

Molecular Pathways of the Therapeutic Effects of Ayahuasca, a Botanical Psychedelic and Potential Rapid-Acting Antidepressant

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. A Brief Review of the Current Evidence for the Antidepressant Effects of Ayahuasca in Humans
3. Safety and Tolerability of Ayahuasca Administration
4. Ayahuasca’s Alkaloid Content
5. Pharmacokinetics of Ayahuasca
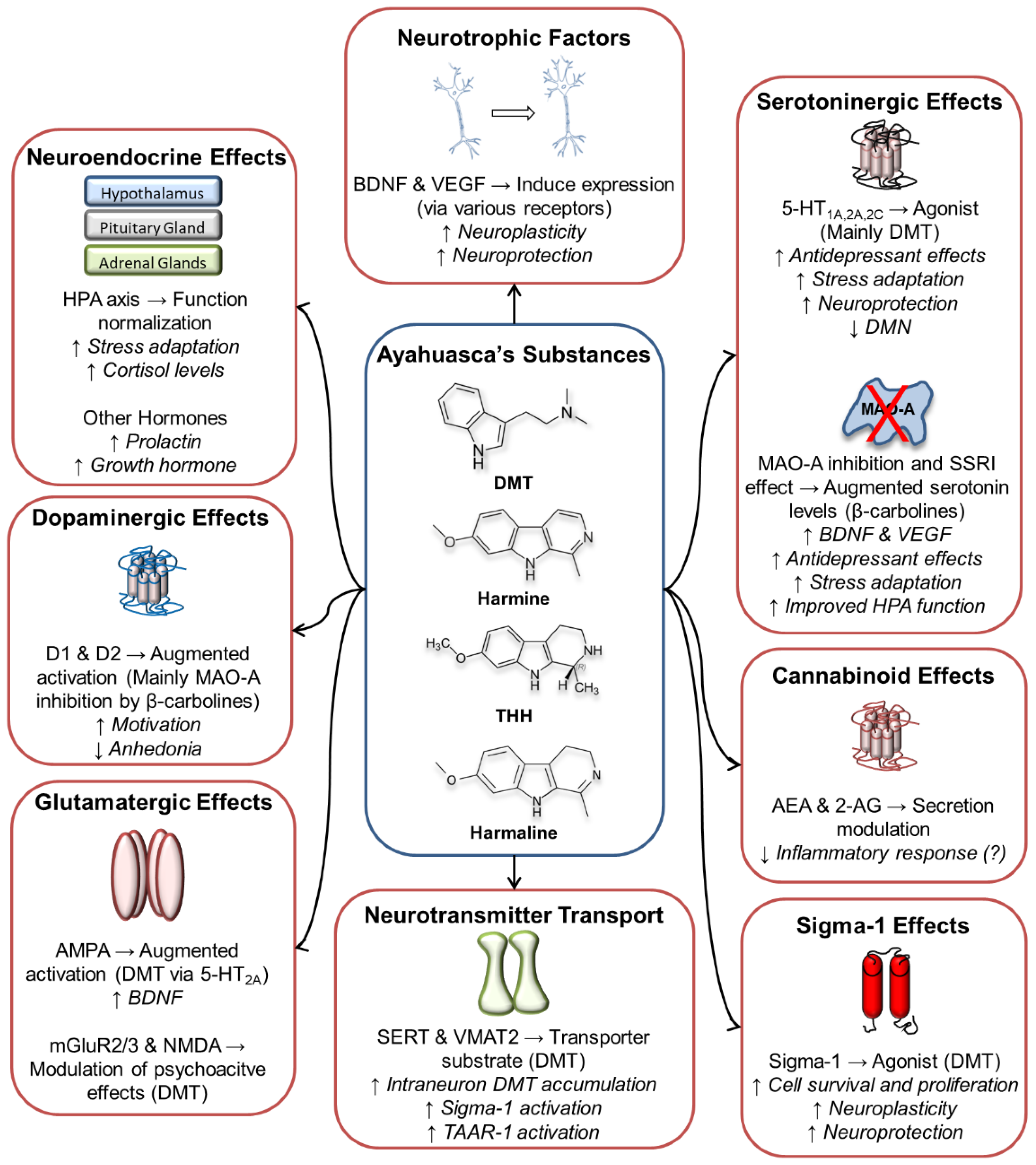
6. Ayahuasca’s Molecular Targets and Their Relation to Possible Antidepressant Effects
6.1. Serotoninergic System
6.2. Glutamatergic System
6.3. Dopaminergic System
6.4. Endocannabinoid System
6.5. Sigma-1 Receptors
6.6. Neuroendocrine System
6.7. Other Molecular Targets/Systems
7. Final Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- dos Santos, R.G.; Valle, M.; Bouso, J.C.; Nomdedéu, J.F.; Rodríguez-Espinosa, J.; McIlhenny, E.H.; Barker, S.A.; Barbanoj, M.J.; Riba, J. Autonomic, Neuroendocrine, and Immunological Effects of Ayahuasca: A Comparative Study With d-Amphetamine. J. Clin. Psychopharmacol. 2011, 31, 717–726. [Google Scholar] [CrossRef]
- dos Santos, R.G.; Osório, F.L.; Crippa, J.A.; Hallak, J.E. Antidepressive and Anxiolytic Effects of Ayahuasca: A Systematic Literature Review of Animal and Human Studies. Braz. J. Psychiatry 2016, 38, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Schultes, R.E.; Ceballos, L.F.; Castillo, A. El desarrollo histórico de la identificación de las malpigiáceas empleadas como alucinógenos. Am. Indig. 1986, 46, 9–47. [Google Scholar] [PubMed]
- Cata-Preta, E.G.; Serra, Y.A.; Moreira-Junior, E.D.C.; Reis, H.S.; Kisaki, N.D.; Libarino-Santos, M.; Silva, R.R.R.; Barros-Santos, T.; Santos, L.C.; Barbosa, P.C.R.; et al. Ayahuasca and Its DMT- and β-Carbolines—Containing Ingredients Block the Expression of Ethanol-Induced Conditioned Place Preference in Mice: Role of the Treatment Environment. Front. Pharmacol. 2018, 9, 561. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.J.; Towers, G.H.; Abbott, F. Monoamine Oxidase Inhibitors in South American Hallucinogenic Plants: Tryptamine and Beta-Carboline Constituents of Ayahuasca. J. Ethnopharmacol. 1984, 10, 195–223. [Google Scholar] [CrossRef]
- Buckholtz, N.S.; Boggan, W.O. Monoamine Oxidase Inhibition in Brain and Liver Produced by Beta-Carbolines: Structure-Activity Relationships and Substrate Specificity. Biochem. Pharmacol. 1977, 26, 1991–1996. [Google Scholar] [CrossRef]
- Knudsen, G.M. Sustained Effects of Single Doses of Classical Psychedelics in Humans. Neuropsychopharm 2022, 1–6. [Google Scholar] [CrossRef]
- Hirschfeld, T.; Schmidt, T.T. Dose-Response Relationships of Psilocybin-Induced Subjective Experiences in Humans. J. Psychopharmacol. 2021, 35, 384–397. [Google Scholar] [CrossRef]
