
Early Forms of α-Synuclein Pathology Are Associated with Neuronal Complex I Deficiency in the Substantia Nigra of Individuals with Parkinson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subject Cohort and Tissue Samples
2.2. Immunohistochemistry of Aggregated α-syn
2.3. Fluorescence Immunohistochemistry
2.4. Image Analysis
2.5. Statistical Analysis
3. Results
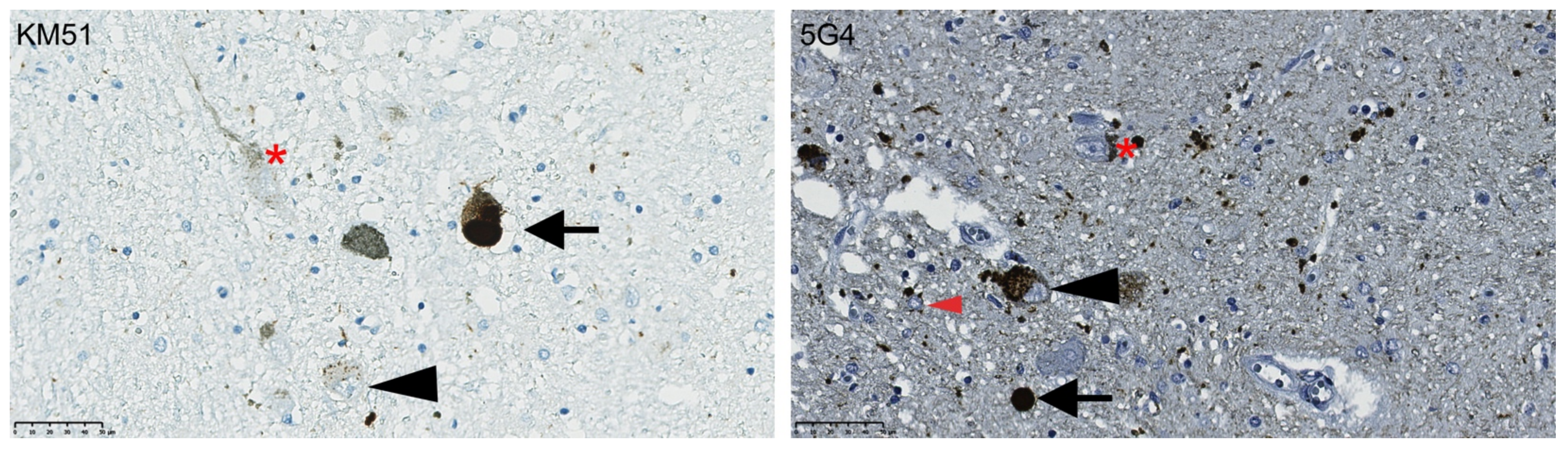
3.1. The 5G4 Antibody Detects Numerous Intraneuronal and Glial Immunopositive Structures
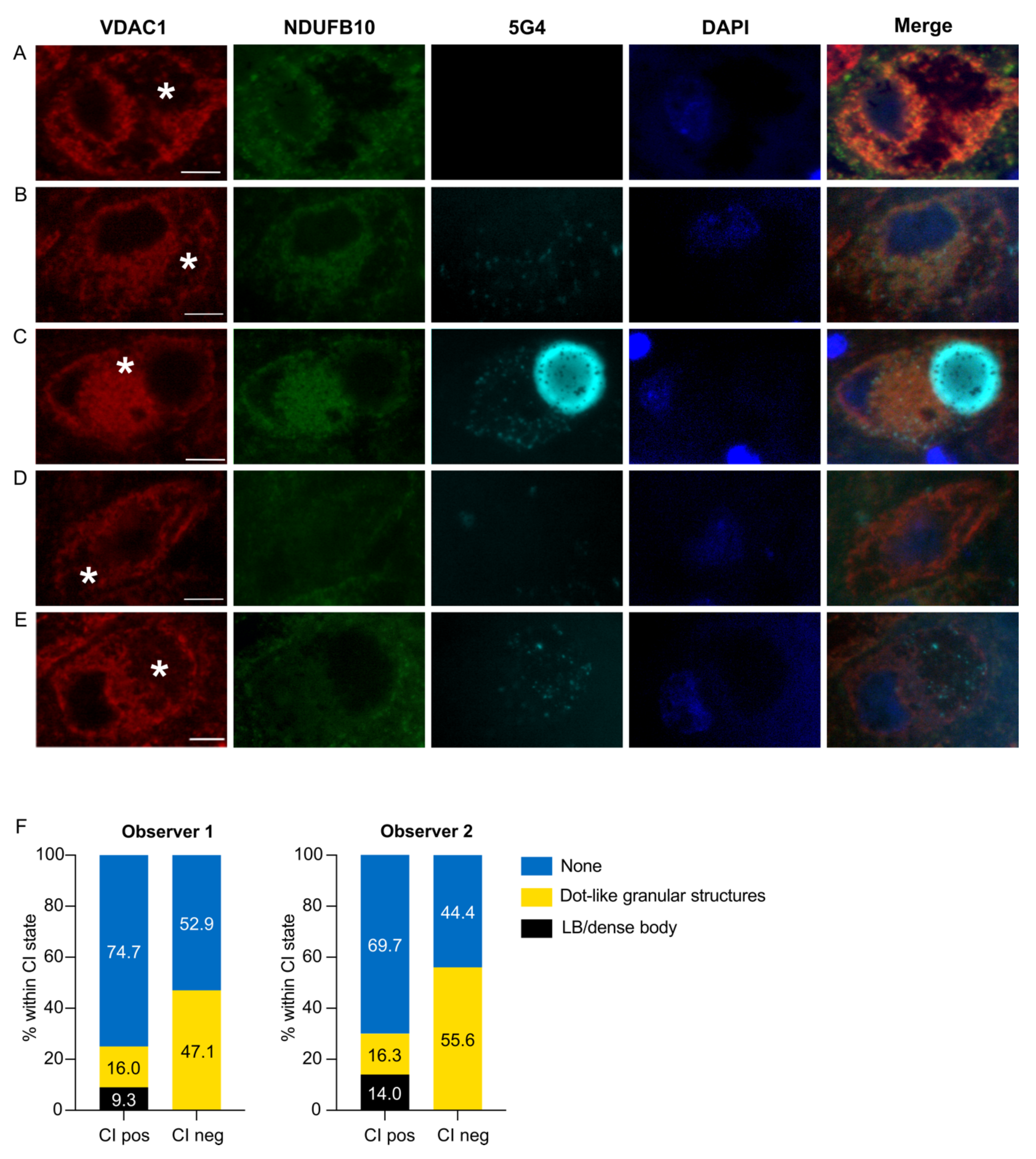
3.2. Early α-syn Pathology Has a Predilection for CI-Negative Dopaminergic SNc Neurons
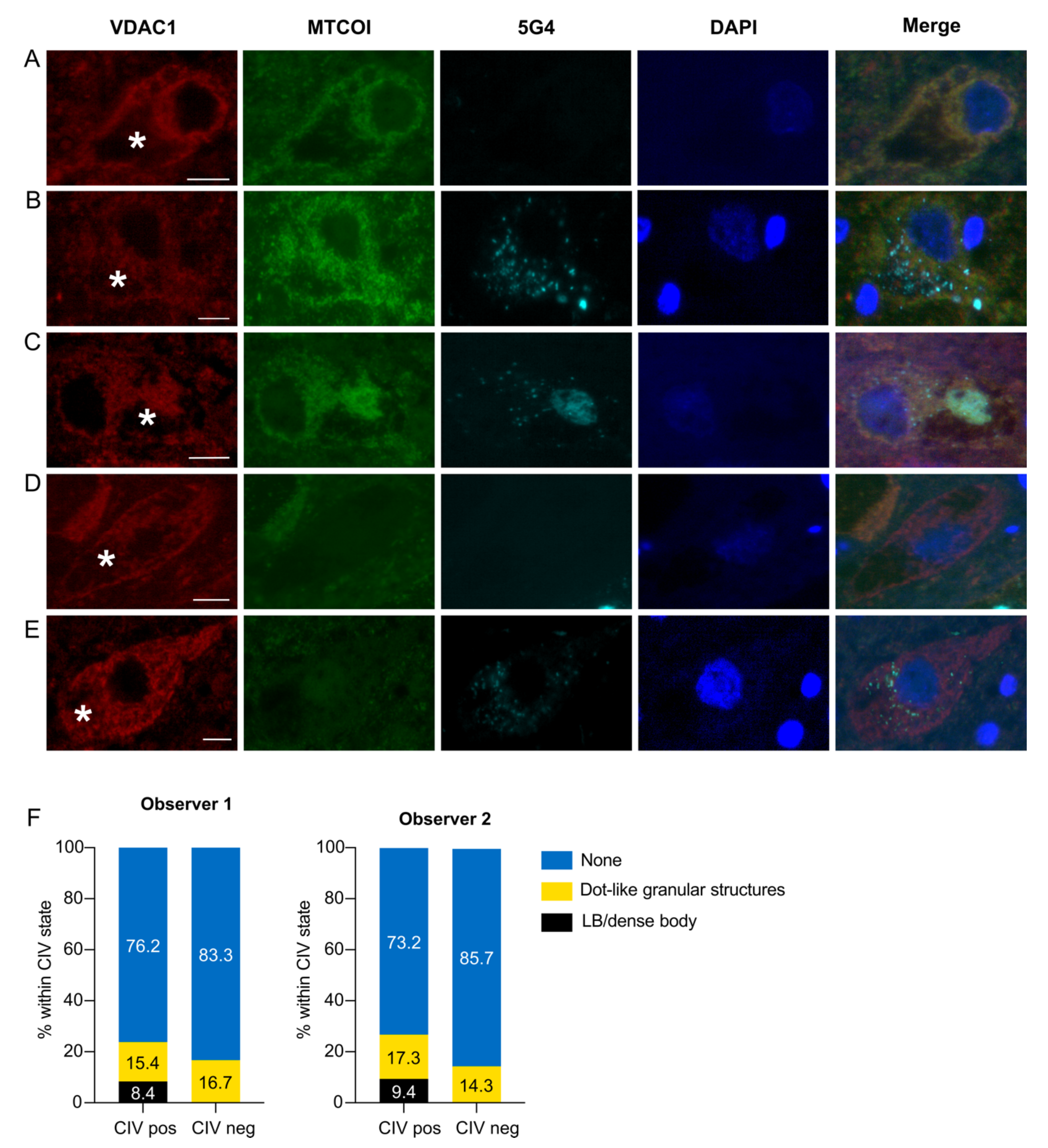
3.3. CIV Deficiency Is Not Associated with α-syn Pathology in Dopaminergic Neurons
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PD | Parkinson’s disease |
| iPD | idiopathic Parkinson’s disease |
| SNc | substantia nigra pars compacta |
| LP | Lewy pathology |
| -syn | -synuclein |
| LB | Lewy body |
| PB | pale body |
| mtDNA | mitochondrial DNA |
| MRC | mitochondrial respiratory chain |
| CI | complex I |
| ROS | reactive oxygen species |
| CIV | complex IV |
| PMI | post-mortem interval |
| IHC | Immunohistochemistry |
| MELAS | mitochondrial encephalomyopathy, lactic acidosis and stroke-like episodes |
References
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef]
- GBD 2016 Parkinson’s Disease Collaborators. Global, regional, and national burden of Parkinson’s disease, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Surmeier, D.J.; Obeso, J.A.; Halliday, G.M. Selective neuronal vulnerability in Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Dickson, D.W. Parkinson’s disease and parkinsonism: Neuropathology. Cold Spring Harb. Perspect. Med. 2012, 2, a009258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. Alpha-synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef]
- Fares, M.B.; Jagannath, S.; Lashuel, H.A. Reverse engineering Lewy bodies: How far have we come and how far can we go? Nat. Rev. Neurosci. 2021, 22, 111–131. [Google Scholar] [CrossRef]
- Gelpi, E.; Colom-Cadena, M. Oligomers: A hot topic for neurodegeneration and a note of caution for experimental models. Brain 2019, 142, 228–230. [Google Scholar] [CrossRef]
