
Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma


Abstract
1. Introduction
2. Development of Anti-GD2 Antibody for Therapeutic Use
3. Different Anti-GD2 Preparations and Their Pros and Cons
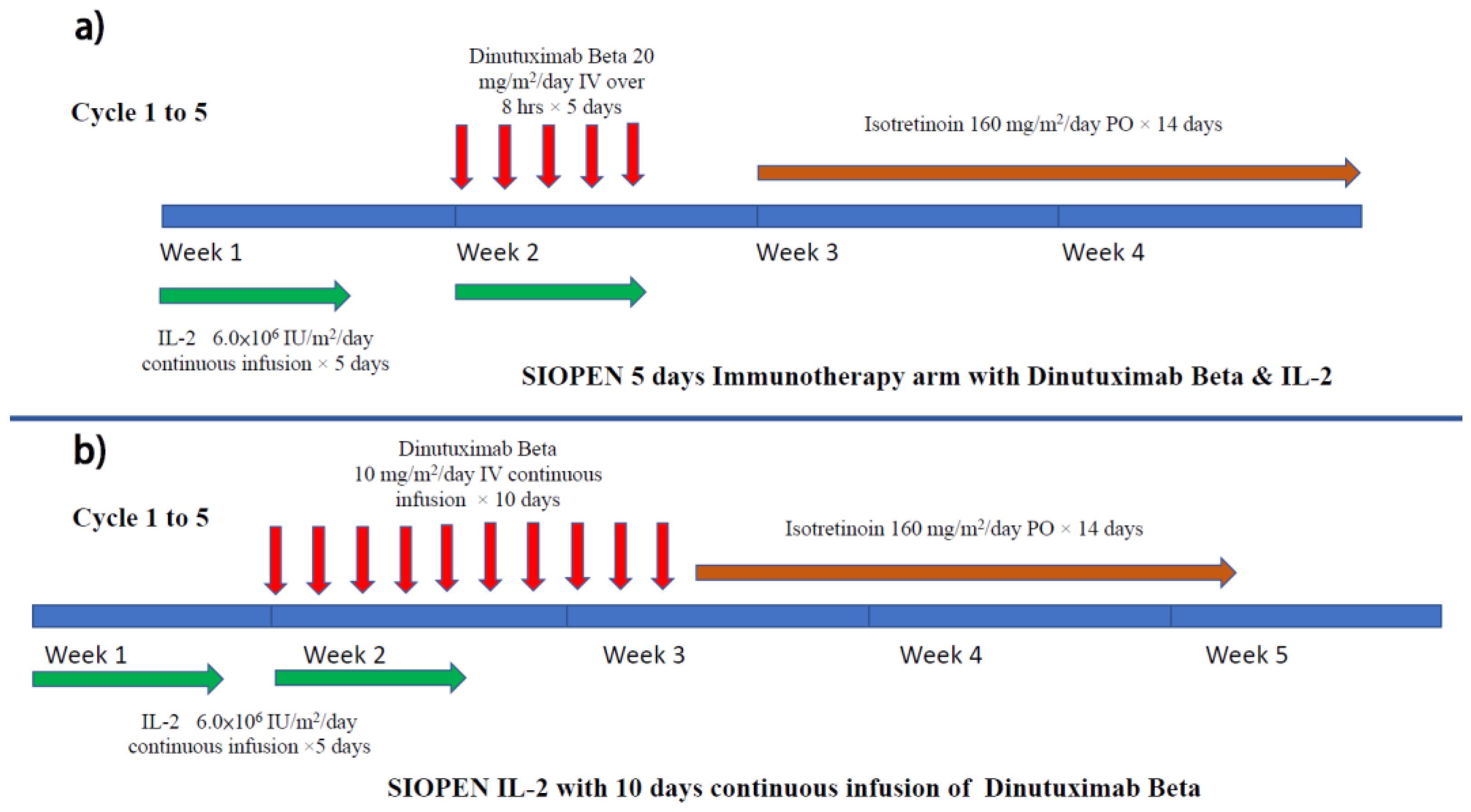
4. Clinical Data on Anti-GD2 for Newly Diagnosed Patients
5. Clinical Data on Anti-GD2 for Relapsed Patients
6. Side Effects of Anti-GD2
7. Future Prospective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mertens, R.; Granzen, B.; Lassay, L.; Bucsky, P.; Hundgen, M.; Stetter, G.; Heimann, G.; Weiss, C.; Hess, C.F.; Gademann, G.; et al. Treatment of nasopharyngeal carcinoma in children and adolescents: Definitive results of a multicenter study (NPC-91-GPOH). Cancer 2005, 104, 1083–1089. [Google Scholar] [CrossRef]
- El-Mallawany, N.K.; Cairo, M.S. Advances in the diagnosis and treatment of childhood and adolescent B-cell non-Hodgkin lymphoma. Clin. Adv. Hematol. Oncol. H&O 2015, 13, 113–123. [Google Scholar]
- Massimino, M.; Bode, U.; Biassoni, V.; Fleischhack, G. Nimotuzumab for pediatric diffuse intrinsic pontine gliomas. Expert Opin. Biol. Ther. 2010, 11, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Baroni, L.V.; Alderete, D.; Solano-Paez, P.; Rugilo, C.; Freytes, C.; Laughlin, S.; Fonseca, A.; Bartels, U.; Tabori, U.; Bouffet, E.; et al. Bevacizumab for pediatric radiation necrosis. Neuro-Oncol. Pract. 2020, 7, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.K.V.; Cheung, I.Y.; Kushner, B.H.; Ostrovnaya, I.; Chamberlain, E.; Kramer, K.; Modak, S. Murine Anti-GD2 Monoclonal Antibody 3F8 Combined with Granulocyte-Macrophage Colony-Stimulating Factor and 13-Cis-Retinoic Acid in High-Risk Patients with Stage 4 Neuroblastoma in First Remission. J. Clin. Oncol. 2012, 30, 3264–3270. [Google Scholar] [CrossRef]
- Hongeng, S.; Pctviscs, S.; Worapongpaiboon, S.; Rcrkamnuaychoke, B.; Pakakasama, S.; Jootar, S. Generation of CD3+ CD56+ Cytokine-Induced Killer Cells and Their In Vitro Cytotoxicity against Pediatric Cancer Cells. Int. J. Hematol. 2003, 77, 175–179. [Google Scholar] [CrossRef]
- De Bruijn, S.; Anguille, S.; Verlooy, J.; Smits, E.L.; van Tendeloo, V.F.; de Laere, M.; Norga, K.; Berneman, Z.N.; Lion, E. Dendritic Cell-Based and Other Vaccination Strategies for Pediatric Cancer. Cancers 2019, 11, 1396. [Google Scholar] [CrossRef]
- Nguyen, R.; Wu, H.; Pounds, S.; Inaba, H.; Ribeiro, R.C.; Cullins, D.; Rooney, B.; Bell, T.; Lacayo, N.J.; Heym, K.; et al. A phase II clinical trial of adoptive transfer of haploidentical natural killer cells for consolidation therapy of pediatric acute myeloid leukemia. J. Immunother. Cancer 2019, 7, 81. [Google Scholar] [CrossRef]
- Myers, R.M.; Dolan, J.; Teachey, D.T. Chimeric antigen receptor T cell therapy for pediatric and young adult B cell acute lymphoblastic leukemia. Expert Rev. Clin. Immunol. 2020, 16, 1029–1042. [Google Scholar] [CrossRef]
- Kabir, T.; Chauhan, A.; Anthony, L.; Hildebrandt, G.C. Immune Checkpoint Inhibitors in Pediatric Solid Tumors: Status in 2018. Ochsner J. 2018, 18, 370–376. [Google Scholar] [CrossRef]
- Friedman, G.K.; Johnston, J.M.; Bag, A.K.; Bernstock, J.D.; Li, R.; Aban, I.; Kachurak, K.; Nan, L.; Kang, K.-D.; Totsch, S.; et al. Oncolytic HSV-1 G207 Immunovirotherapy for Pediatric High-Grade Gliomas. N. Engl. J. Med. 2021, 384, 1613–1622. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.-L.; Yu, B.; Ning, B.-T.; Chan, S.; Lin, Q.-B.; Li, J.C.-B.; Huang, J.-D.; Chan, G.C.-F. Genetically modified “obligate” anaerobic Salmonella typhimurium as a therapeutic strategy for neuroblastoma. J. Hematol. Oncol. 2015, 8, 99. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Majzner, R.G.; Heitzeneder, S.; Mackall, C.L. Harnessing the Immunotherapy Revolution for the Treatment of Childhood Cancers. Cancer Cell 2017, 31, 476–485. [Google Scholar] [CrossRef]
