
Examining the Effect of Charged Lipids on Mitochondrial Outer Membrane Dynamics Using Atomistic Simulations

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
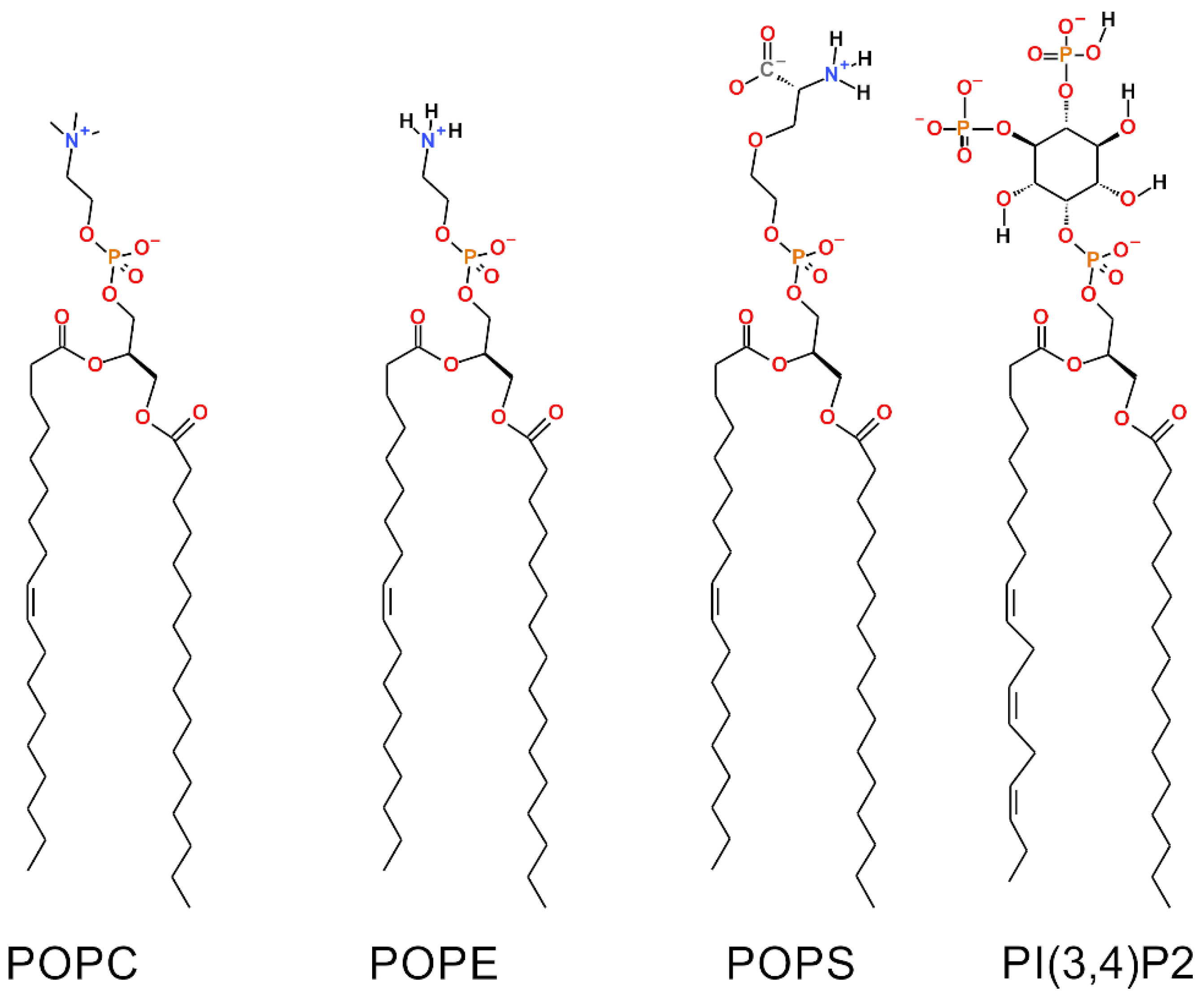
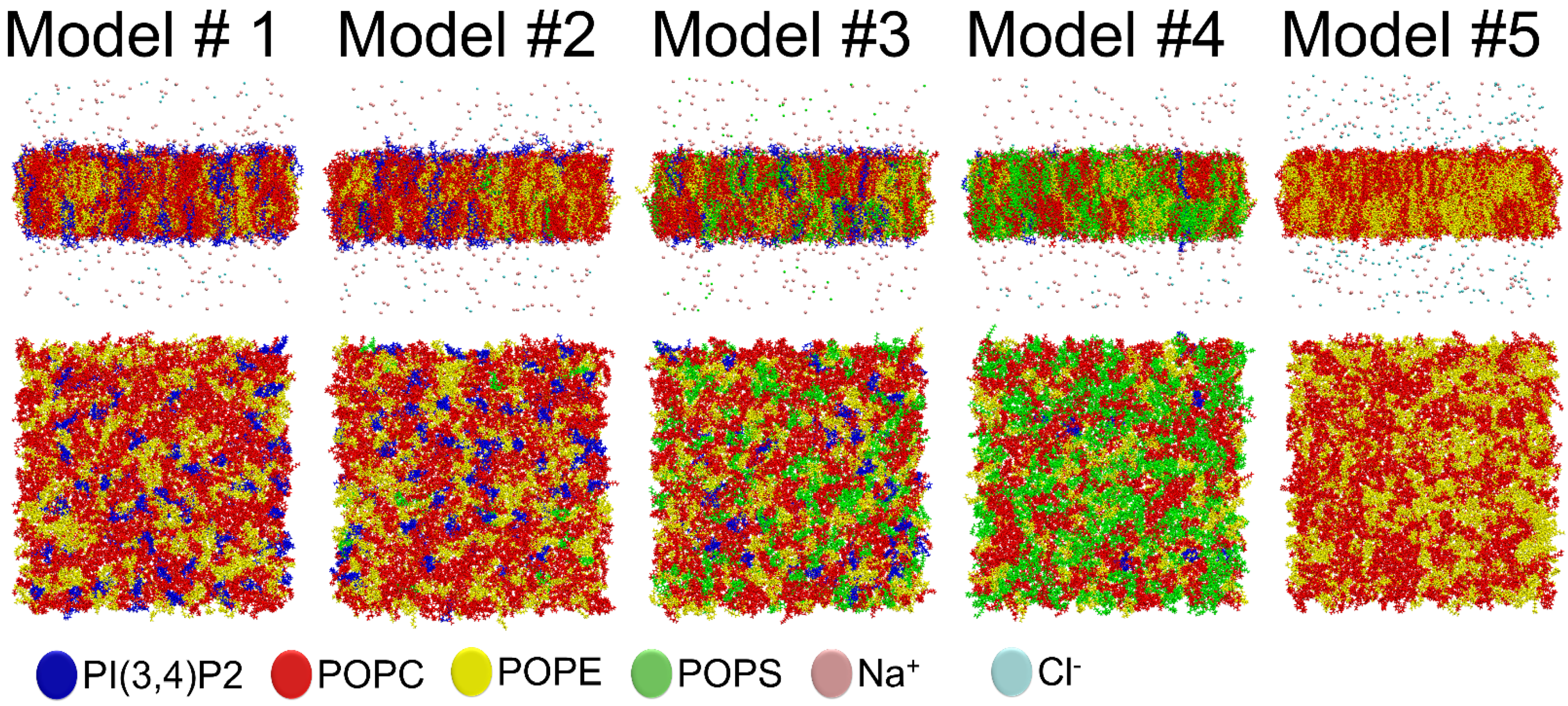
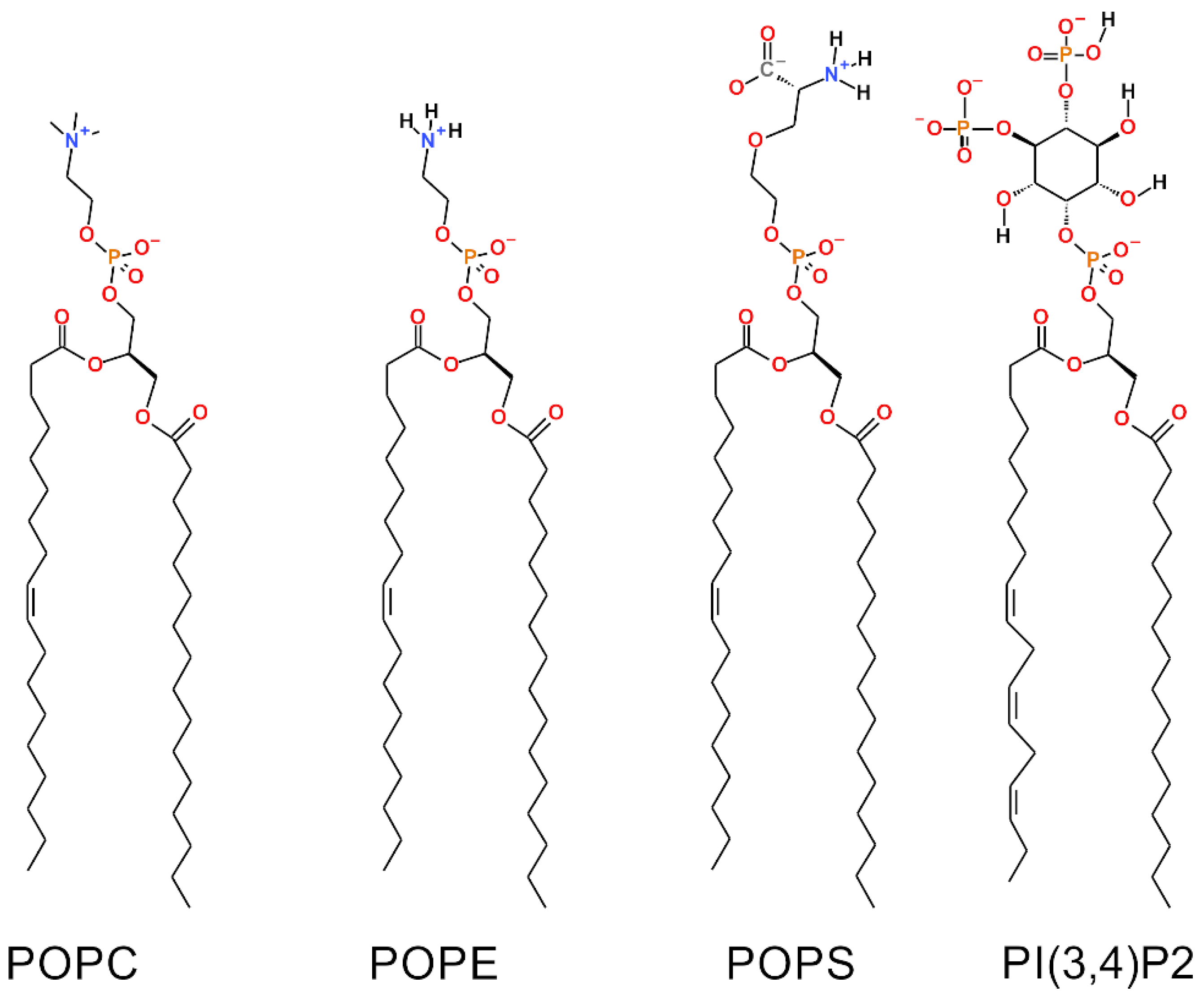
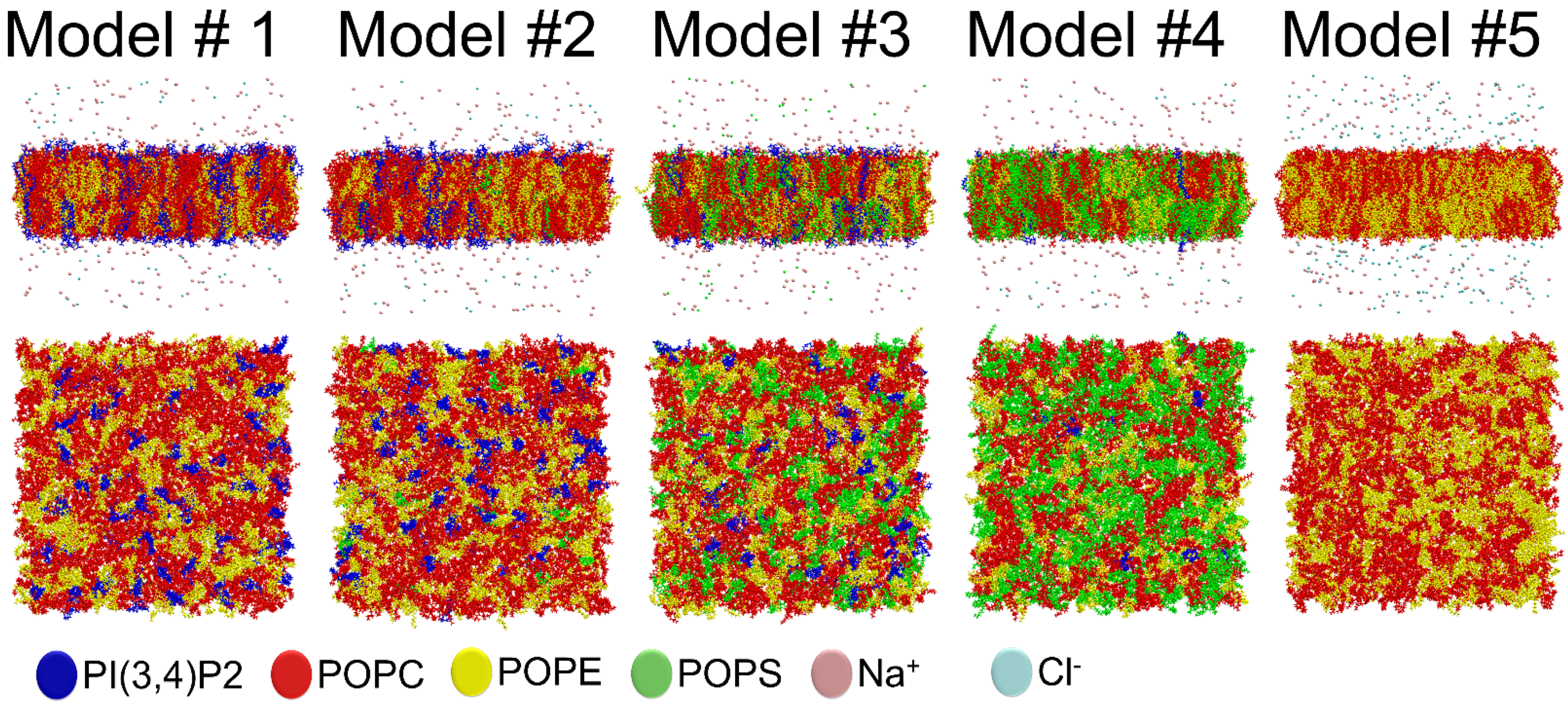
2.1. Outer Mitochondrial Membrane Models
2.2. Molecular Dynamics Simulations
2.3. Analysis and Figure Preparation
3. Results
3.1. Phosphatidylinositol Increases the Surface Area per Lipid
3.2. Phosphatidylserine Increases the Bilayer Thickness
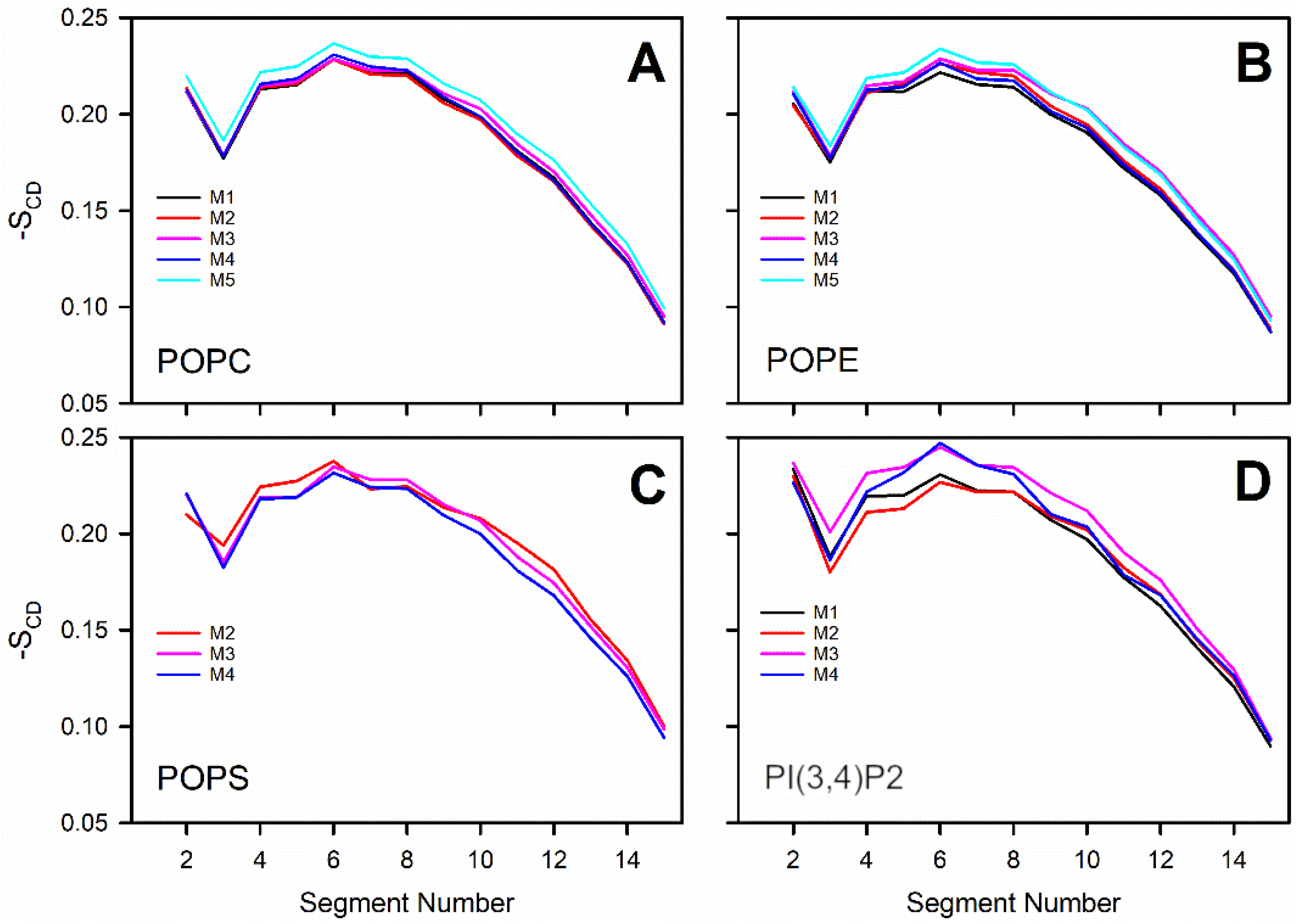
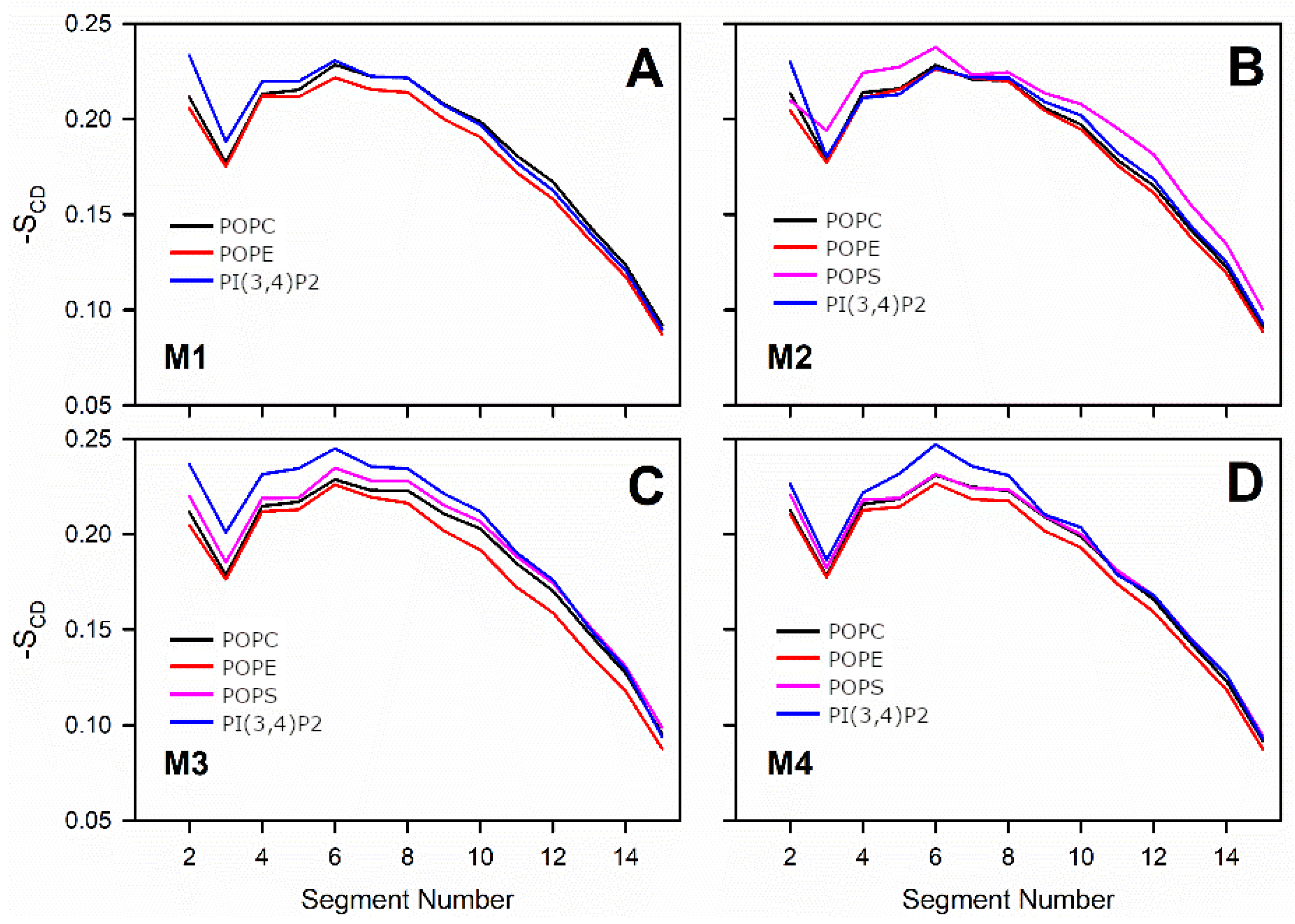
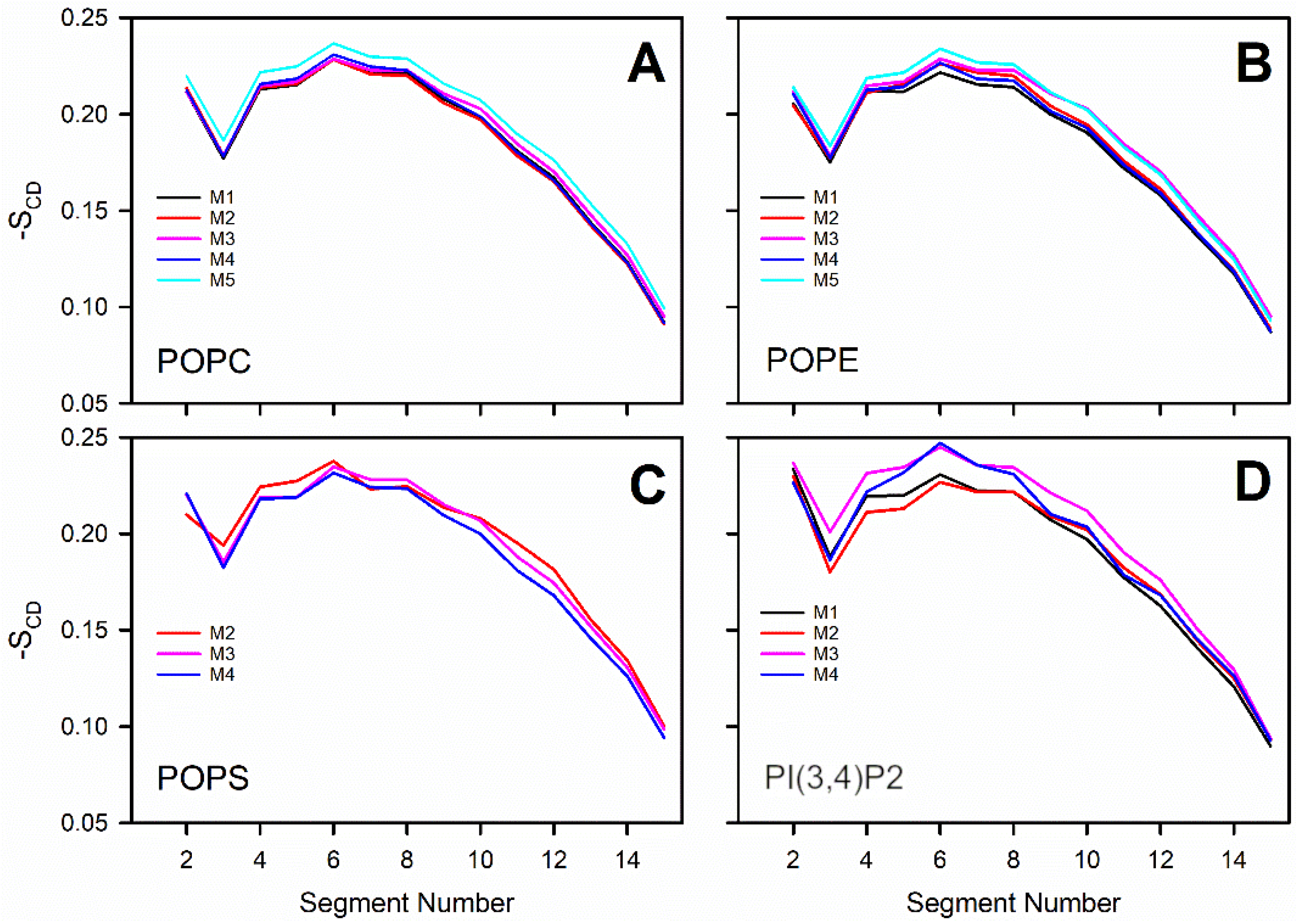
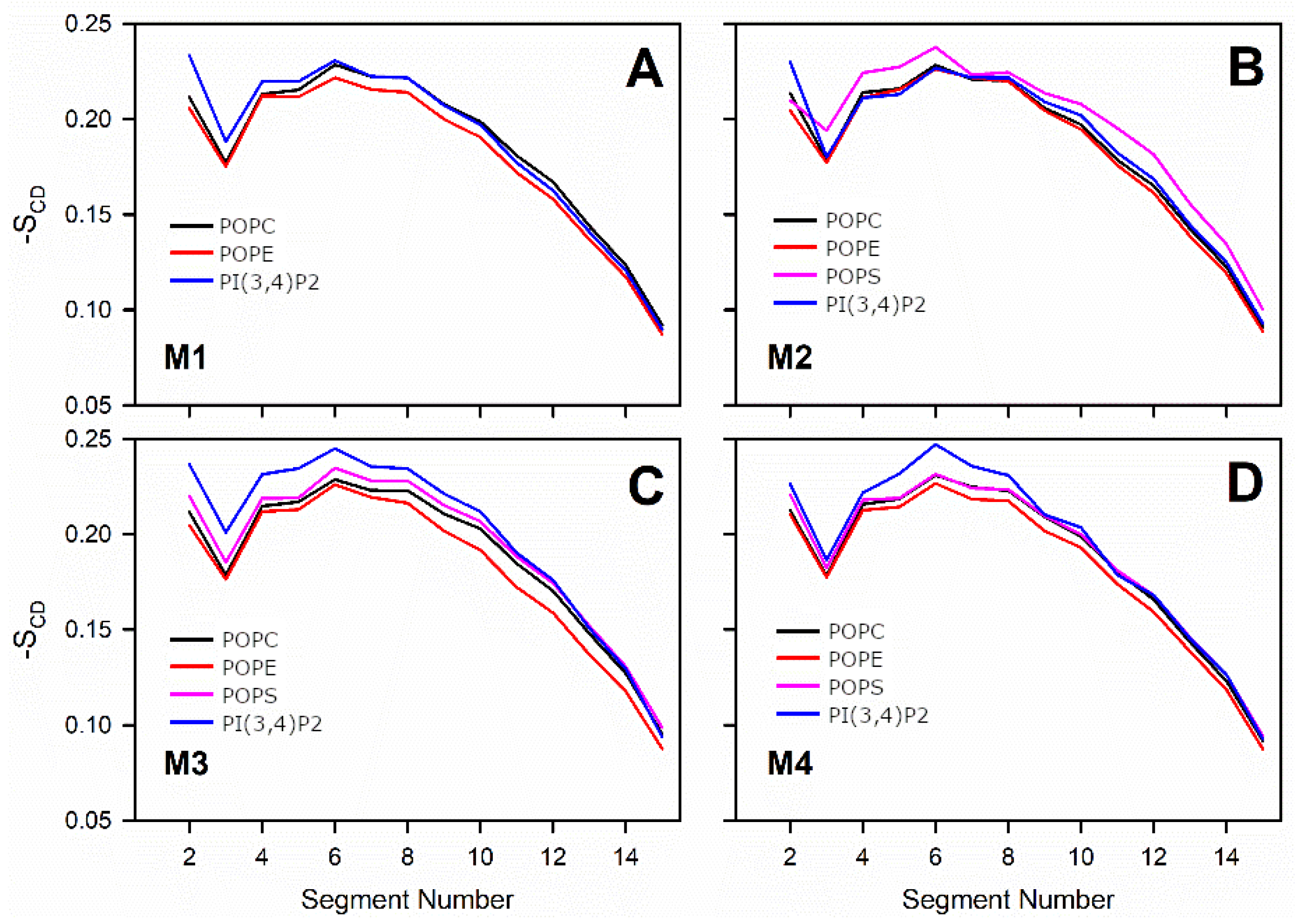
3.3. Negatively Charged Lipids Lower the Order Parameter
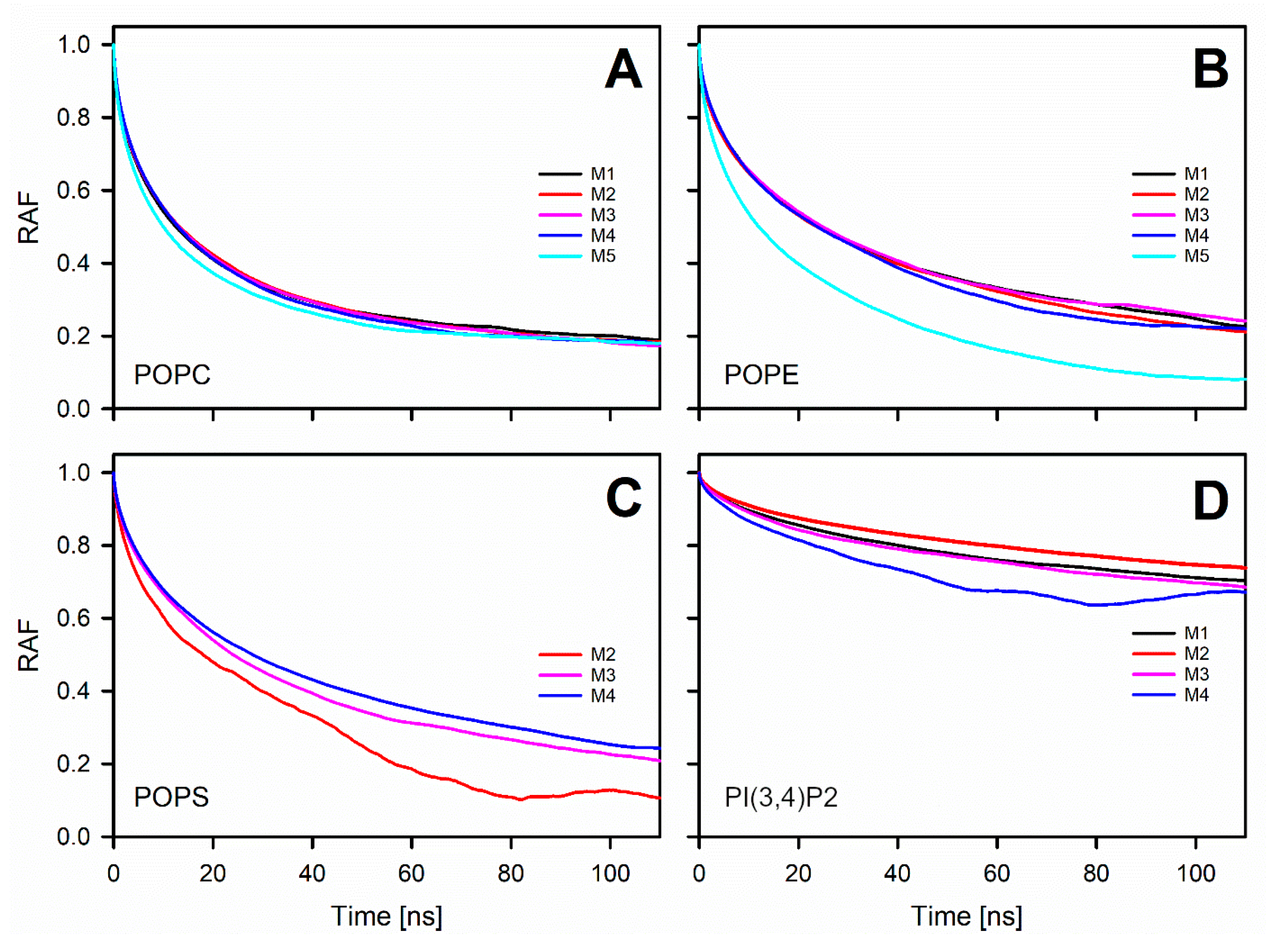
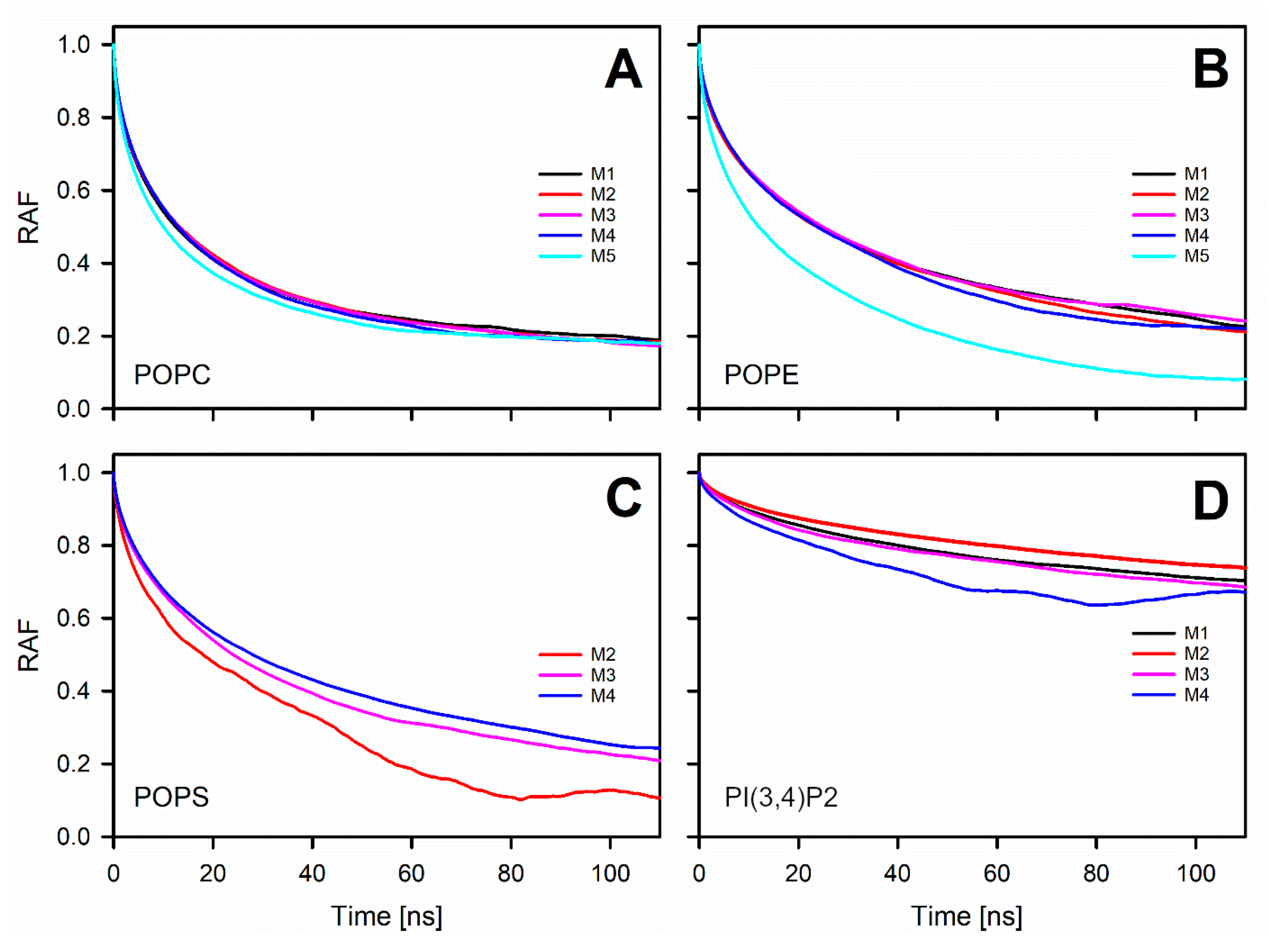
3.4. Phosphatidylserine Reduces the Lateral Diffusion
3.5. Charged Lipids Reduce the Rotational Diffusion Rate of the POPE Headgroup
3.6. Interactions at the Water-Membrane Interface
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pöyry, S.; Vattulainen, I. Role of charged lipids in membrane structures—Insight given by simulations. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2322–2333. [Google Scholar] [CrossRef]
- Leventis, P.A.; Grinstein, S. The Distribution and Function of Phosphatidylserine in Cellular Membranes. Annu. Rev. Biophys. 2010, 39, 407–427. [Google Scholar] [CrossRef]
- Ngo, V.A.; Sarkar, S.; Neale, C.; Garcia, A.E. How Anionic Lipids Affect Spatiotemporal Properties of KRAS4B on Model Membranes. J. Phys. Chem. B 2020, 124, 5434–5453. [Google Scholar] [CrossRef]
- Platre, M.P.; Jaillais, Y. Anionic lipids and the maintenance of membrane electrostatics in eukaryotes. Plant Signal. Behav. 2017, 12, e1282022. [Google Scholar] [CrossRef] [Green Version]
- Róg, T.; Girych, M.; Bunker, A. Mechanistic Understanding from Molecular Dynamics in Pharmaceutical Research 2: Lipid Membrane in Drug Design. Pharmaceuticals 2021, 14, 1062. [Google Scholar] [CrossRef] [PubMed]
- Postila, P.A.; Vattulainen, I.; Róg, T. Selective effect of cell membrane on synaptic neurotransmission. Sci. Rep. 2016, 6, 19345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postila, P.A.; Róg, T. A perspective: Active role of lipids in neurotransmitter dynamics. Mol. Neurobiol. 2020, 57, 910–925. [Google Scholar] [CrossRef] [Green Version]