- Holze, F.; Avedisian, I.; Varghese, N.; Eckert, A.; Liechti, M.E. Role of the 5-HT2A Receptor in Acute Effects of LSD on Empathy and Circulating Oxytocin. Front. Pharmacol. 2021, 12, 711255. [Google Scholar] [CrossRef]
- Nichols, D.E. Hallucinogens. Pharmacol. Ther. 2004, 101, 131–181. [Google Scholar] [CrossRef]
- Riba, J.; Valle, M.; Urbano, G.; Yritia, M.; Morte, A.; Barbanoj, M.J. Human Pharmacology of Ayahuasca: Subjective and Cardiovascular Effects, Monoamine Metabolite Excretion, and Pharmacokinetics. J. Pharmacol. Exp. Ther. 2003, 306, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Barker, S.A.; Monti, J.A.; Christian, S.T. Metabolism of the Hallucinogen N,N-Dimethyltryptamine in Rat Brain Homogenates. Biochem. Pharmacol. 1980, 29, 1049–1057. [Google Scholar] [CrossRef]
- Riba, J.; McIlhenny, E.H.; Bouso, J.C.; Barker, S.A. Metabolism and Urinary Disposition of N,N-Dimethyltryptamine after Oral and Smoked Administration: A Comparative Study. Drug Test. Anal. 2015, 7, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Abelaira, H.M.; Réus, G.Z.; Scaini, G.; Streck, E.L.; Crippa, J.A.; Quevedo, J. β-Carboline Harmine Reverses the Effects Induced by Stress on Behaviour and Citrate Synthase Activity in the Rat Prefrontal Cortex. Acta Neuropsychiatr. 2013, 25, 328–333. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.G.; Daros, G.C.; Santos, F.P.; Yonamine, M.; de Bitencourt, R.M. Antidepressant and Anxiolytic-like Effects of Ayahuasca in Rats Subjected to LPS-Induced Neuroinflammation. Behav. Brain Res. 2022, 434, 114007. [Google Scholar] [CrossRef]
- Daldegan-Bueno, D.; Favaro, V.M.; Morais, P.R.; Sussulini, A.; Oliveira, M.G.M. Effects of Repeated Ayahuasca Administration on Behaviour and C-Fos Expression in Male Rats Exposed to the Open Field. Behav. Brain Res. 2022, 427, 113878. [Google Scholar] [CrossRef]
- Farzin, D.; Mansouri, N. Antidepressant-like Effect of Harmane and Other β-Carbolines in the Mouse Forced Swim Test. Eur. Neuropsychopharmacol. 2006, 16, 324–328. [Google Scholar] [CrossRef]
- Fortunato, J.J.; Réus, G.Z.; Kirsch, T.R.; Stringari, R.B.; Stertz, L.; Kapczinski, F.; Pinto, J.P.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; et al. Acute Harmine Administration Induces Antidepressive-like Effects and Increases BDNF Levels in the Rat Hippocampus. Prog. Neuropsychopharmacol. Biol. Psychiatry 2009, 33, 1425–1430. [Google Scholar] [CrossRef]
- Fortunato, J.J.; Réus, G.Z.; Kirsch, T.R.; Stringari, R.B.; Fries, G.R.; Kapczinski, F.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Quevedo, J. Effects of β-Carboline Harmine on Behavioral and Physiological Parameters Observed in the Chronic Mild Stress Model: Further Evidence of Antidepressant Properties. Brain Res. Bull. 2010, 81, 491–496. [Google Scholar] [CrossRef]