- Irwin, M.S.; Naranjo, A.; Zhang, F.F.; Cohn, S.L.; London, W.B.; Gastier-Foster, J.M.; Ramirez, N.C.; Pfau, R.; Reshmi, S.; Wagner, E.; et al. Revised Neuroblastoma Risk Classification System: A Report from the Children’s Oncology Group. J. Clin. Oncol. 2021, 39, 3229–3241. [Google Scholar] [CrossRef] [PubMed]
- Katano, M.; Irie, R.F. Human monoclonal antibody to tumor-associated ganglioside GD2: Suppressed growth of human melanoma in nude mice. Immunol. Lett. 1984, 8, 169–174. [Google Scholar] [CrossRef]
- Cheung, N.K.; Saarinen, U.M.; Neely, J.E.; Landmeier, B.; Donovan, D.; Coccia, P.F. Monoclonal antibodies to a glycolipid antigen on human neuroblastoma cells. Cancer Res. 1985, 45, 2642–2649. [Google Scholar] [PubMed]
- Yu, A.L.; Gilman, A.L.; Ozkaynak, M.F.; London, W.B.; Kreissman, S.G.; Chen, H.X.; Smith, M.; Anderson, B.; Villablanca, J.G.; Matthay, K.K.; et al. Anti-GD2 Antibody with GM-CSF, Interleukin-2, and Isotretinoin for Neuroblastoma. N. Engl. J. Med. 2010, 363, 1324–1334. [Google Scholar] [CrossRef]
- Ladenstein, R.; Pötschger, U.; Valteau-Couanet, D.; Luksch, R.; Castel, V.; Ash, S.; Laureys, G.; Brock, P.; Michon, J.M.; Owens, C.; et al. Investigation of the Role of Dinutuximab Beta-Based Immunotherapy in the SIOPEN High-Risk Neuroblastoma 1 Trial (HR-NBL1). Cancers 2020, 12, 309. [Google Scholar] [CrossRef]
- Ahmed, M.; Cheung, N.-K.V. Engineering anti-GD2 monoclonal antibodies for cancer immunotherapy. FEBS Lett. 2013, 588, 288–297. [Google Scholar] [CrossRef]
- Dobrenkov, K.; Ostrovnaya, I.; Gu, J.; Cheung, I.Y.; Cheung, N.-K.V. Oncotargets GD2 and GD3 are highly expressed in sarcomas of children, adolescents, and young adults. Pediatr. Blood Cancer 2016, 63, 1780–1785. [Google Scholar] [CrossRef]
- Hakomori, S.-I. Tumor-Associated Carbohydrate Antigens Defining Tumor Malignancy: Basis for Development of Anti-Cancer Vaccines. Adv. Exp. Med. Biol. 2001, 491, 369–402. [Google Scholar] [CrossRef] [PubMed]
- Groux-Degroote, S.; Delannoy, P. Cancer-Associated Glycosphingolipids as Tumor Markers and Targets for Cancer Immunotherapy. Int. J. Mol. Sci. 2021, 22, 6145. [Google Scholar] [CrossRef] [PubMed]
- Vakkila, J.; Jaffe, R.; Michelow, M.; Lotze, M.T. Pediatric Cancers Are Infiltrated Predominantly by Macrophages and Contain a Paucity of Dendritic Cells: A Major Nosologic Difference with Adult Tumors. Clin. Cancer Res. 2006, 12, 2049–2054. [Google Scholar] [CrossRef] [PubMed]
- Juhl, H.; Petrella, E.C.; Cheung, N.-K.V.; Bredehorst, R.; Vogel, C.-W. Additive cytotoxicity of different monoclonal antibody-cobra venom factor conjugates for human neuroblastoma cells. Immunobiology 1997, 197, 444–459. [Google Scholar] [CrossRef]
- Imai, M.; Landen, C.; Ohta, R.; Cheung, N.-K.V.; Tomlinson, S. Complement-Mediated Mechanisms in Anti-GD2 Monoclonal Antibody Therapy of Murine Metastatic Cancer. Cancer Res. 2005, 65, 10562–10568. [Google Scholar] [CrossRef]
- Forlenza, C.J.; Boudreau, J.; Zheng, J.; Le Luduec, J.-B.; Chamberlain, E.; Heller, G.; Cheung, N.-K.V.; Hsu, K.C. KIR3DL1 Allelic Polymorphism and HLA-B Epitopes Modulate Response to Anti-GD2 Monoclonal Antibody in Patients with Neuroblastoma. J. Clin. Oncol. 2016, 34, 2443–2451. [Google Scholar] [CrossRef]
- Siebert, N.; Jensen, C.; Troschke-Meurer, S.; Zumpe, M.; Jüttner, M.; Ehlert, K.; Kietz, S.; Müller, I.; Lode, H.N. Neuroblastoma patients with high-affinity FCGR2A, -3A and stimulatory KIR 2DS2 treated by long-term infusion of anti-GD2 antibody ch14.18/CHO show higher ADCC levels and improved event-free survival. OncoImmunology 2016, 5, e1235108. [Google Scholar] [CrossRef]