- Vauquelin, G. On the ‘micro’-pharmacodynamic and pharmacokinetic mechanisms that contribute to long-lasting drug action. Expert Opin. Drug Discov. 2015, 10, 1085–1098. [Google Scholar] [CrossRef] [PubMed]
- Róg, T.; Orłowski, A.; Llorente, A.; Skotland, T.; Sylvänne, T.; Kauhanen, D.; Ekroos, K.; Sandvig, K.; Vattulainen, I. Interdigitation of long-chain sphingomyelin induces coupling of membrane leaflets in a cholesterol dependent manner. Biochim. Biophys. Acta Biomembr. 2016, 1858, 281–288. [Google Scholar] [CrossRef]
- Llorente, A.; Skotland, T.; Sylvänne, T.; Kauhanen, D.; Róg, T.; Orłowski, A.; Vattulainen, I.; Ekroos, K.; Sandvig, K.; Róg, T.; et al. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Monje-Galvan, V.; Klauda, J.B. Modeling Yeast Organelle Membranes and How Lipid Diversity Influences Bilayer Properties. Biochemistry 2015, 54, 6852–6861. [Google Scholar] [CrossRef] [PubMed]
- Pietiläinen, K.H.; Róg, T.; Seppänen-Laakso, T.; Virtue, S.; Gopalacharyulu, P.; Tang, J.; Rodriguez-Cuenca, S.; Maciejewski, A.; Naukkarinen, J.; Ruskeepää, A.L.; et al. Association of Lipidome remodeling in the Adipocyte membrane with acquired obesity in humans. PLoS Biol. 2011, 9, e1000623. [Google Scholar] [CrossRef] [Green Version]
- Buckman, T.D.; Eiduson, S.; Sutphin, M.S.; Chang, R. Selective effects on catalysis by the multiple forms of monoamine oxidase produced by interactions of acidic phospholipids with mitochondrial membranes. J. Biol. Chem. 1983, 258, 8670–8676. [Google Scholar] [CrossRef]
- Tachiki, K.H.; Buckman, T.D.; Eiduson, S.; Kling, A.S.; Hullett, J. Phosphatidylserine inhibition of monoamine oxidase in platelets of schizophrenics. Biol. Psychiatry 1986, 21, 59–68. [Google Scholar] [CrossRef]
- Yeung, A.W.K.; Georgieva, M.G.; Atanasov, A.G.; Tzvetkov, N.T. Monoamine Oxidases (MAOs) as Privileged Molecular Targets in Neuroscience: Research Literature Analysis. Front. Mol. Neurosci. 2019, 12, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fišar, Z. Drugs related to monoamine oxidase activity. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 69, 112–124. [Google Scholar] [CrossRef]
- Tait, S.W.G.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef]
- Green, D.R.; Galluzzi, L.; Kroemer, G. Mitochondria and the Autophagy–Inflammation–Cell Death Axis in Organismal Aging. Science 2011, 333, 1109–1112. [Google Scholar] [CrossRef] [Green Version]