- Bender, A.; Krishnan, K.J.; Morris, C.M.; Taylor, G.A.; Reeve, A.K.; Perry, R.H.; Jaros, E.; Hersheson, J.S.; Betts, J.; Klopstock, T.; et al. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat. Genet. 2006, 38, 515–517. [Google Scholar] [CrossRef]
- Grunewald, A.; Rygiel, K.A.; Hepplewhite, P.D.; Morris, C.M.; Picard, M.; Turnbull, D.M. Mitochondrial DNA Depletion in Respiratory Chain-Deficient Parkinson Disease Neurons. Ann. Neurol. 2016, 79, 366–378. [Google Scholar] [CrossRef] [Green Version]
- Dölle, C.; Flones, I.; Nido, G.S.; Miletic, H.; Osuagwu, N.; Kristoffersen, S.; Lilleng, P.K.; Larsen, J.P.; Tysnes, O.B.; Haugarvoll, K.; et al. Defective mitochondrial DNA homeostasis in the substantia nigra in Parkinson disease. Nat. Commun. 2016, 7, 13548. [Google Scholar] [CrossRef]
- Subrahmanian, N.; LaVoie, M.J. Is there a special relationship between complex I activity and nigral neuronal loss in Parkinson’s disease? A critical reappraisal. Brain Res. 2021, 1767, 147434. [Google Scholar] [CrossRef] [PubMed]
- Ludtmann, M.H.R.; Angelova, P.R.; Horrocks, M.H.; Choi, M.L.; Rodrigues, M.; Baev, A.Y.; Berezhnov, A.V.; Yao, Z.; Little, D.; Banushi, B.; et al. α-synuclein oligomers interact with ATP synthase and open the permeability transition pore in Parkinson’s disease. Nat. Commun. 2018, 9, 2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeve, A.K.; Ludtmann, M.H.; Angelova, P.R.; Simcox, E.M.; Horrocks, M.H.; Klenerman, D.; Gandhi, S.; Turnbull, D.M.; Abramov, A.Y. Aggregated alpha-synuclein and complex I deficiency: Exploration of their relationship in differentiated neurons. Cell Death Dis. 2015, 6, e1820. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.H.; Fuentes, F.; Vanasco, V.; Alvarez, S.; Alaimo, A.; Cassina, A.; Coluccio Leskow, F.; Velazquez, F. Alpha-synuclein mitochondrial interaction leads to irreversible translocation and complex I impairment. Arch. Biochem. Biophys. 2018, 651, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Vicario, M.; Cieri, D.; Brini, M.; Calì, T. The Close Encounter Between Alpha-Synuclein and Mitochondria. Front. Neurosci. 2018, 12, 388. [Google Scholar] [CrossRef] [Green Version]