- Fortunato, J.J.; Réus, G.Z.; Kirsch, T.R.; Stringari, R.B.; Fries, G.R.; Kapczinski, F.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Quevedo, J. Chronic Administration of Harmine Elicits Antidepressant-like Effects and Increases BDNF Levels in Rat Hippocampus. J. Neural Transm. 2010, 117, 1131–1137. [Google Scholar] [CrossRef]
- Hilber, P.; Chapillon, P. Effects of Harmaline on Anxiety-Related Behavior in Mice. Physiol. Behav. 2005, 86, 164–167. [Google Scholar] [CrossRef]
- Réus, G.Z.; Stringari, R.B.; de Souza, B.; Petronilho, F.; Dal-Pizzol, F.; Hallak, J.E.; Zuardi, A.W.; Crippa, J.A.; Quevedo, J. Harmine and Imipramine Promote Antioxidant Activities in Prefrontal Cortex and Hippocampus. Oxidative Med. Cell. Longev. 2010, 3, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Argento, E.; Capler, R.; Thomas, G.; Lucas, P.; Tupper, K.W. Exploring Ayahuasca-Assisted Therapy for Addiction: A Qualitative Analysis of Preliminary Findings among an Indigenous Community in Canada. Drug Alcohol Rev. 2019, 38, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Bouso, J.C.; González, D.; Fondevila, S.; Cutchet, M.; Fernández, X.; Ribeiro Barbosa, P.C.; Alcázar-Córcoles, M.; Araújo, W.S.; Barbanoj, M.J.; Fábregas, J.M.; et al. Personality, Psychopathology, Life Attitudes and Neuropsychological Performance among Ritual Users of Ayahuasca: A Longitudinal Study. PLoS ONE 2012, 7, e42421. [Google Scholar] [CrossRef] [PubMed]
- Bouso, J.C.; Palhano-Fontes, F.; Rodríguez-Fornells, A.; Ribeiro, S.; Sanches, R.; Crippa, J.A.S.; Hallak, J.E.C.; de Araujo, D.B.; Riba, J. Long-Term Use of Psychedelic Drugs Is Associated with Differences in Brain Structure and Personality in Humans. Eur. Neuropsychopharmacol. 2015, 25, 483–492. [Google Scholar] [CrossRef]
- Domínguez-Clavé, E.; Soler, J.; Pascual, J.C.; Elices, M.; Franquesa, A.; Valle, M.; Alvarez, E.; Riba, J. Ayahuasca Improves Emotion Dysregulation in a Community Sample and in Individuals with Borderline-like Traits. Psychopharmacology 2019, 236, 573–580. [Google Scholar] [CrossRef]
- Gonzalez, D.; Cantillo, J.; Perez, I.; Carvalho, M.; Aronovich, A.; Farre, M.; Feilding, A.; Obiols, J.E.; Bouso, J.C. The Shipibo Ceremonial Use of Ayahuasca to Promote Well-Being: An Observational Study. Front. Pharmacol. 2021, 12, 623923. [Google Scholar] [CrossRef]
- González, D.; Cantillo, J.; Pérez, I.; Farré, M.; Feilding, A.; Obiols, J.E.; Bouso, J.C. Therapeutic Potential of Ayahuasca in Grief: A Prospective, Observational Study. Psychopharmacology 2020, 237, 1171–1182. [Google Scholar] [CrossRef]
