
Biologically Relevant In Vitro 3D-Model to Study Bone Regeneration Potential of Human Adipose Stem Cells

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Platelet Lysate
2.2. Adipose Stem Cells
2.3. Culture of hASCs
2.4. Treatment of hASCs with Cytokines
2.5. Protein and DNA
2.6. AlamarBlue Assay
2.7. RNA Isolation and Real-Time PCR
2.8. ALP Activity
2.9. Mineralization
2.10. Statistical Analysis
3. Results
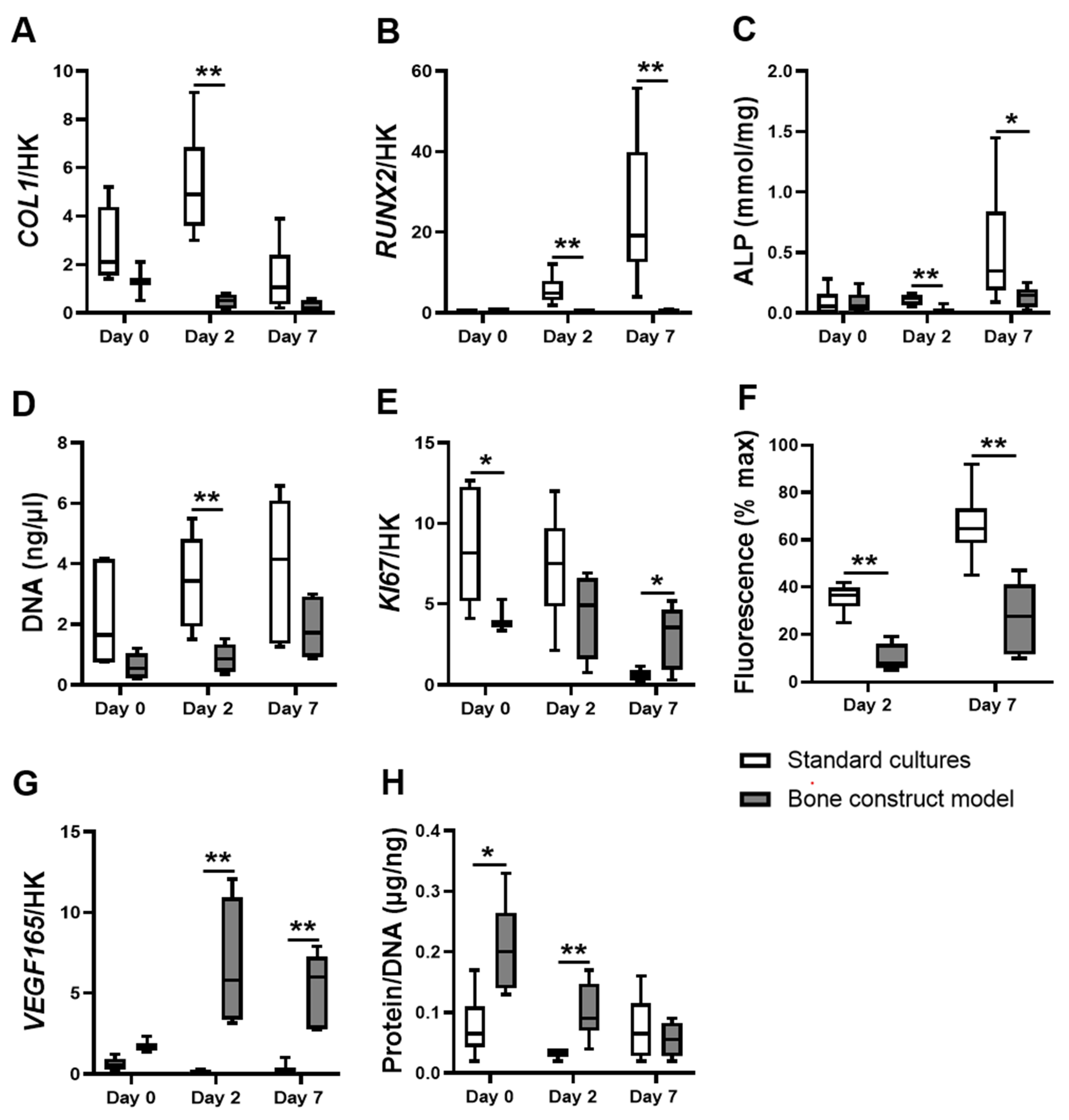
3.1. hASCs in the Bone Construct Model Showed Altered Proliferation but Decreased Differentiation and Metabolic Activity Compared to hASCs in Standard Cultures
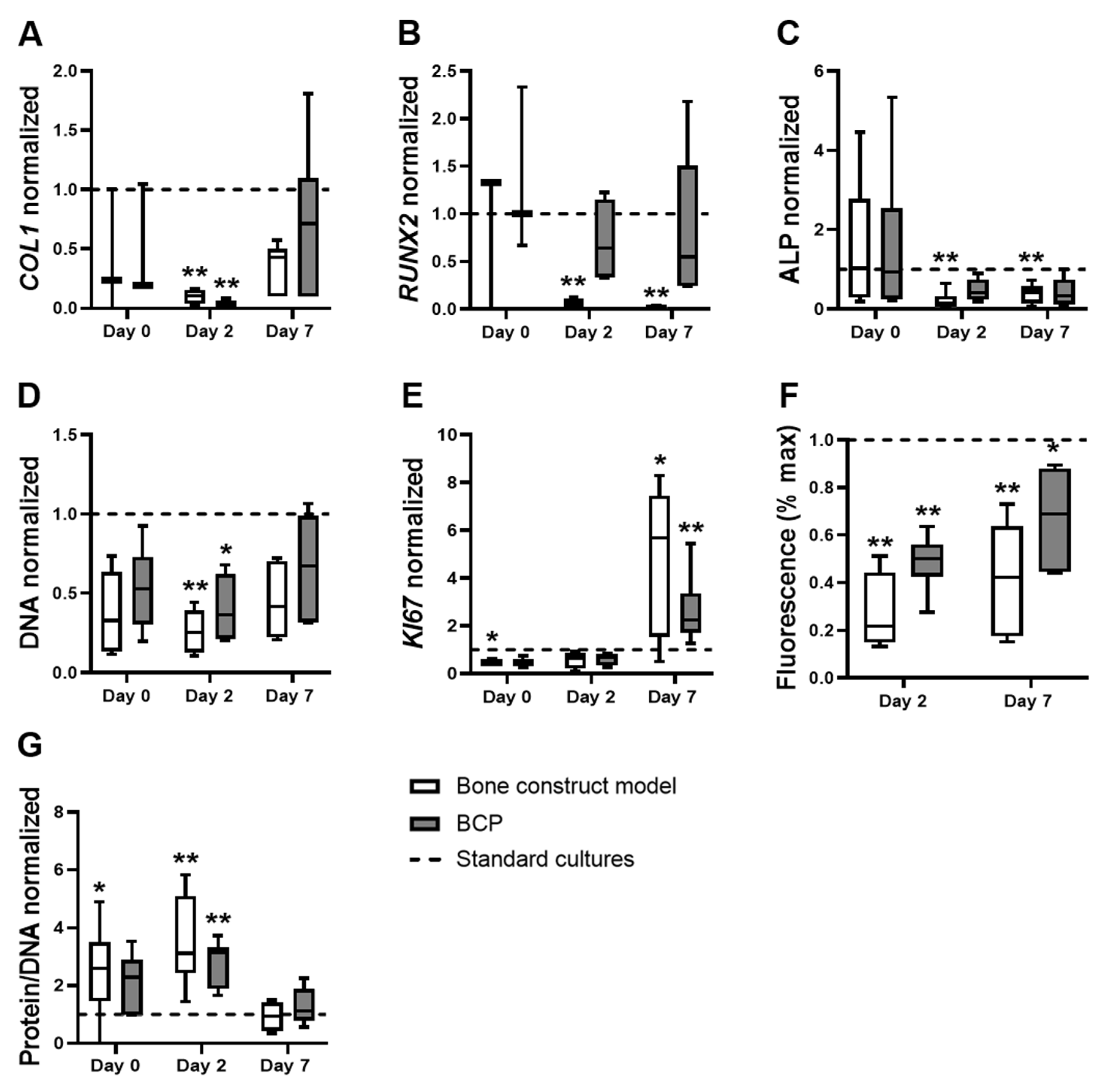
3.2. The Effects of the Bone Construct Model on hASC Proliferation, Metabolic Activity and Total Protein Were Comparable to hASCs on BCP under Normoxia, without Cytokines
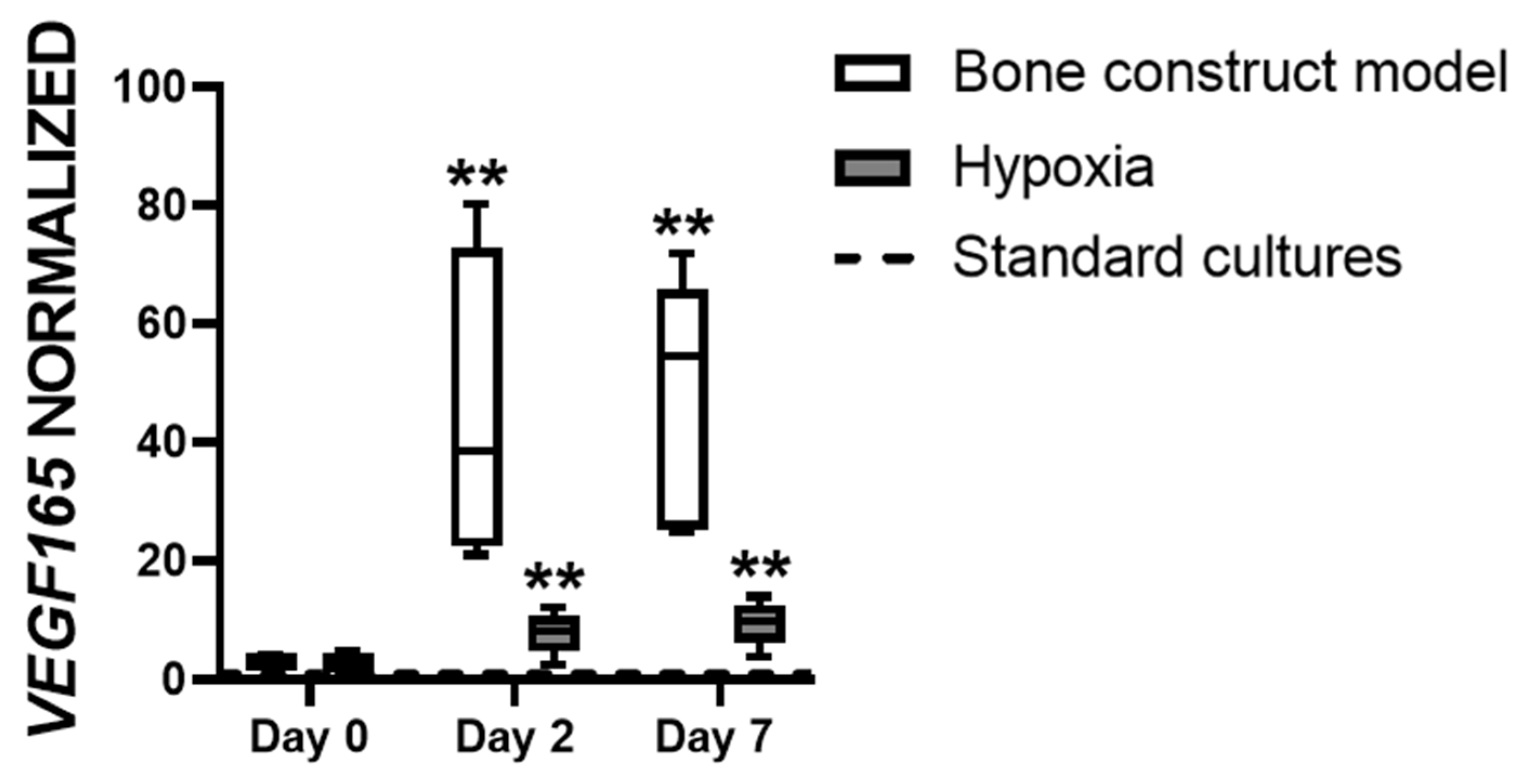
3.3. Hypoxia Affects VEGF165 Expression in hASCs
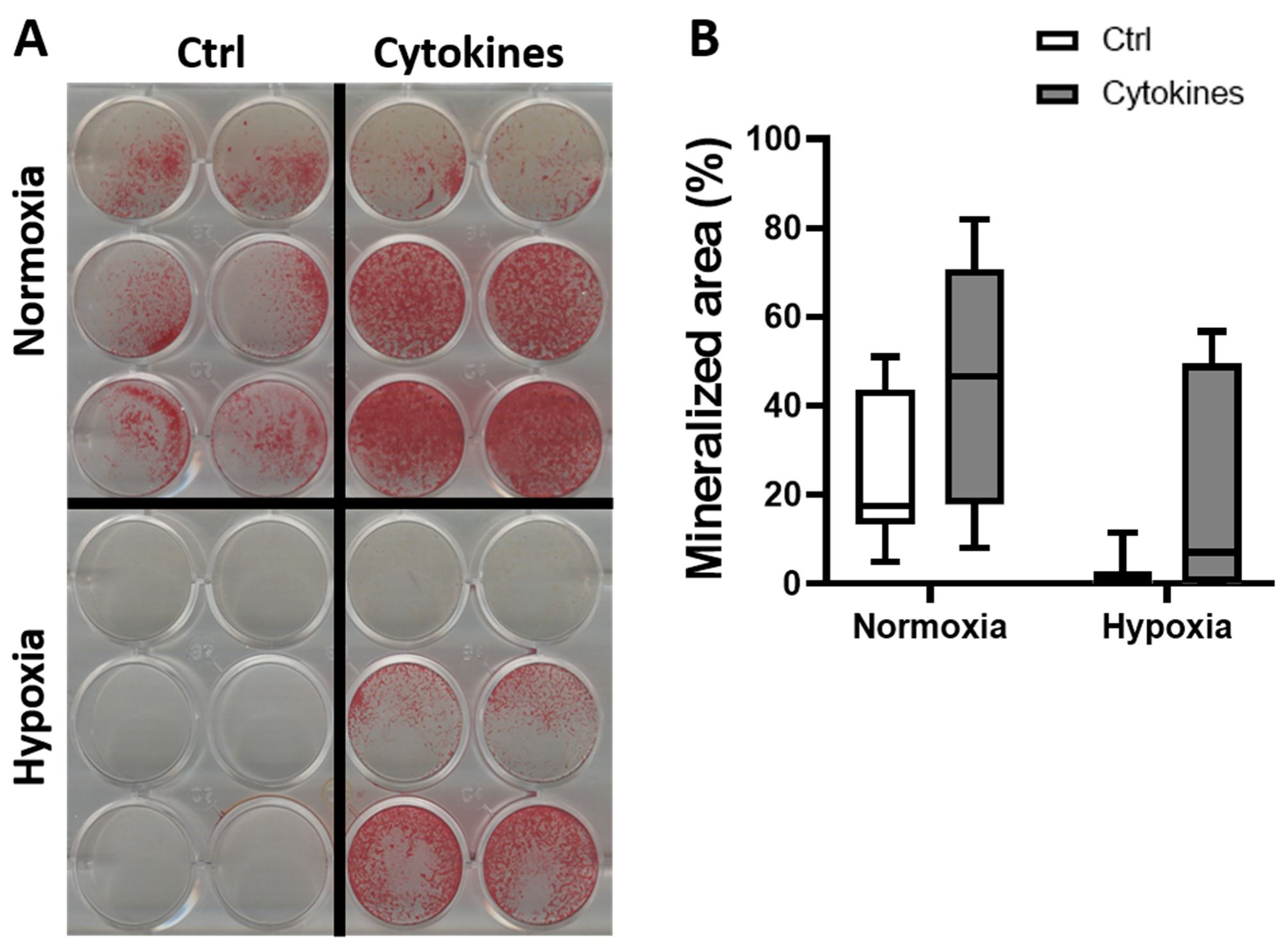
3.4. A Cytokine Cocktail Does Not Decrease hASC Differentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krebsbach, P.H.; Kuznetsov, S.A.; Bianco, P.; Robey, P.G. Bone marrow stromal cells: Characterization and clinical application. Crit. Rev. Oral Biol. Med. 1999, 10, 165–181. [Google Scholar] [CrossRef]
- Tuan, R.S.; Boland, G.; Tuli, R. Adult mesenchymal stem cells and cell-based tissue engineering. Arthritis Res. Ther. 2003, 5, 32–45. [Google Scholar] [CrossRef]
- Farré-Guasch, E.; Bravenboer, N.; Helder, M.N.; Schulten, E.A.J.M.; ten Bruggenkate, C.M.; Klein-Nulend, J. Blood vessel formation and bone regeneration potential of the stromal vascular fraction seeded on a calcium phosphate scaffold in the human maxillary sinus floor elevation model. Materials 2018, 11, 161. [Google Scholar] [CrossRef]
- Overman, J.R.; Farré-Guasch, E.; Helder, M.N.; Ten Bruggenkate, C.M.; Schulten, E.A.J.M.; Klein-Nulend, J. Short (15 minutes) bone morphogenetic protein-2 treatment stimulates osteogenic differentiation of human adipose stem cells seeded on calcium phosphate scaffolds in vitro. Tissue Eng. Part A 2013, 19, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Meyerson, H.; Anderson, J.M. Quantitative in vivo cytokine analysis at synthetic biomaterial implant sites. J. Biomed. Mater. Res. Part A 2009, 89, 152–159. [Google Scholar] [CrossRef]