- Devi, L.; Raghavendran, V.; Prabhu, B.M.; Avadhani, N.G.; Anandatheerthavarada, H.K. Mitochondrial import and accumulation of alpha-synuclein impair complex I in human dopaminergic neuronal cultures and Parkinson disease brain. J. Biol. Chem. 2008, 283, 9089–9100. [Google Scholar] [CrossRef] [Green Version]
- Klivenyi, P.; Siwek, D.; Gardian, G.; Yang, L.; Starkov, A.; Cleren, C.; Ferrante, R.J.; Kowall, N.W.; Abeliovich, A.; Beal, M.F. Mice lacking alpha-synuclein are resistant to mitochondrial toxins. Neurobiol. Dis. 2006, 21, 541–548. [Google Scholar] [CrossRef]
- Song, D.D.; Shults, C.W.; Sisk, A.; Rockenstein, E.; Masliah, E. Enhanced substantia nigra mitochondrial pathology in human alpha-synuclein transgenic mice after treatment with MPTP. Exp. Neurol. 2004, 186, 158–172. [Google Scholar] [CrossRef]
- Reeve, A.K.; Park, T.K.; Jaros, E.; Campbell, G.R.; Lax, N.Z.; Hepplewhite, P.D.; Krishnan, K.J.; Elson, J.L.; Morris, C.M.; McKeith, I.G.; et al. Relationship between mitochondria and α-synuclein: A study of single substantia nigra neurons. Arch. Neurol. 2012, 69, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Flones, I.H.; Fernandez-Vizarra, E.; Lykouri, M.; Brakedal, B.; Skeie, G.O.; Miletic, H.; Lilleng, P.K.; Alves, G.; Tysnes, O.B.; Haugarvoll, K.; et al. Neuronal complex I deficiency occurs throughout the Parkinson’s disease brain, but is not associated with neurodegeneration or mitochondrial DNA damage. Acta Neuropathol. 2017, 135, 409–425. [Google Scholar] [CrossRef] [Green Version]
- Alves, G.; Muller, B.; Herlofson, K.; HogenEsch, I.; Telstad, W.; Aarsland, D.; Tysnes, O.B.; Larsen, J.P.; Norwegian ParkWest study, g. Incidence of Parkinson’s disease in Norway: The Norwegian ParkWest study. J. Neurol. Neurosurg. Psychiatry 2009, 80, 851–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaare, J.J.; Nido, G.S.; Sztromwasser, P.; Knappskog, P.M.; Dahl, O.; Lund-Johansen, M.; Maple-Grødem, J.; Alves, G.; Tysnes, O.B.; Johansson, S.; et al. Rare genetic variation in mitochondrial pathways influences the risk for Parkinson’s disease. Mov. Disord. 2018, 33, 1591–1600. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Del Tredici, K.; Rub, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Braak, H.; Alafuzoff, I.; Arzberger, T.; Kretzschmar, H.; Del Tredici, K. Staging of Alzheimer disease-associated neurofibrillary pathology using paraffin sections and immunocytochemistry. Acta Neuropathol. 2006, 112, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Thal, D.R.; Rüb, U.; Orantes, M.; Braak, H. Phases of A beta-deposition in the human brain and its relevance for the development of AD. Neurology 2002, 58, 1791–1800. [Google Scholar] [CrossRef]
- Kumar, S.T.; Jagannath, S.; Francois, C.; Vanderstichele, H.; Stoops, E.; Lashuel, H.A. How specific are the conformation-specific α-synuclein antibodies? Characterization and validation of 16 α-synuclein conformation-specific antibodies using well-characterized preparations of α-synuclein monomers, fibrils and oligomers with distinct structures and morphology. Neurobiol. Dis. 2020, 146, 105086. [Google Scholar] [CrossRef]
- Kovacs, G.G.; Wagner, U.; Dumont, B.; Pikkarainen, M.; Osman, A.A.; Streichenberger, N.; Leisser, I.; Verchère, J.; Baron, T.; Alafuzoff, I.; et al. An antibody with high reactivity for disease-associated α-synuclein reveals extensive brain pathology. Acta Neuropathol. 2012, 124, 37–50. [Google Scholar] [CrossRef]
- Kovacs, G.G.; Breydo, L.; Green, R.; Kis, V.; Puska, G.; Lőrincz, P.; Perju-Dumbrava, L.; Giera, R.; Pirker, W.; Lutz, M.; et al. Intracellular processing of disease-associated α-synuclein in the human brain suggests prion-like cell-to-cell spread. Neurobiol. Dis. 2014, 69, 76–92. [Google Scholar] [CrossRef]
- Cicchetti, D.V. Guidelines, Criteria, and Rules of Thumb for Evaluating Normed and Standardized Assessment Instruments in Psychology. Psychol. Assess. 1994, 6, 284–290. [Google Scholar] [CrossRef]
- Distelmaier, F.; Koopman, W.J.; van den Heuvel, L.P.; Rodenburg, R.J.; Mayatepek, E.; Willems, P.H.; Smeitink, J.A. Mitochondrial complex I deficiency: From organelle dysfunction to clinical disease. Brain 2009, 132, 833–842. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.W.; Li, X.; Thompson, D.; Wooding, K.; Chang, T.L.; Tang, Z.; Yu, H.; Thomas, P.J.; DeMartino, G.N. ATP binding and ATP hydrolysis play distinct roles in the function of 26S proteasome. Mol. Cell 2006, 24, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mindell, J.A. Lysosomal acidification mechanisms. Annu. Rev. Physiol. 2012, 74, 69–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, T.; Tanaka, K. Dissecting various ATP-dependent steps involved in proteasomal degradation. Mol. Cell 2003, 11, 3–5. [Google Scholar] [CrossRef]