- Jiménez-Garrido, D.F.; Gómez-Sousa, M.; Ona, G.; Dos Santos, R.G.; Hallak, J.E.C.; Alcázar-Córcoles, M.; Bouso, J.C. Effects of Ayahuasca on Mental Health and Quality of Life in Naïve Users: A Longitudinal and Cross-Sectional Study Combination. Sci. Rep. 2020, 10, 4075. [Google Scholar] [CrossRef]
- Kohek, M.; Ona, G.; van Elk, M.; dos Santos, R.G.; Hallak, J.E.C.; Alcázar-Córcoles, M.; Bouso, J.C. Ayahuasca and Public Health II: Health Status in a Large Sample of Ayahuasca-Ceremony Participants in the Netherlands. J. Psychoact. Drugs 2022, 1–12. [Google Scholar] [CrossRef]
- Thomas, G.; Lucas, P.; Capler, N.R.; Tupper, K.W.; Martin, G. Ayahuasca-Assisted Therapy for Addiction: Results from a Preliminary Observational Study in Canada. Curr. Drug Abuse Rev. 2013, 6, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Uthaug, M.V.; van Oorsouw, K.; Kuypers, K.P.C.; van Boxtel, M.; Broers, N.J.; Mason, N.L.; Toennes, S.W.; Riba, J.; Ramaekers, J.G. Sub-Acute and Long-Term Effects of Ayahuasca on Affect and Cognitive Thinking Style and Their Association with Ego Dissolution. Psychopharmacology 2018, 235, 2979–2989. [Google Scholar] [CrossRef] [PubMed]
- van Oorsouw, K.; Toennes, S.W.; Ramaekers, J.G. Therapeutic Effect of an Ayahuasca Analogue in Clinically Depressed Patients: A Longitudinal Observational Study. Psychopharmacology 2022, 239, 1839–1852. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, R.G.; Landeira-Fernandez, J.; Strassman, R.J.; Motta, V.; Cruz, A.P. Effects of Ayahuasca on Psychometric Measures of Anxiety, Panic-like and Hopelessness in Santo Daime Members. J. Ethnopharmacol. 2007, 112, 507–513. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Rocha, J.M.; Rossi, G.N.; Osório, F.L.; Ona, G.; Bouso, J.C.; Silveira, G.O.; Yonamine, M.; Marchioni, C.; Crevelin, E.J.; et al. Effects of Ayahuasca on the Endocannabinoid System of Healthy Volunteers and in Volunteers with Social Anxiety Disorder: Results from Two Pilot, Proof-of-Concept, Randomized, Placebo-Controlled Trials. Hum. Psychopharmacol. 2022, 37, e2834. [Google Scholar] [CrossRef]
- Galvão-Coelho, N.L.; de Menezes Galvão, A.C.; de Almeida, R.N.; Palhano-Fontes, F.; Campos Braga, I.; Lobão Soares, B.; Maia-de-Oliveira, J.P.; Perkins, D.; Sarris, J.; de Araujo, D.B. Changes in Inflammatory Biomarkers Are Related to the Antidepressant Effects of Ayahuasca. J. Psychopharmacol. 2020, 34, 1125–1133. [Google Scholar] [CrossRef]
- Osório Fde, L.; Sanches, R.F.; Macedo, L.R.; Santos, R.G.; Maia-de-Oliveira, J.P.; Wichert-Ana, L.; Araujo, D.B.; Riba, J.; Crippa, J.A.; Hallak, J.E. Antidepressant Effects of a Single Dose of Ayahuasca in Patients with Recurrent Depression: A Preliminary Report. Braz. J. Psychiatry 2015, 37, 13–20. [Google Scholar] [CrossRef]