- Bollati, D.; Morra, M.; Cassinelli, C.; Lupi, S.M.; Baena, R.R.Y. In Vitro Cytokine Expression and In Vivo Healing and Inflammatory Response to a Collagen-Coated Synthetic Bone Filler. Biomed Res. Int. 2016, 2016, 1–10. [Google Scholar] [CrossRef][Green Version]
- Gao, L.; Li, M.; Yin, L.; Zhao, C.; Chen, J.; Zhou, J.; Duan, K.; Feng, B. Dual-inflammatory cytokines on TiO2 nanotube-coated surfaces used for regulating macrophage polarization in bone implants. J. Biomed. Mater. Res. Part A 2018, 106, 1878–1886. [Google Scholar] [CrossRef] [PubMed]
- Mountziaris, P.M.; Mikos, A.G. Modulation of the inflammatory response for enhanced bone tissue regeneration. Tissue Eng. Part B Rev. 2008, 14, 179–186. [Google Scholar] [CrossRef]
- Marsell, R.; Einhorn, T.A. The biology of fracture healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef]
- Kon, T.; Cho, T.J.; Aizawa, T.; Yamazaki, M.; Nooh, N.; Graves, D.; Gerstenfeld, L.C.; Einhorn, T.A. Expression of osteoprotegerin, receptor activator of NF-κB ligand (osteoprotegerin ligand) and related proinflammatory cytokines during fracture healing. J. Bone Miner. Res. 2001, 16, 1004–1014. [Google Scholar] [CrossRef]
- Al-Aql, Z.S.; Alagl, A.S.; Graves, D.T.; Gerstenfeld, L.C.; Einhorn, T.A. Molecular mechanisms controlling bone formation during fracture healing and distraction osteogenesis. J. Dent. Res. 2008, 87, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Toben, D.; Schroeder, I.; El Khassawna, T.; Mehta, M.; Hoffmann, J.E.; Frisch, J.T.; Schell, H.; Lienau, J.; Serra, A.; Radbruch, A.; et al. Fracture healing is accelerated in the absence of the adaptive immune system. J. Bone Miner. Res. 2011, 26, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Cho, T.J.; Gerstenfeld, L.C.; Einhorn, T.A. Differential temporal expression of members of the transforming growth factor β superfamily during murine fracture healing. J. Bone Miner. Res. 2002, 17, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Mau, E.; Wang, Y.; Wright, D.; Silkstone, D.; Whetstone, H.; Whyne, C.; Alman, B. T-lymphocytes enable osteoblast maturation via IL-17F during the early phase of fracture repair. PLoS ONE 2012, 7, e40044. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Cho, T.J.; Kon, T.; Aizawa, T.; Cruceta, J.; Graves, B.D.; Einhorn, T.A. Impaired intramembranous bone formation during bone repair in the absence of tumor necrosis factor-alpha signaling. Cells Tissues Organs 2001, 169, 285–294. [Google Scholar] [CrossRef]