- Scudamore, O.; Ciossek, T. Increased Oxidative Stress Exacerbates α-Synuclein Aggregation In Vivo. J. Neuropathol. Exp. Neurol. 2018, 77, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Musgrove, R.E.; Helwig, M.; Bae, E.J.; Aboutalebi, H.; Lee, S.J.; Ulusoy, A.; Di Monte, D.A. Oxidative stress in vagal neurons promotes parkinsonian pathology and intercellular α-synuclein transfer. J. Clin. Investig. 2019, 129, 3738–3753. [Google Scholar] [CrossRef]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar] [CrossRef]
- Sherer, T.B.; Kim, J.H.; Betarbet, R.; Greenamyre, J.T. Subcutaneous rotenone exposure causes highly selective dopaminergic degeneration and alpha-synuclein aggregation. Exp. Neurol. 2003, 179, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Erskine, D.; Reeve, A.K.; Polvikoski, T.; Schaefer, A.M.; Taylor, R.W.; Lax, N.Z.; El-Agnaf, O.; Attems, J.; Gorman, G.S.; Turnbull, D.M.; et al. Lewy body pathology is more prevalent in older individuals with mitochondrial disease than controls. Acta Neuropathol. 2020, 139, 219–221. [Google Scholar] [CrossRef] [Green Version]
- Tzoulis, C.; Tran, G.T.; Coxhead, J.; Bertelsen, B.; Lilleng, P.K.; Balafkan, N.; Payne, B.; Miletic, H.; Chinnery, P.F.; Bindoff, L.A. Molecular pathogenesis of polymerase gamma-related neurodegeneration. Ann. Neurol. 2014, 76, 66–81. [Google Scholar] [CrossRef] [Green Version]
- Chrysostomou, A.; Grady, J.P.; Laude, A.; Taylor, R.W.; Turnbull, D.M.; Lax, N.Z. Investigating complex I deficiency in Purkinje cells and synapses in patients with mitochondrial disease. Neuropathol. Appl. Neurobiol. 2016, 42, 477–492. [Google Scholar] [CrossRef] [Green Version]