- Palhano-Fontes, F.; Barreto, D.; Onias, H.; Andrade, K.C.; Novaes, M.M.; Pessoa, J.A.; Mota-Rolim, S.A.; Osório, F.L.; Sanches, R.; Dos Santos, R.G.; et al. Rapid Antidepressant Effects of the Psychedelic Ayahuasca in Treatment-Resistant Depression: A Randomized Placebo-Controlled Trial. Psychol. Med. 2019, 49, 655–663. [Google Scholar] [CrossRef]
- Riba, J.; Romero, S.; Grasa, E.; Mena, E.; Carrió, I.; Barbanoj, M.J. Increased Frontal and Paralimbic Activation Following Ayahuasca, the Pan-Amazonian Inebriant. Psychopharmacology 2006, 186, 93–98. [Google Scholar] [CrossRef]
- Sanches, R.F.; Osório, F.L.; dos Santos, R.G.; Macedo, L.R.H.; Maia-de-Oliveira, J.P.; Wichert-Ana, L.; de Araujo, D.B.; Riba, J.; Crippa, J.A.S.; Hallak, J.E.C. Antidepressant Effects of a Single Dose of Ayahuasca in Patients with Recurrent Depression: A SPECT Study. J. Clin. Psychopharmacol. 2016, 36, 77–81. [Google Scholar] [CrossRef]
- Zeifman, R.J.; Singhal, N.; Dos Santos, R.G.; Sanches, R.F.; de Lima Osório, F.; Hallak, J.E.C.; Weissman, C.R. Rapid and Sustained Decreases in Suicidality Following a Single Dose of Ayahuasca among Individuals with Recurrent Major Depressive Disorder: Results from an Open-Label Trial. Psychopharmacology 2021, 238, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Aaghaz, S.; Sharma, K.; Jain, R.; Kamal, A. β-Carbolines as Potential Anticancer Agents. Eur. J. Med. Chem. 2021, 216, 113321. [Google Scholar] [CrossRef]
- dos Santos, R.G.; Osório, F.L.; Crippa, J.A.; Riba, J.; Zuardi, A.W.; Hallak, J.E. Antidepressive, Anxiolytic, and Antiaddictive Effects of Ayahuasca, Psilocybin and Lysergic Acid Diethylamide (LSD): A Systematic Review of Clinical Trials Published in the Last 25 Years. Ther. Adv. Psychopharmacol. 2016, 6, 193–213. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, R.G.; Hallak, J.E.C. Effects of the Natural β-Carboline Alkaloid Harmine, a Main Constituent of Ayahuasca, in Memory and in the Hippocampus: A Systematic Literature Review of Preclinical Studies. J. Psychoact. Drugs 2017, 49, 1–10. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, R.G.; Hallak, J.E.C. Ayahuasca, an Ancient Substance with Traditional and Contemporary Use in Neuropsychiatry and Neuroscience. Epilepsy Behav. 2021, 121, 106300. [Google Scholar] [CrossRef]
- Galvão-Coelho, N.L.; Marx, W.; Gonzalez, M.; Sinclair, J.; de Manincor, M.; Perkins, D.; Sarris, J. Classic Serotonergic Psychedelics for Mood and Depressive Symptoms: A Meta-Analysis of Mood Disorder Patients and Healthy Participants. Psychopharmacology 2021, 238, 341–354. [Google Scholar] [CrossRef]
- Hamill, J.; Hallak, J.; Dursun, S.M.; Baker, G. Ayahuasca: Psychological and Physiologic Effects, Pharmacology and Potential Uses in Addiction and Mental Illness. Curr. Neuropharmacol. 2019, 17, 108–128. [Google Scholar] [CrossRef]