- Woodward, E.A.; Kolesnik, T.B.; Nicholson, S.E.; Prêle, C.M.; Hart, P.H. The anti-inflammatory actions of IL-4 in human monocytes are not mediated by IL-10, RP105 or the kinase activity of RIPK2. Cytokine 2012, 58, 415–423. [Google Scholar] [CrossRef]
- Mangashetti, L.S.; Khapli, S.M.; Wani, M.R. IL-4 Inhibits Bone-Resorbing Activity of Mature Osteoclasts by Affecting NF-κB and Ca 2+ Signaling. J. Immunol. 2005, 175, 917–925. [Google Scholar] [CrossRef]
- Wallace, A.; Cooney, T.E.; Englund, R.; Lubahn, J.D. Effects of interleukin-6 ablation on fracture healing in mice. J. Orthop. Res. 2011, 29, 1437–1442. [Google Scholar] [CrossRef]
- Marenzana, M.; Arnett, T.R. The Key Role of the Blood Supply to Bone. Bone Res. 2013, 1, 203–215. [Google Scholar] [CrossRef]
- Maes, C.; Carmeliet, G.; Schipani, E. Hypoxia-driven pathways in bone development, regeneration and disease. Nat. Rev. Rheumatol. 2012, 8, 358–366. [Google Scholar] [CrossRef]
- Weijers, E.M.; Broek, L.J.V.D.; Waaijman, T.; Van Hinsbergh, V.W.M.; Gibbs, S.; Koolwijk, P. The influence of hypoxia and fibrinogen variants on the expansion and differentiation of adipose tissue-derived mesenchymal stem cells. Tissue Eng. Part A 2011, 17, 2675–2685. [Google Scholar] [CrossRef]
- van Esterik, F.A.S.; Hogervorst, J.M.A.; Koolwijk, P.; Klein-Nulend, J. Hypoxia decreases osteogenic differentiation potential but enhances vasculogenic differentiation potential of human adipose stem cells on biphasic calcium phosphate scaffolds in fibrin gels. J. Stem Cells 2017, 12, 41–52. [Google Scholar]
- Leegwater, N.C.; Bakker, A.D.; Hogervorst, J.M.A.; Nolte, P.A.; Klein-Nulend, J. Hypothermia reduces VEGF-165 expression, but not osteogenic differentiation of human adipose stem cells under hypoxia. PLoS ONE 2017, 12, e0171492. [Google Scholar] [CrossRef]
- Fuchs, R.K.; Warden, S.J.; Turner, C.H. Bone anatomy, physiology and adaptation to mechanical loading. In Bone Repair Biomaterials; Elsevier: Amsterdam, The Netherlands, 2009; pp. 25–68. ISBN 9781845693855. [Google Scholar]