- Rueda, F.; Martí, F.; Pardo, N.; Badell, I.; Peiró, M.; Bertran, E.; Villén, E.; García, J.; Cubells, J. Interleukin-2 in neuroblastoma: Clinical perspectives based on biological studies. Cancer Biother. Radiopharm. 1996, 11, 303–308. [Google Scholar] [CrossRef]
- Thomann, M.; Schlothauer, T.; Dashivets, T.; Malik, S.; Avenal, C.; Bulau, P.; Rüger, P.; Reusch, D. In Vitro Glycoengineering of IgG1 and Its Effect on Fc Receptor Binding and ADCC Activity. PLoS ONE 2015, 10, e0134949. [Google Scholar] [CrossRef]
- Cheung, N.-K.V.; Sowers, R.; Vickers, A.; Cheung, I.Y.; Kushner, B.H.; Gorlick, R. FCGR2A Polymorphism Is Correlated with Clinical Outcome After Immunotherapy of Neuroblastoma With Anti-GD2 Antibody and Granulocyte Macrophage Colony-Stimulating Factor. J. Clin. Oncol. 2006, 24, 2885–2890. [Google Scholar] [CrossRef]
- Long, K.B.; Beatty, G.L. Harnessing the antitumor potential of macrophages for cancer immunotherapy. OncoImmunology 2013, 2, e26860. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.E.; Hirano, M.; Yu, R.K.; Seyfried, T. GD3 ganglioside is a glycolipid characteristic of immature neuroectodermal cells. J. Neuroimmunol. 1985, 7, 179–192. [Google Scholar] [CrossRef]
- Schulz, G.; Cheresh, D.A.; Varki, N.M.; Yu, A.; Staffileno, L.K.; Reisfeld, R.A. Detection of ganglioside GD2 in tumor tissues and sera of neuroblastoma patients. Cancer Res. 1984, 44, 5914–5920. [Google Scholar]
- Saito, M.; Yu, R.K.; Cheung, N.-K. Ganglioside GD2 specificity of monoclonal antibodies to human neuroblastoma cell. Biochem. Biophys. Res. Commun. 1984, 127, 1–7. [Google Scholar] [CrossRef]
- Cheung, N.K.; Cheung, I.Y.; Cañete, A.; Yeh, S.J.; Kushner, B.; Bonilla, M.A.; Heller, G.; Larson, S.M. Antibody response to murine anti-GD2 monoclonal antibodies: Correlation with patient survival. Cancer Res. 1994, 54, 2228–2233. [Google Scholar]
- Barker, E.; Mueller, B.M.; Handgretinger, R.; Herter, M.; Yu, A.L.; Reisfeld, R.A. Effect of a chimeric anti-ganglioside GD2 antibody on cell-mediated lysis of human neuroblastoma cells. Cancer Res. 1991, 51, 144–149. [Google Scholar] [PubMed]
- Uttenreuther-Fischer, M.M.; Huang, C.S.; Yu, A.L. Pharmacokinetics of human-mouse chimeric anti-GD2 mAb ch14.18 in a phase I trial in neuroblastoma patients. Cancer Immunol. Immunother. 1995, 41, 331–338. [Google Scholar] [CrossRef]
- Ladenstein, R.; Weixler, S.; Baykan, B.; Bleeke, M.; Kunert, R.; Katinger, D.; Pribill, I.; Glander, P.; Bauer, S.; Pistoia, V.; et al. Ch14.18 antibody produced in CHO cells in relapsed or refractory Stage 4 neuroblastoma patients: A SIOPEN Phase 1 study. MAbs 2013, 5, 801–809. [Google Scholar] [CrossRef]
- Navid, F.; Sondel, P.M.; Barfield, R.; Shulkin, B.L.; Kaufman, R.A.; Allay, J.A.; Gan, J.; Hutson, P.; Seo, S.; Kim, K.; et al. Phase I Trial of a Novel Anti-GD2 Monoclonal Antibody, Hu14.18K322A, Designed to Decrease Toxicity in Children with Refractory or Recurrent Neuroblastoma. J. Clin. Oncol. 2014, 32, 1445–1452. [Google Scholar] [CrossRef]
- Albertini, M.R.; Hank, J.A.; Gadbaw, B.; Kostlevy, J.; Haldeman, J.; Schalch, H.; Gan, J.; Kim, K.M.; Eickhoff, J.; Gillies, S.D.; et al. Phase II trial of hu14.18-IL2 for patients with metastatic melanoma. Cancer Immunol. Immunother. 2012, 61, 2261–2271. [Google Scholar] [CrossRef]
- Cheung, I.Y.; Kushner, B.H.; Modak, S.; Basu, E.M.; Roberts, S.S.; Cheung, N.-K.V. Phase I trial of anti-GD2 monoclonal antibody hu3F8 plus GM-CSF: Impact of body weight, immunogenicity and anti-GD2 response on pharmacokinetics and survival. OncoImmunology 2017, 6, e1358331. [Google Scholar] [CrossRef] [PubMed]