- Nunes, A.A.; Dos Santos, R.G.; Osório, F.L.; Sanches, R.F.; Crippa, J.A.; Hallak, J.E. Effects of Ayahuasca and Its Alkaloids on Drug Dependence: A Systematic Literature Review of Quantitative Studies in Animals and Humans. J. Psychoact. Drugs 2016, 48, 195–205. [Google Scholar] [CrossRef]
- Rodrigues, L.S.; Rossi, G.N.; Rocha, J.M.; Osório, F.L.; Bouso, J.C.; Hallak, J.E.C.; Dos Santos, R.G. Effects of Ayahuasca and Its Alkaloids on Substance Use Disorders: An Updated (2016–2020) Systematic Review of Preclinical and Human Studies. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 272, 541–556. [Google Scholar] [CrossRef]
- Calder, A.E.; Hasler, G. Towards an Understanding of Psychedelic-Induced Neuroplasticity. Neuropsychopharmacology 2022, 1–9. [Google Scholar] [CrossRef]
- Wolf, M.E.; Abi-Dargham, A. Synaptic Plasticity as a Therapeutic Target to Modulate Circuits in Psychiatric Disorders. Neuropsychopharmacology 2022, 1–2. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, D.C.; Syed, S.A.; Flynn, L.T.; Safi-Aghdam, H.; Cozzi, N.V.; Ranganathan, M. Exploratory Study of the Dose-Related Safety, Tolerability, and Efficacy of Dimethyltryptamine (DMT) in Healthy Volunteers and Major Depressive Disorder. Neuropsychopharmacology 2022, 47, 1854–1862. [Google Scholar] [CrossRef] [PubMed]
- Gasser, P.; Holstein, D.; Michel, Y.; Doblin, R.; Yazar-Klosinski, B.; Passie, T.; Brenneisen, R. Safety and Efficacy of Lysergic Acid Diethylamide-Assisted Psychotherapy for Anxiety Associated With Life-Threatening Diseases. J. Nerv. Ment. Dis. 2014, 202, 513–520. [Google Scholar] [CrossRef]
- Jelen, L.A.; Stone, J.M. Ketamine for Depression. Int. Rev. Psychiatry 2021, 33, 207–228. [Google Scholar] [CrossRef] [PubMed]
- Delille, H.K.; Becker, J.M.; Burkhardt, S.; Bleher, B.; Terstappen, G.C.; Schmidt, M.; Meyer, A.H.; Unger, L.; Marek, G.J.; Mezler, M. Heterocomplex Formation of 5-HT2A-MGlu2 and Its Relevance for Cellular Signaling Cascades. Neuropsychopharmacology 2012, 62, 2184–2191. [Google Scholar] [CrossRef]
- Moreno, J.L.; Holloway, T.; Albizu, L.; Sealfon, S.C.; González-Maeso, J. Metabotropic Glutamate MGlu2 Receptor Is Necessary for the Pharmacological and Behavioral Effects Induced by Hallucinogenic 5-HT2A Receptor Agonists. Neurosci. Lett. 2011, 493, 76–79. [Google Scholar] [CrossRef]
- Brierley, D.I.; Davidson, C. Harmine Augments Electrically Evoked Dopamine Efflux in the Nucleus Accumbens Shell. J. Psychopharmacol. 2013, 27, 98–108. [Google Scholar] [CrossRef]