- Nasello, G.; Alamán-Díez, P.; Schiavi, J.; Pérez, M.Á.; McNamara, L.; García-Aznar, J.M. Primary Human Osteoblasts Cultured in a 3D Microenvironment Create a Unique Representative Model of Their Differentiation Into Osteocytes. Front. Bioeng. Biotechnol. 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Bulnheim, U.; Diener, A.; Lüthen, F.; Teller, M.; Klinkenberg, E.D.; Neumann, H.G.; Nebe, B.; Liebold, A.; Steinhoff, G.; et al. Calcium phosphate surfaces promote osteogenic differentiation of mesenchymal stem cells. J. Cell Mol. Med. 2008, 12, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Strioga, M.; Viswanathan, S.; Darinskas, A.; Slaby, O.; Michalek, J. Same or not the same? comparison of adipose tissue-derived versus bone marrow-derived mesenchymal stem and stromal cells. Stem Cells Dev. 2012, 21, 2724–2752. [Google Scholar] [CrossRef] [PubMed]
- Barba, M.; Di Taranto, G.; Lattanzi, W. Adipose-derived stem cell therapies for bone regeneration. Expert Opin. Biol. Ther. 2017, 17, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Prins, H.-J.; Schulten, E.A.J.M.; ten Bruggenkate, C.M.; Klein-Nulend, J.; Helder, M.N. Bone Regeneration Using the Freshly Isolated Autologous Stromal Vascular Fraction of Adipose Tissue in Combination With Calcium Phosphate Ceramics. Stem Cells Transl. Med. 2016, 5, 1362–1374. [Google Scholar] [CrossRef]
- Varma, M.J.O.; Breuls, R.G.M.; Schouten, T.E.; Jurgens, W.J.F.M.; Bontkes, H.J.; Schuurhuis, G.J.; Van Ham, S.M.; Van Milligen, F.J. Phenotypical and functional characterization of freshly isolated adipose tissue-derived stem cells. Stem Cells Dev. 2007, 16, 91–104. [Google Scholar] [CrossRef]
- Ma, J.; van den Beucken, J.J.J.P.; Both, S.K.; Prins, H.J.; Helder, M.N.; Yang, F.; Jansen, J.A. Osteogenic capacity of human BM-MSCs, AT-MSCs and their co-cultures using HUVECs in FBS and PL supplemented media. J. Tissue Eng. Regen. Med. 2015, 9, 779–788. [Google Scholar] [CrossRef]
- Jurgens, W.J.; Kroeze, R.J.; Bank, R.A.; Ritt, M.J.P.F.; Helder, M.N. Rapid attachment of adipose stromal cells on resorbable polymeric scaffolds facilitates the one-step surgical procedure for cartilage and bone tissue engineering purposes. J. Orthop. Res. 2011, 29, 853–860. [Google Scholar] [CrossRef]
- Langenbach, F.; Handschel, J. Effects of dexamethasone, ascorbic acid and β-glycerophosphate on the osteogenic differentiation of stem cells in vitro. Stem Cell Res. Ther. 2013, 4, 117. [Google Scholar] [CrossRef]