- Siebert, N.; Eger, C.; Seidel, D.; Jüttner, M.; Lode, H.N. Validated detection of human anti-chimeric immune responses in serum of neuroblastoma patients treated with ch14.18/CHO. J. Immunol. Methods 2014, 407, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.L.; Gilman, A.L.; Ozkaynak, M.F.; Naranjo, A.; Diccianni, M.B.; Gan, J.; Hank, J.A.; Batova, A.; London, W.B.; Tenney, S.C.; et al. Long-Term Follow-up of a Phase III Study of ch14.18 (Dinutuximab) + Cytokine Immunotherapy in Children with High-Risk Neuroblastoma: COG Study ANBL0032. Clin. Cancer Res. 2021, 27, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Uttenreuther-Fischer, M.M.; Kruger, J.A.; Fischer, P. Molecular characterization of the anti-idiotypic immune response of a relapse-free neuroblastoma patient following antibody therapy: A possible vaccine against tumors of neuroectodermal origin? J. Immunol. 2006, 176, 7775–7786. [Google Scholar] [CrossRef]
- Chatterjee, M.B.; Foon, K.A.; Kohler, H. Idiotypic antibody immunotherapy of cancer. Cancer Immunol. Immunother. 1994, 38, 75–82. [Google Scholar] [CrossRef]
- Cheung, N.K.; Guo, H.F.; Cheung, I.Y. Correlation of anti-idiotype network with survival following anti-G(D2) monoclonal antibody 3F8 therapy of stage 4 neuroblastoma. Med. Pediatr. Oncol. 2000, 35, 635–637. [Google Scholar] [CrossRef]
- Cheung, N.K.; Kushner, B.H.; Cheung, I.Y.; Kramer, K.; Canete, A.; Gerald, W.; Bonilla, M.A.; Finn, R.; Yeh, S.J.; Larson, S.M. Anti-G(D2) antibody treatment of minimal residual stage 4 neuroblastoma diagnosed at more than 1 year of age. J. Clin. Oncol. 1998, 16, 3053–3060. [Google Scholar] [CrossRef]
- Lode, H.N.; Schmidt, M.; Seidel, D.; Hübener, N.; Brackrock, D.; Bleeke, M.; Reker, D.; Brandt, S.; Mueller, H.-P.; Helm, C.A.; et al. Vaccination with anti-idiotype antibody ganglidiomab mediates a GD2-specific anti-neuroblastoma immune response. Cancer Immunol. Immunother. 2013, 62, 999–1010. [Google Scholar] [CrossRef]
- Kushner, B.H.; Cheung, I.Y.; Modak, S.; Kramer, K.; Ragupathi, G.; Cheung, N.K. Phase I trial of a bivalent gangliosides vaccine in combination with beta-glucan for high-risk neuroblastoma in second or later remission. Clin. Cancer Res. 2014, 20, 1375–1382. [Google Scholar] [CrossRef]
- Cheung, I.Y.; Cheung, N.K.V.; Modak, S.; Mauguen, A.; Feng, Y.; Basu, E.; Roberts, S.S.; Ragupathi, G.; Kushner, B.H. Survival Impact of Anti-GD2 Antibody Response in a Phase II Ganglioside Vaccine Trial Among Patients with High-Risk Neuroblastoma With Prior Disease Progression. J. Clin. Oncol. 2021, 39, 215–226. [Google Scholar] [CrossRef]
- Ahmed, M.; Hu, J.; Cheung, N.-K.V. Structure Based Refinement of a Humanized Monoclonal Antibody That Targets Tumor Antigen Disialoganglioside GD2. Front. Immunol. 2014, 5, 372. [Google Scholar] [CrossRef]
- Park, J.A.; Cheung, N.K.V. Targets and Antibody Formats for Immunotherapy of Neuroblastoma. J. Clin. Oncol. 2020, 38, 1836–1848. [Google Scholar] [CrossRef]
- Markham, A. Naxitamab: First Approval. Drugs 2021, 81, 291–296. [Google Scholar] [CrossRef]
- Dumont, J.A.; Euwart, D.; Mei, B.; Estes, S.; Kshirsagar, R.R. Human cell lines for biopharmaceutical manufacturing: History, status, and future perspectives. Crit. Rev. Biotechnol. 2015, 36, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Yuile, A.; Fanuli, C.; van Nunen, S.; Diakos, C.; Clarke, S.; Steiner, C.; MacMillan, R.; Oatley, M.; Pavlakis, N. Increased rates of cetuximab reactions in tick prevalent regions and a proposed protocol for risk mitigation. Asia-Pac. J. Clin. Oncol. 2020, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Ladenstein, R.; Pötschger, U.; Valteau-Couanet, D.; Luksch, R.; Castel, V.; Yaniv, I.; Laureys, G.; Brock, P.; Michon, J.M.; Owens, C.; et al. Interleukin 2 with anti-GD2 antibody ch14.18/CHO (dinutuximab beta) in patients with high-risk neuroblastoma (HR-NBL1/SIOPEN): A multicentre, randomised, phase 3 trial. Lancet Oncol. 2018, 19, 1617–1629. [Google Scholar] [CrossRef]