- González-Rodríguez, P.; Zampese, E.; Stout, K.A.; Guzman, J.N.; Ilijic, E.; Yang, B.; Tkatch, T.; Stavarache, M.A.; Wokosin, D.L.; Gao, L.; et al. Disruption of mitochondrial complex I induces progressive parkinsonism. Nature 2021, 599, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Tzoulis, C.; Tran, G.T.; Schwarzlmuller, T.; Specht, K.; Haugarvoll, K.; Balafkan, N.; Lilleng, P.K.; Miletic, H.; Biermann, M.; Bindoff, L.A. Severe nigrostriatal degeneration without clinical parkinsonism in patients with polymerase gamma mutations. Brain 2013, 136, 2393–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langston, J.W.; Ballard, P.; Tetrud, J.W.; Irwin, I. Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science 1983, 219, 979–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, Y.C.; Krainc, D. α-synuclein toxicity in neurodegeneration: Mechanism and therapeutic strategies. Nat. Med. 2017, 23, 1–13. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Study ID | Diagnosis | AOD | DD | Sex | PMI | BRAAK Syn | Braak Tau | Thal Phase |
|---|---|---|---|---|---|---|---|---|
| PD 01 | PD | 88 | 11 | M | 30 | 6 | III | 1 |
| PD 02 | PD | 82 | 12 | M | 46 | 6 | I | 2 |
| PD 03 | PD | 78 | 12 | F | 24 | 6 | III | 3 |
| PD 04 | PD | 90 | 10 | F | 24 | 6 | III | 0 |
| PD 05 | PD | 72 | 10 | F | 72 | 6 | II | 0 |
| PD 06 | PD | 75 | 12 | M | 11 | 6 | I | 0 |
| PD 07 | PD | 94 | 14 | M | 36 | 6 | IV | 4 |
| PD 08 | PD | 85 | 14 | F | 23 | 6 | III | N/A |
| Ctrl 01 | Ctrl | 79 | - | M | 48 | 0 | I | 2 |
| Ctrl 02 | Ctrl | 96 | - | M | 24 | 0 | II | 3 |
| Ctrl 03 | Ctrl | 78 | - | F | 43 | 0 | III | 4 |
| Ctrl 04 | Ctrl | 95 | - | F | 30 | 0 | III | 4 |
| Ctrl 05 | Ctrl | 95 | - | F | 30 | 0 | III | 3 |
| Cytoplasmic α-syn Inclusion | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LB/PB | Punctate | No Inclusion | ||||||||
| O1 | O2 | O1 | O2 | O1 | O2 | O1 | O2 | |||
| CI state | CI+ | Count | 23 | 18 | 28 | 31 | 125 | 146 | 176 | 195 |
| Expected count | 20.9 | 16.6 | 34.5 | 35.9 | 120.7 | 142.6 | ||||
| % Within CI state | 14.0 | 9.2 | 15.9 | 15.9 | 71.0 | 74.9 | ||||
| Adjusted residual | 1.6 | 1.3 | −4.0 | −3.2 | 2.3 | 2.0 | ||||
| CI− | Count | 0 | 0 | 10 | 8 | 8 | 9 | 18 | 17 | |
| Expected count | 2.1 | 1.4 | 3.5 | 3.1 | 12.3 | 12.4 | ||||
| % Within CI state | 0.0 | 0.0 | 55.6 | 47.1 | 44.4 | 52.9 | ||||
| Adjusted residual | −1.6 | −1.3 | 4.0 | 3.2 | −2.3 | −2.0 | ||||