- Nauta, T.D.; Duyndam, M.C.A.; Weijers, E.M.; Van Hinsbergh, V.M.W.; Koolwijk, P. HIF-2α expression regulates sprout formation into 3d fibrin matrices in prolonged hypoxia in human microvascular endothelial cells. PLoS ONE 2016, 11, e0160700. [Google Scholar] [CrossRef][Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H. [17] Micromethods for the assay of enzymes. Methods Enzymol. 1957, 4, 366–381. [Google Scholar] [CrossRef]
- Fukuyo, S.; Yamaoka, K.; Sonomoto, K.; Oshita, K.; Okada, Y.; Saito, K.; Yoshida, Y.; Kanazawa, T.; Minami, Y.; Tanaka, Y. IL-6-accelerated calcification by induction of ROR2 in human adipose tissue-derived mesenchymal stem cells is STAT3 dependent. Rheumatology 2014, 53, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.M.; Stern, M.M.; Stern, A.R.; Pathak, J.L.; Bravenboer, N.; Bakker, A.D. Osteocyte isolation and culture methods. Bonekey Rep. 2016, 5, 838. [Google Scholar] [CrossRef][Green Version]
- Volpin, G.; Cohen, M.; Assaf, M.; Meir, T.; Katz, R.; Pollack, S. Cytokine levels (IL-4, IL-6, IL-8 and TGFβ) as potential biomarkers of systemic inflammatory response in trauma patients. Int. Orthop. 2014, 38, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Miramini, S.; Patel, M.; Richardson, M.; Ebeling, P.; Zhang, L. Role of TNF-α in early-stage fracture healing under normal and diabetic conditions. Comput. Methods Programs Biomed. 2022, 213, 106536. [Google Scholar] [CrossRef]
- Lord, J.M.; Midwinter, M.J.; Chen, Y.F.; Belli, A.; Brohi, K.; Kovacs, E.J.; Koenderman, L.; Kubes, P.; Lilford, R.J. The systemic immune response to trauma: An overview of pathophysiology and treatment. Lancet 2014, 384, 1455–1465. [Google Scholar] [CrossRef]
- Kolar, P.; Gaber, T.; Perka, C.; Duda, G.N.; Buttgereit, F. Human early fracture hematoma is characterized by inflammation and hypoxia. Clin. Orthop. Relat. Res. 2011, 469, 3118–3126. [Google Scholar] [CrossRef] [PubMed]
- Oe, K.; Miwa, M.; Sakai, Y.; Lee, S.Y.; Kuroda, R.; Kurosaka, M. An in vitro study demonstrating that haematomas found at the site of human fractures contain progenitor cells with multilineage capacity. J. Bone Jt. Surg. Ser. B 2007, 89, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Bleek, K.; Schell, H.; Lienau, J.; Schulz, N.; Hoff, P.; Pfaff, M.; Schmidt, G.; Martin, C.; Perka, C.; Buttgereit, F.; et al. Initial immune reaction and angiogenesis in bone healing. J. Tissue Eng. Regen. Med. 2014, 8, 120–130. [Google Scholar] [CrossRef]