- Sorkin, L.S.; Otto, M.; Baldwin, W.M., III; Vail, E.; Gillies, S.D.; Handgretinger, R.; Barfield, R.C.; Yu, H.M.; Yu, A.L. Anti-GD(2) with an FC point mutation reduces complement fixation and decreases antibody-induced allodynia. Pain 2010, 149, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Terme, M.; Dorvillius, M.; Cochonneau, D.; Chaumette, T.; Xiao, W.; Diccianni, M.B.; Barbet, J.; Yu, A.L.; Paris, F.; Sorkin, L.S.; et al. Chimeric Antibody c.8B6 to O-Acetyl-GD2 Mediates the Same Efficient Anti-Neuroblastoma Effects as Therapeutic ch14.18 Antibody to GD2 without Antibody Induced Allodynia. PLoS ONE 2014, 9, e87210. [Google Scholar] [CrossRef] [PubMed]
- Fleurence, J.; Fougeray, S.; Bahri, M.; Cochonneau, D.; Clémenceau, B.; Paris, F.; Heczey, A.; Birklé, S. Targeting O-Acetyl-GD2 Ganglioside for Cancer Immunotherapy. J. Immunol. Res. 2017, 2017, 5604891. [Google Scholar] [CrossRef]
- Cavdarli, S.; Delannoy, P.; Groux-Degroote, S. O-acetylated Gangliosides as Targets for Cancer Immunotherapy. Cells 2020, 9, 741. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Shi, W.; Shi, J.-J.; Lu, J.-J. Progress of CD47 immune checkpoint blockade agents in anticancer therapy: A hematotoxic perspective. J. Cancer Res. Clin. Oncol. 2021, 148, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bahri, M.; Kailayangiri, S.; Vermeulen, S.; Galopin, N.; Rossig, C.; Paris, F.; Fougreay, S.; Birkle, S. SIRPalpha-specific monoclonal antibody enables antibody-dependent phagocytosis of neuroblastoma cells. Cancer Immunol. Immunother 2021, 71, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Dobrenkov, K.; Cheung, N.-K.V. GD2-Targeted Immunotherapy and Radioimmunotherapy. Semin. Oncol. 2014, 41, 589–612. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.K.; Kalogriopoulos, N.A.; Rakhmilevich, A.L.; Ranheim, E.A.; Seo, S.; Kim, K.; Alderson, K.L.; Gan, J.; Reisfeld, R.A.; Gillies, S.D.; et al. Intratumoral hu14.18–IL-2 (IC) Induces Local and Systemic Antitumor Effects That Involve Both Activated T and NK Cells as Well as Enhanced IC Retention. J. Immunol. 2012, 189, 2656–2664. [Google Scholar] [CrossRef] [PubMed]
- Sondel, P.M.; Gillies, S.D. Current and Potential Uses of Immunocytokines as Cancer Immunotherapy. Antibodies 2012, 1, 149–171. [Google Scholar] [CrossRef]
- Dhillon, S. Dinutuximab: First global approval. Drugs 2015, 75, 923–927. [Google Scholar] [CrossRef]
- Simon, T.; Hero, B.; Faldum, A.; Handgretinger, R.; Schrappe, M.; Klingebiel, T.; Berthold, F. Long term outcome of high-risk neuroblastoma patients after immunotherapy with antibody ch14.18 or oral metronomic chemotherapy. BMC Cancer 2011, 11, 21. [Google Scholar] [CrossRef]
- Lissoni, P. Therapy implications of the role of interleukin-2 in cancer. Expert Rev. Clin. Immunol. 2016, 13, 491–498. [Google Scholar] [CrossRef]
- Mora, J.; Castañeda, A.; Gorostegui, M.; Santa-María, V.; Garraus, M.; Muñoz, J.P.; Varo, A.; Perez-Jaume, S.; Mañe, S. Naxitamab combined with granulocyte-macrophage colony-stimulating factor as consolidation for high-risk neuroblastoma patients in complete remission. Pediatr. Blood Cancer 2021, 68, e29121. [Google Scholar] [CrossRef]
