. Chem. 2001, 276, 24690–24696. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kondo, K.; Matsumoto, Y.; Zhao, B.-Q.; Otsuguro, K.-I.; Maeda, T.; Tsukamoto, Y.; Urano, T.; Umemura, K. Dietary supplementation of fermented soybean, natto, suppresses intimal thickening and modulates the lysis of mural thrombi after endothelial injury in rat femoral artery. Life Sci. 2003, 73, 1289–1298. [Google Scholar] [CrossRef]
- Jang, J.-Y.; Kim, T.-S.; Cai, J.; Kim, J.; Kim, Y.; Shin, K.; Kim, K.S.; Park, S.K.; Lee, S.-P.; Choi, E.-K.; et al. Nattokinase improves blood flow by inhibiting platelet aggregation and thrombus formation. Lab. Anim. Res. 2013, 29, 221–225. [Google Scholar] [CrossRef]
- Xu, J.; Du, M.; Yang, X.; Chen, Q.; Chen, H.; Lin, D.-H. Thrombolytic Effects in vivo of Nattokinase in a Carrageenan-Induced Rat Model of Thrombosis. Acta Haematol. 2014, 132, 247–253. [Google Scholar] [CrossRef]
- Kamiya, S.; Hagimori, M.; Ogasawara, M.; Arakawa, M. In vivo Evaluation Method of the Effect of Nattokinase on Carrageenan-Induced Tail Thrombosis in a Rat Model. Acta Haematol. 2010, 124, 218–224. [Google Scholar] [CrossRef]
- Ren, N.N.; Chen, H.J.; Li, Y.; McGowan, E.; Lin, Y.G. A clinical study on the effect of nattokinase on carotid artery atherosclerosis and hyperlipidaemia. Zhonghua Yi Xue Za Zhi 2017, 97, 2038–2042. [Google Scholar] [CrossRef]
- Hodis, H.N.; Mack, W.J.; Meiselman, H.J.; Kalra, V.; Liebman, H.; Hwang-Levine, J.; Dustin, L.; Kono, N.; Mert, M.; Wenby, R.B.; et al. Nattokinase atherothrombotic prevention study: A randomized controlled trial. Clin. Hemorheol. Microcirc. 2021, 78, 339–353. [Google Scholar] [CrossRef]
- Ero, M.P.; Ng, C.M.; Mihailovski, T.; Harvey, N.R.; Lewis, B.H. A pilot study on the serum pharmacokinetics of nattokinase in humans following a single, oral, daily dose. Altern. Ther. Health Med. 2013, 19, 16–19. [Google Scholar]
- Kurosawa, Y.; Nirengi, S.; Homma, T.; Esaki, K.; Ohta, M.; Clark, J.F.; Hamaoka, T. A single-dose of oral nattokinase potentiates thrombolysis and anti-coagulation profiles. Sci. Rep. 2015, 5, 11601. [Google Scholar] [CrossRef]
- Dabbagh, F.; Negahdaripour, M.; Berenjian, A.; Behfar, A.; Mohammadi, F.; Zamani, M.; Irajie, C.; Ghasemi, Y. Nattokinase: Production and application. Appl. Microbiol. Biotechnol. 2014, 98, 9199–9206. [Google Scholar] [CrossRef]
- Milner, M.; Makise, K. Natto and Its Active Ingredient Nattokinase: A Potent and Safe Thrombolytic Agent. Altern. Complement. Ther. 2002, 8, 157–164. [Google Scholar] [CrossRef]
- Awatani-Yoshidome, K.; Hashimoto, T.; Satoh, T. Anaphylaxis from nattokinase in a patient with fermented soybean (natto) allergy. Allergol. Int. 2022, 71, 153–154. [Google Scholar] [CrossRef]
- Suzuki, S.; Nakamura, Y.; Kawano, Y.; Nishioka, K. A case of “late-onset” anaphylaxis caused by fermented soybeans; Natto. Arerugi 2006, 55, 832–836. [Google Scholar]
- Inomata, N.; Osuna, H.; Kawano, K.; Yamaguchi, J.; Yanagimachi, M.; Matsukura, S.; Ikezawa, Z. Late-onset anaphylaxis after ingestion of Bacillus subtilis-fermented soybeans (Natto): Clinical review of 7 patients. Allergol. Int. 2007, 56, 257–261. [Google Scholar] [CrossRef]
- Esteras, R.; Perez-Gomez, M.V.; Rodriguez-Osorio, L.; Ortiz, A.; Fernandez-Fernandez, B. Combination use of medicines from two classes of renin–angiotensin system blocking agents: Risk of hyperkalemia, hypotension, and impaired renal function. Ther. Adv. Drug Saf. 2015, 6, 166–176. [Google Scholar] [CrossRef]
- Fujita, M.; Ohnishi, K.; Takaoka, S.; Ogasawara, K.; Fukuyama, R.; Nakamuta, H. Antihypertensive effects of continuous oral administration of nattokinase and its fragments in spontaneously hypertensive rats. Biol. Pharm. Bull. 2011, 34, 1696–1701. [Google Scholar] [CrossRef]