- Loi, F.; Córdova, L.A.; Pajarinen, J.; Lin, T.H.; Yao, Z.; Goodman, S.B. Inflammation, fracture and bone repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.C.; Raggatt, L.J.; Alexander, K.A.; Pettit, A.R. Unraveling macrophage contributions to bone repair. Bonekey Rep. 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Saless, N.; Wang, X.; Sinha, A.; Decker, S.; Kazakia, G.; Hou, H.; Williams, B.; Swartz, H.M.; Hunt, T.K.; et al. The role of oxygen during fracture healing. Bone 2013, 52, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Wenger, R.; Kurtcuoglu, V.; Scholz, C.; Marti, H.; Hoogewijs, D. Frequently asked questions in hypoxia research. Hypoxia 2015, 18, 35–43. [Google Scholar] [CrossRef]
- Yang, D.C.; Yang, M.H.; Tsai, C.C.; Huang, T.F.; Chen, Y.H.; Hung, S.C. Hypoxia inhibits osteogenesis in human mesenchymal stem cells through direct regulation of RUNX2 by TWIST. PLoS ONE 2011, 6, e23965. [Google Scholar] [CrossRef] [PubMed]
- Steinbrech, D.S.; Mehrara, B.J.; Saadeh, P.B.; Chin, G.; Dudziak, M.E.; Gerrets, R.P.; Gittes, G.K.; Longaker, M.T. Hypoxia regulates VEGF expression and cellular proliferation by osteoblasts in vitro. Plast. Reconstr. Surg. 1999, 104, 737–747. [Google Scholar] [CrossRef]
- Lacey, D.C.; Simmons, P.J.; Graves, S.E.; Hamilton, J.A. Proinflammatory cytokines inhibit osteogenic differentiation from stem cells: Implications for bone repair during inflammation. Osteoarthr. Cartil. 2009, 17, 735–742. [Google Scholar] [CrossRef]
- Peruzzi, B.; Cappariello, A.; Del Fattore, A.; Rucci, N.; De Benedetti, F.; Teti, A. C-Src and IL-6 inhibit osteoblast differentiation and integrate IGFBP5 signalling. Nat. Commun. 2012, 3, 630. [Google Scholar] [CrossRef] [PubMed]
- Riancho, J.A.; Zarrabeitia, M.T.; Olmos, J.M.; Amado, J.A.; Gonzalez-Macias, J. Effects of interleukin-4 on human osteoblast-like cells. Bone Miner. 1993, 21, 53–61. [Google Scholar] [CrossRef]