- Furman, W.L.; Federico, S.M.; McCarville, M.B.; Shulkin, B.L.; Davidoff, A.M.; Krasin, M.J.; Sahr, N.; Sykes, A.D.; Wu, J.; Brennan, R.C.; et al. A Phase II Trial of Hu14.18K322A in Combination with Induction Chemotherapy in Children with Newly Diagnosed High-Risk Neuroblastoma. Clin. Cancer Res. 2019, 25, 6320–6328. [Google Scholar] [CrossRef]
- Furman, W.L.; McCarville, B.; Shulkin, B.L.; Davidoff, A.; Krasin, M.; Hsu, C.-W.; Pan, H.; Wu, J.; Brennan, R.; Bishop, M.W.; et al. Improved Outcome in Children with Newly Diagnosed High-Risk Neuroblastoma Treated with Chemoimmunotherapy: Updated Results of a Phase II Study Using hu14.18K322A. J. Clin. Oncol. 2022, 40, 335–344. [Google Scholar] [CrossRef]
- Mody, R.; Naranjo, A.; Van Ryn, C.; Alice, L.Y.; London, W.B.; Shulkin, B.L.; Parisi, M.T.; Servaes, S.E.N.; Dicciani, M.B.; Sondel, P.M.; et al. Irinotecan-temozolomide with temsirolimus or dinutuximab in children with refractory or relapsed neuroblastoma (COG ANBL1221): An open-label, randomised, phase 2 trial. Lancet Oncol. 2017, 18, 946–957. [Google Scholar] [CrossRef]
- Siebert, N.; Eger, C.; Seidel, D.; Jüttner, M.; Zumpe, M.; Wegner, D.; Kietz, S.; Ehlert, K.; Veal, G.J.; Siegmund, W.; et al. Pharmacokinetics and pharmacodynamics of ch14.18/CHO in relapsed/refractory high-risk neuroblastoma patients treated by long-term infusion in combination with IL-2. MAbs 2016, 8, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Kushner, B.H.; Cheung, I.Y.; Modak, S.; Basu, E.M.; Roberts, S.S.; Cheung, N.K. Humanized 3F8 Anti-GD2 Monoclonal Antibody Dosing with Granulocyte-Macrophage Colony-Stimulating Factor in Patients With Resistant Neuroblastoma: A Phase 1 Clinical Trial. JAMA Oncol. 2018, 4, 1729–1735. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.; Castañeda, A.; Colombo, M.; Gorostegui, M.; Gomez, F.; Mañe, S.; Santa-Maria, V.; Garraus, M.; Macias, N.; Perez-Jaume, S.; et al. Clinical and Pathological Evidence of Anti-GD2 Immunotherapy Induced Differentiation in Relapsed/Refractory High-Risk Neuroblastoma. Cancers 2021, 13, 1264. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.; Kushner, B.; Heller, G.; Cheung, N.K. Neuroblastoma metastatic to the central nervous system. The Memorial Sloan-kettering Cancer Center Experience and A Literature Review. Cancer 2001, 91, 1510–1519. [Google Scholar] [CrossRef]
- Berlanga, P.; Pasqualini, C.; Pötschger, U.; Sangüesa, C.; Castellani, M.R.; Cañete, A.; Luksch, R.; Elliot, M.; Schreier, G.; Kropf, M.; et al. Central nervous system relapse in high-risk stage 4 neuroblastoma: The HR-NBL1/SIOPEN trial experience. Eur. J. Cancer 2021, 144, 1–8. [Google Scholar] [CrossRef]
- Kramer, K.; Kushner, B.H.; Modak, S.; Pandit-Taskar, N.; Smith-Jones, P.; Zanzonico, P.; Humm, J.L.; Xu, H.; Wolden, S.L.; Souweidane, M.M.; et al. Compartmental intrathecal radioimmunotherapy: Results for treatment for metastatic CNS neuroblastoma. J. Neuro-Oncol. 2009, 97, 409–418. [Google Scholar] [CrossRef]
- Yerrabelli, R.S.; He, P.; Fung, E.K.; Kramer, K.; Zanzonico, P.B.; Humm, J.L.; Guo, H.; Pandit-Taskar, N.; Larson, S.M.; Cheung, N.-K.V. IntraOmmaya compartmental radioimmunotherapy using 131I-omburtamab—pharmacokinetic modeling to optimize therapeutic index. Eur. J. Nucl. Med. Mol. Imaging 2020, 48, 1166–1177. [Google Scholar] [CrossRef]
- Martinez, C.; Hofmann, T.J.; Marino, R.; Dominici, M.; Horwitz, E.M. Human bone marrow mesenchymal stromal cells express the neural ganglioside GD2: A novel surface marker for the identification of MSCs. Blood 2007, 109, 4245–4248. [Google Scholar] [CrossRef]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.N.; Khazaeli, M.B.; Wheeler, R.H.; Dropcho, E.; Liu, T.; Urist, M.; Miller, D.M.; Lawson, S.; Dixon, P.; Russell, C.H. Phase I trial of the murine monoclonal anti-GD2 antibody 14G2a in metastatic melanoma. Cancer Res. 1992, 52, 4342–4347. [Google Scholar] [PubMed]