- Kim, J.Y.; Gum, S.N.; Paik, J.K.; Lim, H.H.; Kim, K.-C.; Ogasawara, K.; Inoue, K.; Park, S.; Jang, Y.; Lee, J.H. Effects of Nattokinase on Blood Pressure: A Randomized, Controlled Trial. Hypertens. Res. 2008, 31, 1583–1588. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Yamanaka, N.; Ohnishi, K.; Fukayama, M.; Yoshino, M. Inhibition of angiotensin I converting enzyme by subtilisin NAT (nattokinase) in natto, a Japanese traditional fermented food. Food Funct. 2012, 3, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.C.; Pathak, S.; Kumar, V.; Panda, B.P. Attenuation of neurobehavioral and neurochemical abnormalities in animal model of cognitive deficits of Alzheimer’s disease by fermented soybean nanonutraceutical. Inflammopharmacology 2018, 26, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-J.; Kang, J.-I.; Kim, T.-S.; Yeo, I.-H. The Antithrombotic and Fibrinolytic Effect of Natto in Hypercholesterolemia Rats. Prev. Nutr. Food Sci. 2012, 17, 78–82. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, Y.; Yuan, T.; Yang, M.; Fang, W.; Li, L.; Fei, F.; Gong, A. Nattokinase crude extract enhances oral mucositis healing. BMC Oral Health 2021, 21, 555. [Google Scholar] [CrossRef]
- Walshe, L.J.; Malak, S.F.; Eagan, J.; Sepkowitz, K.A. Complication Rates among Cancer Patients with Peripherally Inserted Central Catheters. J. Clin. Oncol. 2002, 20, 3276–3281. [Google Scholar] [CrossRef]
- Menter, D.G.; Tucker, S.C.; Kopetz, S.; Sood, A.K.; Crissman, J.D.; Honn, K.V. Platelets and cancer: A casual or causal relationship: Revisited. Cancer Metastasis Rev. 2014, 33, 231–269. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, T.; Peng, B.; Luo, X.; Liu, X.; Hu, L.; Deng, Y. Targeted delivery of epirubicin to tumor-associated macrophages by sialic acid-cholesterol conjugate modified liposomes with improved antitumor activity. Int. J. Pharm. 2017, 523, 203–216. [Google Scholar] [CrossRef]
- Matsumoto, J.; Kojima, T.; Hiraguchi, E.; Abe, M. Portal vein tumor thrombus from colorectal cancer with no definite metastatic nodules in liver parenchyma. J. Hepato-Biliary-Pancreat. Surg. 2009, 16, 688–691. [Google Scholar] [CrossRef]
- Huang, M.; Ji, Y.; Yan, J.; Qi, T.; Zhang, S.-F.; Li, T.; Lü, S.; Liu, Y.; Liu, M. A nano polymer conjugate for dual drugs sequential release and combined treatment of colon cancer and thrombotic complications. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 110, 110697. [Google Scholar] [CrossRef]
- Wu, C.; Gao, C.; Lü, S.; Xu, X.; Wen, N.; Zhang, S.; Liu, M. Construction of polylysine dendrimer nanocomposites carrying nattokinase and their application in thrombolysis. J. Biomed. Mater. Res. Part A 2018, 106, 440–449. [Google Scholar] [CrossRef]
- Kou, Y.; Feng, R.; Chen, J.; Duan, L.; Wang, S.; Hu, Y.; Zhang, N.; Wang, T.; Deng, Y.; Song, Y. Development of a nattokinase–polysialic acid complex for advanced tumor treatment. Eur. J. Pharm. Sci. 2020, 145, 105241. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.-L.; Lee, K.-T.; Wang, J.-H.; Lee, L.Y.-L.; Chen, R.P.-Y. Amyloid-Degrading Ability of Nattokinase from Bacillus subtilis Natto. J. Agric. Food Chem. 2009, 57, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Fadl, N.N.; Ahmed, H.H.; Booles, H.F.; Sayed, A.H. Serrapeptase and nattokinase intervention for relieving Alzheimer’s disease pathophysiology in rat model. Hum. Exp. Toxicol. 2013, 32, 721–735. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.C.; Verma, A.; Al-Abbasi, F.A.; Anwar, F.; Kumar, V.; Panda, B.P. Development of surface-engineered PLGA nanoparticulate-delivery system of Tet1-conjugated nattokinase enzyme for inhibition of Abeta40 plaques in Alzheimer’s disease. Int. J. Nanomed. 2017, 12, 8749–8768. [Google Scholar] [CrossRef]
- Takano, A.; Hirata, A.; Ogasawara, K.; Sagara, N.; Inomata, Y.; Kawaji, T.; Tanihara, H. Posterior vitreous detachment induced by nattokinase (subtilisin NAT): A novel enzyme for pharmacologic vitreolysis. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2075–2079. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, L.; Liangqi, C.; Xiyu, T.; Jinyao, L. Biotechnology, Bioengineering and Applications of Bacillus Nattokinase. Biomolecules 2022, 12, 980. https://doi.org/10.3390/biom12070980
Yuan L, Liangqi C, Xiyu T, Jinyao L. Biotechnology, Bioengineering and Applications of Bacillus Nattokinase. Biomolecules. 2022; 12(7):980. https://doi.org/10.3390/biom12070980
Chicago/Turabian StyleYuan, Li, Chen Liangqi, Tang Xiyu, and Li Jinyao. 2022. "Biotechnology, Bioengineering and Applications of Bacillus Nattokinase" Biomolecules 12, no. 7: 980. https://doi.org/10.3390/biom12070980
APA StyleYuan, L., Liangqi, C., Xiyu, T., & Jinyao, L. (2022). Biotechnology, Bioengineering and Applications of Bacillus Nattokinase. Biomolecules, 12(7), 980. https://doi.org/10.3390/biom12070980