- Croes, M.; Öner, F.C.; van Neerven, D.; Sabir, E.; Kruyt, M.C.; Blokhuis, T.J.; Dhert, W.J.A.; Alblas, J. Proinflammatory T cells and IL-17 stimulate osteoblast differentiation. Bone 2016, 84, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, D.; Guo, B.; Yang, X.; Chen, X.; Zhu, X.; Fan, Y.; Zhang, X. Role of biphasic calcium phosphate ceramic-mediated secretion of signaling molecules by macrophages in migration and osteoblastic differentiation of MSCs. Acta Biomater. 2017, 51, 447–460. [Google Scholar] [CrossRef]
- De Lange, G.L.; Overman, J.R.; Farré-Guasch, E.; Korstjens, C.M.; Hartman, B.; Langenbach, G.E.J.; Van Duin, M.A.; Klein-Nulend, J. A histomorphometric and micro-computed tomography study of bone regeneration in the maxillary sinus comparing biphasic calcium phosphate and deproteinized cancellous bovine bone in a human split-mouth model. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, 8–22. [Google Scholar] [CrossRef]
- Street, J.T.; Lenehan, B.; DeSchrijver, K.; Wang, J.H.; Wu, Q.D.; Redmond, H.P. Hypoxia regulates the paracrine coupling of angiogenesis and bone formation. Eur. J. Orthop. Surg. Traumatol. 2005, 15, 214–225. [Google Scholar] [CrossRef]
- Porter, K.M.; Kang, B.Y.; Adesina, S.E.; Murphy, T.C.; Hart, C.M.; Sutliff, R.L. Chronic hypoxia promotes pulmonary artery endothelial cell proliferation through H2O2-induced 5-lipoxygenase. PLoS ONE 2014, 9, e98532. [Google Scholar] [CrossRef]
- Wang, K.; Jiang, Y.-Z.; Chen, D.-B.; Zheng, J. Hypoxia Enhances FGF2- and VEGF-Stimulated Human Placental Artery Endothelial Cell Proliferation: Roles of MEK1/2/ERK1/2 and PI3K/AKT1 Pathways. Placenta 2009, 30, 1045–1051. [Google Scholar] [CrossRef]
- Wang, G.L.; Jiang, B.H.; Rue, E.A.; Semenza, G.L. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. USA 1995, 92, 5510–5514. [Google Scholar] [CrossRef]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-Induced Angiogenesis: Good and Evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef]
- Chen, L.; Endler, A.; Shibasaki, F. Hypoxia and angiogenesis: Regulation of hypoxia-inducible factors via novel binding factors. Exp. Mol. Med. 2009, 41, 849–857. [Google Scholar] [CrossRef]
- Lund, E.L.; Thorsen, C.; Pedersen, M.W.B.; Junker, N.; Kristjansen, P.E.G. Relationship between vessel density and expression of vascular endothelial growth factor and basic fibroblast growth factor in small cell lung cancer in vivo and in vitro. Clin. Cancer Res. 2000, 6, 4287–4291. [Google Scholar]