- Blom, T.; Lurvink, R.; Aleven, L.; Mensink, M.; Wolfs, T.; Dierselhuis, M.; van Eijkelenburg, N.; Kraal, K.; van Noesel, M.; van Grotel, M.; et al. Treatment-Related Toxicities During Anti-GD2 Immunotherapy in High-Risk Neuroblastoma Patients. Front. Oncol. 2021, 10, 601076. [Google Scholar] [CrossRef] [PubMed]
- Gillin, S.; Sorkin, L.S. Gabapentin reverses the allodynia produced by the administration of anti-GD2 ganglioside, an immunotherapeutic drug. Anesth. Analg. 1998, 86, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Tse, B.C.; Navid, F.; Billups, C.A.; O’Donnell, T.; Hoehn, M.E. Ocular abnormalities in patients treated with a novel anti-GD2 monoclonal antibody, hu14.18K322A. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2015, 19, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Kremens, B.H.B.; Kremens, B.; Hero, B.; Esser, J.; Weinel, P.; Filger-Brillinger, J.; Fleischhack, G.; Graf, N.; Grüttner, H.-P.; Niemeyer, C.; et al. Ocular symptoms in children treated with human-mouse chimeric anti-GD2 mAb ch14.18 for neuroblastoma. Cancer Immunol. Immunother. 2002, 51, 107–110. [Google Scholar] [CrossRef]
- Yuki, N.; Yamada, M.; Tagawa, Y.; Takahashi, H.; Handa, S. Pathogenesis of the neurotoxicity caused by anti-GD2 antibody therapy. J. Neurol. Sci. 1997, 149, 127–130. [Google Scholar] [CrossRef]
- Furman, W.L. Monoclonal Antibody Therapies for High Risk Neuroblastoma. Biol. Targets Ther. 2021, 15, 205–219. [Google Scholar] [CrossRef]
- Vanichapol, T.; Chutipongtanate, S.; Anurathapan, U.; Hongeng, S. Immune Escape Mechanisms and Future Prospects for Immunotherapy in Neuroblastoma. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Caforio, M.; Sorino, C.; Caruana, I.; Weber, G.; Camera, A.; Cifaldi, L.; De Angelis, B.; Del Bufalo, F.; Vitale, A.; Goffredo, B.M.; et al. GD2 redirected CAR T and activated NK-cell-mediated secretion of IFNgamma overcomes MYCN-dependent IDO1 inhibition, contributing to neuroblastoma cell immune escape. J. Immunother. Cancer 2021, 9, e001502. [Google Scholar] [CrossRef]
- Liang, H.; Li, T.; Fang, X.; Xing, Z.; Zhang, S.; Shi, L.; Li, W.; Guo, L.; Kuang, C.; Lui, H.; et al. IDO1/TDO dual inhibitor RY103 targets Kyn-AhR pathway and exhibits preclinical efficacy on pancreatic cancer. Cancer Lett. 2021, 522, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Guo, H.F.; Hoseini, S.S.; Suzuki, M.; Xu, H.; Cheung, N.V. Potent antitumor effect of T cells armed with anti-GD2 bispecific antibody. Pediatr. Blood Cancer 2021, 68, e28971. [Google Scholar] [CrossRef] [PubMed]
- Van den Bijgaart, R.J.E.; Kroesen, M.; Brok, I.C.; Reijnen, D.; Wassink, M.; Boon, L.; Hoogerbrugge, P.M.; Adema, G.J. Anti-GD2 antibody and Vorinostat immunocombination therapy is highly effective in an aggressive orthotopic neuroblastoma model. Oncoimmunology 2020, 9, 1817653. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef]
- Sze, D.M.-Y.; Chan, G.C.F. Supplements for immune enhancement in hematologic malignancies. Hematol. Am. Soc. Hematol. Educ. Program 2009, 2009, 313–319. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hou, B.; Tang, Y.; Li, W.; Zeng, Q.; Chang, D. Efficiency of CAR-T Therapy for Treatment of Solid Tumor in Clinical Trials: A Meta-Analysis. Dis. Markers 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Richards, R.M.; Sotillo, E.; Majzner, R.G. CAR T Cell Therapy for Neuroblastoma. Front. Immunol. 2018, 9, 2380. [Google Scholar] [CrossRef]
- Richman, S.A.; Nunez-Cruz, S.; Moghimi, B.; Li, L.; Gershenson, Z.T.; Mourelatos, Z.; Barrett, D.M.; Grupp, S.A.; Milone, M.C. High-Affinity GD2-Specific CAR T Cells Induce Fatal Encephalitis in a Preclinical Neuroblastoma Model. Cancer Immunol. Res. 2017, 6, 36–46. [Google Scholar] [CrossRef]