- Tuominen, A.; Miller, Y.I.; Hansen, L.F.; Kesäniemi, Y.A.; Witztum, J.L.; Hörkkö, S. A Natural Antibody to Oxidized Cardiolipin Binds to Oxidized Low-Density Lipoprotein, Apoptotic Cells, and Atherosclerotic Lesions. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2096–2102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edmondson, D.E.; Binda, C. Monoamine Oxidases. In Membrane Protein Complexes: Structure and Function. Subcellular Biochemistry; Harris, J.R., Boekema, E.J., Eds.; Springer Singapore: Singapore, 2018; Volume 87, pp. 117–139. ISBN 978-981-10-7757-9. [Google Scholar]
- Pöyry, S.; Róg, T.; Karttunen, M.; Vattulainen, I. Mitochondrial membranes with mono- and divalent salt: Changes induced by salt ions on structure and dynamics. J. Phys. Chem. B 2009, 113, 15513–15521. [Google Scholar] [CrossRef] [PubMed]
- Vähäheikkilä, M.; Peltomaa, T.; Róg, T.; Vazdar, M.; Pöyry, S.; Vattulainen, I. How cardiolipin peroxidation alters the properties of the inner mitochondrial membrane? Chem. Phys. Lipids 2018, 214, 15–23. [Google Scholar] [CrossRef]
- Wilson, B.A.; Ramanathan, A.; Lopez, C.F. Cardiolipin-Dependent Properties of Model Mitochondrial Membranes from Molecular Simulations. Biophys. J. 2019, 117, 429–444. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S.E.; Daum, G. Lipids of mitochondria. Prog. Lipid Res. 2013, 52, 590–614. [Google Scholar] [CrossRef] [PubMed]
- Daum, G.; Vance, J.E. Import of lipids into mitochondria. Prog. Lipid Res. 1997, 36, 103–130. [Google Scholar] [CrossRef]
- Wu, E.L.; Qi, Y.; Song, K.C.; Klauda, J.B.; Im, W. Preferred Orientations of Phosphoinositides in Bilayers and Their Implications in Protein Recognition Mechanisms. J. Phys. Chem. B 2014, 118, 4315–4325. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.J.; Bevan, D.R. Steered molecular dynamics simulations reveal important mechanisms in reversible monoamine oxidase B inhibition. Biochemistry 2011, 50, 6441–6454. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, T.; Iyer, V.; Im, W. CHARMM GUI: A web based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Klauda, J.B.; Venable, R.M.; Freites, J.A.; O’Connor, J.W.; Tobias, D.J.; Mondragon-Ramirez, C.; Vorobyov, I.; MacKerell, A.D.; Pastor, R.W. Update of the CHARMM All-Atom Additive Force Field for Lipids: Validation on Six Lipid Types. J. Phys. Chem. B 2010, 114, 7830–7843. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kalé, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Brooks, B.R.; Brooks, C.L.; Mackerell, A.D.