- Iurlo, M.; Leone, G.; Schilström, B.; Linnér, L.; Nomikos, G.; Hertel, P.; Silvestrini, B.; Svensson, H. Effects of Harmine on Dopamine Output and Metabolism in Rat Striatum: Role of Monoamine Oxidase-A Inhibition. Psychopharmacology 2001, 159, 98–104. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Crippa, J.A.; Osório, F.L.; Rocha, J.M.; Rossi, G.N.; Marchioni, C.; Queiroz, M.E.C.; Silveira, G.O.; Yonamine, M.; Hallak, J.E.C. Possible Interactions Between 5-HT2A Receptors and the Endocannabinoid System in Humans: Preliminary Evidence of Interactive Effects of Ayahuasca and Endocannabinoids in a Healthy Human Subject. J. Clin. Psychopharmacol. 2018, 38, 644–646. [Google Scholar] [CrossRef]
- Cozzi, N.V.; Gopalakrishnan, A.; Anderson, L.L.; Feih, J.T.; Shulgin, A.T.; Daley, P.F.; Ruoho, A.E. Dimethyltryptamine and Other Hallucinogenic Tryptamines Exhibit Substrate Behavior at the Serotonin Uptake Transporter and the Vesicle Monoamine Transporter. J. Neural Transm. 2009, 116, 1591–1599. [Google Scholar] [CrossRef]
- Nagai, F.; Nonaka, R.; Satoh Hisashi Kamimura, K. The Effects of Non-Medically Used Psychoactive Drugs on Monoamine Neurotransmission in Rat Brain. Eur. J. Pharmacol. 2007, 559, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Bunzow, J.R.; Sonders, M.S.; Arttamangkul, S.; Harrison, L.M.; Zhang, G.; Quigley, D.I.; Darland, T.; Suchland, K.L.; Pasumamula, S.; Kennedy, J.L.; et al. Amphetamine, 3,4-Methylenedioxymethamphetamine, Lysergic Acid Diethylamide, and Metabolites of the Catecholamine Neurotransmitters Are Agonists of a Rat Trace Amine Receptor. Mol. Pharmacol. 2001, 60, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Fontanilla, D.; Johannessen, M.; Hajipour, A.R.; Cozzi, N.V.; Jackson, M.B.; Ruoho, A.E. The Hallucinogen N,N -Dimethyltryptamine (DMT) Is an Endogenous Sigma-1 Receptor Regulator. Science 2009, 323, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Su, T.P.; Hayashi, T.; Vaupel, D.B. When the Endogenous Hallucinogenic Trace Amine N,N-Dimethyltryptamine Meets the Sigma-1 Receptor. Sci. Signal. 2009, 2, pe12. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, S.; Netzband, N.; Bird, C.; Young, A.H.; Juruena, M.F. The Pharmacological Interaction of Compounds in Ayahuasca: A Systematic Review. Braz. J. Psychiatry 2020, 42, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Grob, C.S.; McKenna, D.J.; Callaway, J.C.; Brito, G.S.; Neves, E.S.; Oberlaender, G.; Saide, O.L.; Labigalini, E.; Tacla, C.; Miranda, C.T.; et al. Human Psychopharmacology of Hoasca, a Plant Hallucinogen Used in Ritual Context in Brazil. J. Nerv. Ment. Dis. 1996, 184, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, D.X.; Grob, C.S.; de Rios, M.D.; Lopez, E.; Alonso, L.K.; Tacla, C.; Doering-Silveira, E. Ayahuasca in Adolescence: A Preliminary Psychiatric Assessment. J. Psychoact. Drugs 2005, 37, 129–133. [Google Scholar] [CrossRef]