- Edelman, J.L.; Castro, M.R.; Wen, Y. Correlation of VEGF expression by leukocytes with the growth and regression of blood vessels in the rat cornea. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1112–1123. [Google Scholar]
- Chakkalakal, D.A.; Mashoof, A.A.; Novak, J.; Strates, B.S.; McGuire, M.H. Mineralization and pH relationships in healing skeletal defects grafted with demineralized bone matrix. J. Biomed. Mater. Res. 1994, 28, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Callewaert, F.; Sinnesael, M.; Gielen, E.; Boonen, S.; Vanderschueren, D. Skeletal sexual dimorphism: Relative contribution of sex steroids, GH-IGF1, and mechanical loading. J. Endocrinol. 2010, 207, 127–134. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Human) | Forward | Reverse |
|---|---|---|
| TBP | 5′-GGTCTGGGAAAATGGTGTGC-3′ | 5′-GCTGGAAAACCAACTTCTG-3′ |
| GUSB | 5′-CGCACAAGAGTGGTGCTGAG-3′ | 5′-GGAGGTGTCAGTCAGGTATT-3′ |
| KI67 | 5′ CCCTCAGCAAGCCTGAGAA 3′ | 5′ AGAGGCGTATTAGGAGGCAAG 3′ |
| RUNX2 | 5′ ATGCTTCATTCGCCTCAC 3′ | 5′ ACTGCTTGCAGCCTTAAAT 3′ |
| COL1 | 5′ TCCGGCTCCTGCTCCTCTTA 3′ | 5′ GGCCAGTGTCTCCCTTG 3′ |
| VEGF165 | 5′-ATCTTCAAGCCATCCTGTGTGC-3′ | 5′-CAAGGCCCACAGGGATTTTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Santen, V.J.B.; Bastidas Coral, A.P.; Hogervorst, J.M.A.; Klein-Nulend, J.; Bakker, A.D. Biologically Relevant In Vitro 3D-Model to Study Bone Regeneration Potential of Human Adipose Stem Cells. Biomolecules 2022, 12, 169. https://doi.org/10.3390/biom12020169
van Santen VJB, Bastidas Coral AP, Hogervorst JMA, Klein-Nulend J, Bakker AD. Biologically Relevant In Vitro 3D-Model to Study Bone Regeneration Potential of Human Adipose Stem Cells. Biomolecules. 2022; 12(2):169. https://doi.org/10.3390/biom12020169
Chicago/Turabian Stylevan Santen, Victor J. B., Angela P. Bastidas Coral, Jolanda M. A. Hogervorst, Jenneke Klein-Nulend, and Astrid D. Bakker. 2022. "Biologically Relevant In Vitro 3D-Model to Study Bone Regeneration Potential of Human Adipose Stem Cells" Biomolecules 12, no. 2: 169. https://doi.org/10.3390/biom12020169
APA Stylevan Santen, V. J. B., Bastidas Coral, A. P., Hogervorst, J. M. A., Klein-Nulend, J., & Bakker, A. D. (2022). Biologically Relevant In Vitro 3D-Model to Study Bone Regeneration Potential of Human Adipose Stem Cells. Biomolecules, 12(2), 169. https://doi.org/10.3390/biom12020169