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.L.; Cheng, X.; Jo, S.; Rui, H.; Song, K.C.; Dávila-Contreras, E.M.; Qi, Y.; Lee, J.; Monje-Galvan, V.; Venable, R.M.; et al. CHARMM-GUI membrane builder toward realistic biological membrane simulations. J. Comput. Chem. 2014, 35, 1997–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Petersen, H.G. Accuracy and efficiency of the particle mesh Ewald method. J. Chem. Phys. 1995, 103, 3668–3679. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. Gromacs: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Kučerka, N.; Nieh, M.-P.; Katsaras, J. Fluid phase lipid areas and bilayer thicknesses of commonly used phosphatidylcholines as a function of temperature. Biochim. Biophys. Acta Biomembr. 2011, 1808, 2761–2771. [Google Scholar] [CrossRef]
- Rand, R.P.; Parsegian, V.A. Hydration forces between phospholipid bilayers. Biochim. Biophys. Acta Rev. Biomembr. 1989, 988, 351–376. [Google Scholar] [CrossRef]
- Vermeer, L.S.; de Groot, B.L.; Réat, V.; Milon, A.; Czaplicki, J. Acyl chain order parameter profiles in phospholipid bilayers: Computation from molecular dynamics simulations and comparison with 2H NMR experiments. Eur. Biophys. J. 2007, 36, 919–931. [Google Scholar] [CrossRef] [Green Version]
- Douliez, J.P.; Léonard, A.; Dufourc, E.J. Restatement of order parameters in biomembranes: Calculation of C-C bond order parameters from C-D quadrupolar splittings. Biophys. J. 1995, 68, 1727–1739. [Google Scholar] [CrossRef] [Green Version]
- Javanainen, M.; Hammaren, H.; Monticelli, L.; Jeon, J.-H.; Miettinen, M.S.; Martinez-Seara, H.; Metzler, R.; Vattulainen, I. Anomalous and normal diffusion of proteins and lipids in crowded lipid membranes. Faraday Discuss. 2013, 161, 397–417. [Google Scholar] [CrossRef]
- Klauda, J.B. Perspective: Computational modeling of accurate cellular membranes with molecular resolution. J. Chem. Phys. 2018, 149, 220901. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Róg, T.; Gurtovenko, A.A.; Vattulainen, I.; Karttunen, M. Role of phosphatidylglycerols in the stability of bacterial membranes. Biochimie 2008, 90, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.N.; Wang, E.; Monje-Galvan, V.; Klauda, J.B. Developing and Testing of Lipid Force Fields with Applications to Modeling Cellular Membranes. Chem. Rev. 2019, 119, 6227–6269. [Google Scholar] [CrossRef]
- Lyubartsev, A.P.; Rabinovich, A.L. Force Field Development for Lipid Membrane Simulations. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2483–2497. [Google Scholar] [CrossRef] [PubMed]
- Róg, T.; Hector, M.S.; Munck, N.; Orešič, M.; Karttunen, M.; Vattulainen, I. Role of cardiolipins in the inner mitochondrial membrane: Insight gained through atom-scale simulations. J. Phys. Chem. B 2009, 113, 3413–3422. [Google Scholar] [CrossRef] [PubMed]