- Sawaisorn, P.; Atjanasuppat, K.; Anurathapan, U.; Chutipongtanate, S.; Hongeng, S. Strategies to Improve Chimeric Antigen Receptor Therapies for Neuroblastoma. Vaccines 2020, 8, 753. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Immunotherapy Included | Treatment Schedule | Number of Subjects | Median Follow-Up | EFS | OS |
|---|---|---|---|---|---|---|
| Yu A, et al., 2011 | Dinutuximab, GMCSF, IL-2 | As maintenance after chemotherapy and auto-PBSCT | Randomized trial N = 113 (anti-GD2) N = 113 (standard) | 2.1 years (4 days–6.9 years) | 2 years EFS 66 ± 5% (anti-GD2) 46 ± 5% (standard) | 2 years OS 86 ± 4% (anti-GD2) 75 ± 5% (standard) |
| Yu A, et al., 2021 (follow-up study) | Same as above | Same as above | Same as above | 9.97 years (0.7 years–5.3 years) | 5 years EFS 56.6 ± 5% (anti-GD2) 46.1 ± 5% (standard) | 5 years OS 73.2 ± 4% (anti-GD2) 56.6 ± 5% (standard) |
| Simon T, et al., 2011 | Ch14.18 (BioInvent) | As maintenance after chemotherapy with or without auto-PBSCT | Non-randomized cohort study N = 164 (anti-GD2) | 11.1 years (2.3 years–8.6 years) | 5 years EFS 51.3 ± 6% (anti-GD2) 34.1 ± 5% (standard) 9 years EFS 44.7 ± 6% (anti-GD2) 31 ± 5% (standard) | 5 years OS 60.3 ± 6% (anti-GD2) 42.2 ± 5% (standard) 9 years EFS 46.6 ± 6% (anti-GD2) 33.9 ± 5% (standard) |
| Ladenstein R, et al., 2018 | Dinutuximab β, with or without IL-2 | As maintenance after chemotherapy and auto-PBSCT | Randomized trial N = 206 (anti-GD2 + Il2) N = 200 (anti-GD2) | 4.7 years (3.9 years–5.3 years) | 5 years EFS 53 ± 7% (anti-GD2) 60 ± 6% (anti-GD2 + IL-2) | 5 years OS 63 ± 8% (anti-GD2) 62 ± 7% (anti-GD2 + IL-2) |
| Ladenstein R, et al., 2020 | Dinutuximab β, with or without IL-2 | As maintenance after chemotherapy and auto-PBSCT | Non-randomized cohort study (Historical control) N = 378 (anti-GD2) N = 466 (standard) | 5.8 years (4.2 years–8.2 years) 4.6 years for anti-GD2 & 8.6 years for standard arm | 5 years EFS 57 ± 6% (anti-GD2) 42 ± 5% (standard) | 5 years EFS 64 ± 5% (anti-GD2) 50 ± 5% (standard) |
| Adverse Events (Gr 3 or 4) | hu3F8 (+GMCSF) (Mora et al.) | Dinutuximab β (no IL-2) (Ladenstein, et al.) | Dinutuximab β (+IL-2) (Ladenstein, et al.) | Dinutuximab (+IL-2 & GMCSF) (Yu, et al.) |
|---|---|---|---|---|
| Hypotension | 63% | 7% | 17% | 18% |
| Pain | 65% | 66% | 86% | 52% |
| Urtricaria | 29% | 5% | 10% | 13% |
| Pyrexia | 2% | 14% | 40% | 39% |
| Bronchospasm | 21% | 0% | 0% | 0% |
| Elevated ALT/AST | 0% | 17% | 23% | 23% |
| Nausea & vomiting/diarrhea | 2%/0% | 5%/7% | 9%/21% | 6%/13% |
| Deranged renal function | 0% | 2% | 1% | Hypokalemia (35%) Hyponatremia (23%) |
| Neutropenia | 15% | 33% | 58% | - |
| Anemia | 0% | 42% | 66% | - |
| Lethargy | 10% | 0% | 6% | 0% |
| Hypoxia | 10% | 0% | 0% | 13% |
| Allergy | 0% | 10% | 20% | 25% |
| Neuropathy | 0% | 3% | 9% | 4% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, G.C.-F.; Chan, C.M. Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma. Biomolecules 2022, 12, 358. https://doi.org/10.3390/biom12030358
Chan GC-F, Chan CM. Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma. Biomolecules. 2022; 12(3):358. https://doi.org/10.3390/biom12030358
Chicago/Turabian StyleChan, Godfrey Chi-Fung, and Carol Matias Chan. 2022. "Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma" Biomolecules 12, no. 3: 358. https://doi.org/10.3390/biom12030358
APA StyleChan, G. C.-F., & Chan, C. M. (2022). Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma. Biomolecules, 12(3), 358. https://doi.org/10.3390/biom12030358