| p-value | 0.11 | 0.19 | 6.3 × 10−5 | 1.3 × 10−3 | 0.021 | 0.046 | ||||
| Total | 23 | 18 | 38 | 39 | 133 | 155 | 194 | 212 | ||
| Cytoplasmic α-syn Inclusion | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| LB/PB | Punctate | No Inclusion | ||||||||
| O1 | O2 | O1 | O2 | O1 | O2 | O1 | O2 | |||
| CIV state | CIV+ | Count | 12 | 12 | 22 | 22 | 109 | 93 | 143 | 127 |
| Expected count | 10.3 | 10.8 | 22.3 | 21.6 | 110.5 | 94.6 | ||||
| % Within CIV state | 8.4 | 9.4 | 15.4 | 17.3 | 76.2 | 73.2 | ||||
| Adjusted residual | 1.5 | 1.2 | −0.2 | 0.3 | −0.8 | −1.0 | ||||
| CIV− | Count | 0 | 0 | 4 | 2 | 20 | 12 | 24 | 14 | |
| Expected count | 1.7 | 1.2 | 3.7 | 2.4 | 18.5 | 10.4 | ||||
| % Within CIV state | 0.0 | 0.0 | 16.7 | 14.3 | 83.3 | 85.7 | ||||
| Adjusted residual | −1.5 | −1.2 | 0.2 | −0.3 | 0.8 | 1.0 | ||||
| p-value | 0.13 | 0.23 | 0.84 | 0.76 | 0.42 | 0.32 | ||||
| Total | 12 | 12 | 26 | 24 | 129 | 105 | 167 | 141 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flønes, I.H.; Nyland, H.; Sandnes, D.-A.; Alves, G.W.; Tysnes, O.-B.; Tzoulis, C. Early Forms of α-Synuclein Pathology Are Associated with Neuronal Complex I Deficiency in the Substantia Nigra of Individuals with Parkinson’s Disease. Biomolecules 2022, 12, 747. https://doi.org/10.3390/biom12060747
Flønes IH, Nyland H, Sandnes D-A, Alves GW, Tysnes O-B, Tzoulis C. Early Forms of α-Synuclein Pathology Are Associated with Neuronal Complex I Deficiency in the Substantia Nigra of Individuals with Parkinson’s Disease. Biomolecules. 2022; 12(6):747. https://doi.org/10.3390/biom12060747
Chicago/Turabian StyleFlønes, Irene Hana, Harald Nyland, Dagny-Ann Sandnes, Guido Werner Alves, Ole-Bjørn Tysnes, and Charalampos Tzoulis. 2022. "Early Forms of α-Synuclein Pathology Are Associated with Neuronal Complex I Deficiency in the Substantia Nigra of Individuals with Parkinson’s Disease" Biomolecules 12, no. 6: 747. https://doi.org/10.3390/biom12060747
APA StyleFlønes, I. H., Nyland, H., Sandnes, D.-A., Alves, G. W., Tysnes, O.-B., & Tzoulis, C. (2022). Early Forms of α-Synuclein Pathology Are Associated with Neuronal Complex I Deficiency in the Substantia Nigra of Individuals with Parkinson’s Disease. Biomolecules, 12(6), 747. https://doi.org/10.3390/biom12060747