- Blosser, M.C.; Starr, J.B.; Turtle, C.W.; Ashcraft, J.; Keller, S.L. Minimal Effect of Lipid Charge on Membrane Miscibility Phase Behavior in Three Ternary Systems. Biophys. J. 2013, 104, 2629–2638. [Google Scholar] [CrossRef] [Green Version]
- Kulig, W.; Korolainen, H.; Zatorska, M.; Kwolek, U.; Kepczynski, M.; Róg, T. Complex Behavior of Phosphatidylcholine−Phosphatidic Acid Bilayers and Monolayers: Effect of Acyl Chain Unsaturation. Langmuir 2019, 35, 5944–5956. [Google Scholar] [CrossRef] [PubMed]
- Pasenkiewicz-Gierula, M.; Baczynski, K.; Markiewicz, M.; Murzyn, K. Computer modelling studies of the bilayer/water interface. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2305–2321. [Google Scholar] [CrossRef]
- Murzyn, K.; Zhao, W.; Karttunen, M.; Kurdziel, M.; Róg, T. Dynamics of water at membrane surfaces: Effect of headgroup structure. Biointerphases 2006, 1, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Murzyn, K.; Róg, T.; Pasenkiewicz-Gierula, M. Phosphatidylethanolamine-phosphatidylglycerol bilayer as a model of the inner bacterial membrane. Biophys. J. 2005, 88, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Karathanou, K.; Bondar, A.-N. Dynamic Water Hydrogen-Bond Networks at the Interface of a Lipid Membrane Containing Palmitoyl-Oleoyl Phosphatidylglycerol. J. Membr. Biol. 2018, 251, 461–473. [Google Scholar] [CrossRef]
- Sarkar, P.; Chattopadhyay, A. Cholesterol interaction motifs in G protein-coupled receptors: Slippery hot spots? WIREs Syst. Biol. Med. 2020, 12, e1481. [Google Scholar] [CrossRef]
- Manna, M.; Niemelä, M.; Tynkkynen, J.; Javanainen, M.; Kulig, W.; Müller, D.J.; Rog, T.; Vattulainen, I. Mechanism of allosteric regulation of β2-adrenergic receptor by cholesterol. Elife 2016, 5, e18432. [Google Scholar] [CrossRef] [PubMed]
- Guixà-González, R.; Albasanz, J.L.; Rodriguez-Espigares, I.; Pastor, M.; Sanz, F.; Martí-Solano, M.; Manna, M.; Martinez-Seara, H.; Hildebrand, P.W.; Martín, M.; et al. Membrane cholesterol access into a G-protein-coupled receptor. Nat. Commun. 2017, 8, 14505. [Google Scholar] [CrossRef] [Green Version]
- Guixà-González, R.; Javanainen, M.; Gómez-Soler, M.; Cordobilla, B.; Domingo, J.C.; Sanz, F.; Pastor, M.; Ciruela, F.; Martinez-Seara, H.; Selent, J. Membrane omega-3 fatty acids modulate the oligomerisation kinetics of adenosine A2A and dopamine D2 receptors. Sci. Rep. 2016, 6, 19839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javanainen, M.; Enkavi, G.; Guixà-Gonzaléz, R.; Kulig, W.; Martinez-Seara, H.; Levental, I.; Vattulainen, I. Reduced level of docosahexaenoic acid shifts GPCR neuroreceptors to less ordered membrane regions. PLOS Comput. Biol. 2019, 15, e1007033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, B.D.; Shrivastava, S.; Chattopadhyay, A. Hydrophobic Mismatch in Membranes: When the Tail Matters BT. In Membrane Organization and Dynamics; Chattopadhyay, A., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 375–387. ISBN 978-3-319-66601-3. [Google Scholar]
- Weiss, T.M.; van der Wel, P.C.A.; Killian, J.A.; Koeppe, R.E., II; Huang, H.W. Hydrophobic Mismatch between Helices and Lipid Bilayers. Biophys. J. 2003, 84, 379–385. [Google Scholar] [CrossRef] [Green Version]
- de Planque, M.R.R.; Killian, J.A. Protein–lipid interactions studied with designed transmembrane peptides: Role of hydrophobic matching and interfacial anchoring (Review). Mol. Membr. Biol. 2003, 20, 271–284. [Google Scholar] [CrossRef]
- Lingwood, D.; Binnington, B.; Róg, T.; Vattulainen, I.; Grzybek, M.; Coskun, Ü.; Lingwood, C.A.; Simons, K. Cholesterol modulates glycolipid conformation and receptor activity. Nat. Chem. Biol. 2011, 7, 260–262. [Google Scholar] [CrossRef]
- Bilkova, E.; Pleskot, R.; Rissanen, S.; Sun, S.; Czogalla, A.; Cwiklik, L.; Róg, T.; Vattulainen, I.; Cremer, P.S.; Jungwirth, P.; et al. Calcium Directly Regulates Phosphatidylinositol 4,5-Bisphosphate Headgroup Conformation and Recognition. J. Am. Chem. Soc. 2017, 139, 4019–4024. [Google Scholar] [CrossRef]
- Ramsay, R.R. Molecular aspects of monoamine oxidase B. Prog. Neuro-Psychopharmacology Biol. Psychiatry 2016, 69, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Lolicato, F.; Juhola, H.; Zak, A.; Postila, P.A.; Saukko, A.; Rissanen, S.; Enkavi, G.; Vattulainen, I.; Kepczynski, M.; Róg, T. Membrane-Dependent Binding and Entry Mechanism of Dopamine into Its Receptor. ACS Chem. Neurosci. 2020, 11, 1914–1924. [Google Scholar] [CrossRef]
- Apostolov, R.; Yonezawa, Y.; Standley, D.M.; Kikugawa, G.; Takano, Y.; Nakamura, H. Membrane attachment facilitates ligand access to the active site in monoamine oxidase A. Biochemistry 2009, 48, 5864–5873. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.B.L.; Crean, R.M.; Mullen, A.; Kendrick, E.G.; Bull, S.D.; Wells, S.A.; Carbery, D.R.; Macmillan, F.; Van Der Kamp, M.W.; Pudney, C.R. Exposing the Interplay between Enzyme Turnover, Protein Dynamics, and the Membrane Environment in Monoamine Oxidase B. Biochemistry 2019, 58, 2362–2372. [Google Scholar] [CrossRef] [PubMed]
- Navrátilová, V.; Paloncýová, M.; Kajšová, M.; Berka, K.; Otyepka, M. Effect of cholesterol on the structure of membrane-attached cytochrome P450 3A4. J. Chem. Inf. Model. 2015, 55, 628–635. [Google Scholar] [CrossRef]
- Navrátilová, V.; Paloncýová, M.; Berka, K.; Otyepka, M. Effect of Lipid Charge on Membrane Immersion of Cytochrome P450 3A4. J. Phys. Chem. B 2016, 120, 11205–11213. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phospholipid Percentage | Model #1 | Model #2 | Model #3 | Model #4 | Model #5 |
|---|---|---|---|---|---|
| POPC | 59% | 57% | 50% | 43% | 57% |
| POPE | 31% | 31% | 24% | 17% | 43% |
| POPS | 0% | 2% | 20% | 39% | 0% |
| PI(3,4)P2 | 10% | 10% | 6% | 1% | 0% |
| Model #1 | Model #2 | Model #3 | Model #4 | Model #5 | |
|---|---|---|---|---|---|
| Area per lipid [nm2] ± 0.02 | 0.621 | 0.620 | 0.617 | 0.616 | 0.612 |
| Total charge of lipids [e] | −312 | −328 | −332 | −338 | 0 |
| Surface charge density [e/nm2] | −0.320 | −0.335 | −0.345 | −0.350 | 0 |
| Total charge (lipids and Na+) [e] | −104 | −104 | −157 | −213 | 18 |
| Thickness [nm] ± 0.05 | 4.06 | 4.13 | 4.10 | 4.13 | 4.19 |
| Model #1 | Model #2 | Model #3 | Model #4 | Model #5 | |
|---|---|---|---|---|---|
| POPC | 8.4 (±0.7) | 6.3 (±0.6) | 7.2 (±0.7) | 6.1 (±0.1) | 8.4 (±0.7) |
| POPE | 8.6 (±0.9) | 6.2 (±0.7) | 7.4 (±0.1) | 5.6 (±0.7) | 9.1 (±0.6) |
| POPS | N/A | 3.5 (±1.4) | 6.3 (±0.9) | 2.9 (±2.9) | N/A |
| PI(3,4)P2 | 8.1 (±1.9) | 4.8 (±3.2) | 5.9 (±0.6) | 5.1 (±0.3) | N/A |
| Model #1 | Model #2 | Model #3 | Model #4 | Model #5 | |
|---|---|---|---|---|---|
| POPC | 6.92 | 6.91 | 6.90 | 6.91 | 6.83 |
| POPE | 7.55 | 7.59 | 7.45 | 7.53 | 7.95 |
| POPS | N/A | 13.44 | 12.98 | 12.79 | N/A |
| PI(3,4)P2 | 20.01 | 19.26 | 20.13 | 20.63 | N/A |
| Partner Lipid | Model #1 | Model #2 | Model #3 | Model #4 | Model #5 |
|---|---|---|---|---|---|
| H-bonds formed by POPE per POPE molecule | |||||
| POPC | 0.48 [0.82] | 0.41 [0.76] | 0.38 [0.80] | 0.32 [0.75] | 0.49 [0.58] |
| POPE | 0.45 [0.45] | 0.45 [0.42] | 0.32 [0.38] | 0.24 [0.30] | 0.53 [0.42] |
| POPS | N/A | 0.04 [0.03] | 0.59 [0.32] | 1.14 [0.68] | N/A |
| PI(3,4)P2 | 0.48 [0.14] | 0.44 [0.13] | 0.31 [0.10] | 0.05 [0.02] | N/A |
| All | 1.41 | 1.34 | 1.60 | 1.75 | 1.02 |
| H-bonds formed by POPS per POPS molecule | |||||
| POPC | N/A | 0.44 [0.76] | 0.29 [0.79] | 0.23 [0.65] | N/A |
| POPE | N/A | 0.62 [0.42] | 0.73 [0.38] | 0.49 [0.26] | N/A |
| POPS | N/A | 0.27 [0.03] | 0.42 [0.31] | 0.76 [0.59] | N/A |
| PI(3,4)P2 | N/A | 0.01 [0.13] | 0.13 [0.09] | 0.03 [0.01] | N/A |
| All | N/A | 1.34 | 1.57 | 1.51 | N/A |
| H-bonds formed by PI(3,4)P2 per PI(3,4)P2 molecule | |||||
| POPC | 0.17 | 0.17 | 0.12 | 0.16 | N/A |
| POPE | 1.49 | 1.37 | 1.36 | 1.08 | N/A |
| POPS | N/A | 0.00 | 0.45 | 1.08 | N/A |
| PI(3,4)P2 | 1.86 | 1.86 | 1.86 | 1.98 | N/A |
| All | 3.52 | 3.40 | 3.79 | 4.3 | N/A |
| H-bonds formed by POPC per POPC molecule | |||||
| POPE | 0.25 [0.21] | 0.22 [0.19] | 0.18 [0.14] | 0.12 [0.11] | 0.36 |
| POPS | N/A | 0.02 [0.02] | 0.11 [0.12] | 0.21 [0.22] | N/A |
| PI(3,4)P2 | 0.03 [0.07] | 0.03 [0.06] | 0.01 [0.02] | 0.00 [0.00] | N/A |
| All | 0.28 | 0.27 | 0.30 | 0.33 | 0.36 |
| Charge pairs formed by POPC per POPC molecule (or POPC per partner lipid) | |||||
| POPC | 1.89 | 1.84 | 1.66 | 1.52 | 1.75 |
| POPE | 0.69 (1.33) * | 0.69 (1.33) * | 0.56 (1.15) | 0.39 (0.69) | 0.94 (1.25) |
| POPS | N/A * | N/A * | 0.74 (1.88) | 1.29 (1.42) | N/A |
| PI(3,4)P2 | 0.70 (4.14) | 0.69 (3.94) | 0.40 (3.55) | 0.1 (4.25) | N/A |
| All | 3.28 * | 3.28 * | 3.36 | 3.30 | 2.69 |
| Model #1 | Model #2 | Model #3 | Model #4 | Model #5 | |
|---|---|---|---|---|---|
| Number of Na+ bound per lipid | 0.27 | 0.29 | 0.22 | 0.16 | 0.02 |
| Multiplicity | 3.14 | 3.24 | 2.49 | 1.80 | 1.56 |
| Number of Na+ bound per lipid—carbonyl groups | 0.05 | 0.10 | 0.09 | 0.08 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, A.A.; Róg, T.; da Silva, A.B.F.; Amaro, R.E.; Johnson, M.S.; Postila, P.A. Examining the Effect of Charged Lipids on Mitochondrial Outer Membrane Dynamics Using Atomistic Simulations. Biomolecules 2022, 12, 183. https://doi.org/10.3390/biom12020183
Oliveira AA, Róg T, da Silva ABF, Amaro RE, Johnson MS, Postila PA. Examining the Effect of Charged Lipids on Mitochondrial Outer Membrane Dynamics Using Atomistic Simulations. Biomolecules. 2022; 12(2):183. https://doi.org/10.3390/biom12020183
Chicago/Turabian StyleOliveira, Aline A., Tomasz Róg, Albérico B. F. da Silva, Rommie E. Amaro, Mark S. Johnson, and Pekka A. Postila. 2022. "Examining the Effect of Charged Lipids on Mitochondrial Outer Membrane Dynamics Using Atomistic Simulations" Biomolecules 12, no. 2: 183. https://doi.org/10.3390/biom12020183
APA StyleOliveira, A. A., Róg, T., da Silva, A. B. F., Amaro, R. E., Johnson, M. S., & Postila, P. A. (2022). Examining the Effect of Charged Lipids on Mitochondrial Outer Membrane Dynamics Using Atomistic Simulations. Biomolecules, 12(2), 183. https://doi.org/10.3390/biom12020183