- Sarris, J.; Perkins, D.; Cribb, L.; Schubert, V.; Opaleye, E.; Bouso, J.C.; Scheidegger, M.; Aicher, H.; Simonova, H.; Horák, M.; et al. Ayahuasca Use and Reported Effects on Depression and Anxiety Symptoms: An International Cross-Sectional Study of 11,912 Consumers. J. Affect. Disord. Rep. 2021, 4, 100098. [Google Scholar] [CrossRef]
- Galvão, A.C.M.; de Almeida, R.N.; Silva, E.; Freire, F.A.M.; Palhano-Fontes, F.; Onias, H.; Arcoverde, E.; Maia-de-Oliveira, J.P.; de Araújo, D.B.; Lobão-Soares, B.; et al. Cortisol Modulation by Ayahuasca in Patients With Treatment Resistant Depression and Healthy Controls. Front. Psychiatry 2018, 9, 185. [Google Scholar] [CrossRef]
- Zeifman, R.J.; Palhano-Fontes, F.; Hallak, J.; Arcoverde, E.; Maia-Oliveira, J.P.; Araujo, D.B. The Impact of Ayahuasca on Suicidality: Results From a Randomized Controlled Trial. Front. Pharmacol. 2019, 10, 1325. [Google Scholar] [CrossRef]
- de Almeida, R.N.; Galvão, A.C.M.; da Silva, F.S.; Silva, E.; Palhano-Fontes, F.; Maia-de-Oliveira, J.P.; de Araújo, L.B.; Lobão-Soares, B.; Galvão-Coelho, N.L. Modulation of Serum Brain-Derived Neurotrophic Factor by a Single Dose of Ayahuasca: Observation From a Randomized Controlled Trial. Front. Psychol. 2019, 10, 1234. [Google Scholar] [CrossRef] [PubMed]
- Bauer, I.L. Ayahuasca: A risk for travellers? Travel Med. Infect. Dis. 2018, 21, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Callaway, J.C.; Grob, C.S. Ayahuasca Preparations and Serotonin Reuptake Inhibitors: A Potential Combination for Severe Adverse Interactions. J. Psychoact. Drugs 1998, 30, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.N.; Dias, I.C.D.S.; Baker, G.; Bouso Saiz, J.C.; Dursun, S.M.; Hallak, J.E.C.; Dos Santos, R.G. Ayahuasca, a Potentially Rapid Acting Antidepressant: Focus on Safety and Tolerability. Expert Opin. Drug Saf. 2022, 21, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.N.; Hallak, J.E.C.; Baker, G.; Dursun, S.M.; Dos Santos, R.G. The Effects of Ketamine and Classic Hallucinogens on Neurotrophic and Inflammatory Markers in Unipolar Treatment-Resistant Depression: A Systematic Review of Clinical Trials. Eur. Arch. Psychiatry Clin. Neurosci. 2022, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.M.; Rossi, G.N.; Osório, F.L.; Hallak, J.E.C.; Dos Santos, R.G. Adverse Effects After Ayahuasca Administration in the Clinical Setting. J. Clin. Psychopharmacol. 2022, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.J. Clinical Investigations of the Therapeutic Potential of Ayahuasca: Rationale and Regulatory Challenges. Pharmacol. Ther. 2004, 102, 111–129. [Google Scholar] [CrossRef]
- Callaway, J.C. Various Alkaloid Profiles in Decoctions of Banisteriopsis caapi. J. Psychoact. Drugs 2005, 37, 151–155. [Google Scholar] [CrossRef]
- Santos, M.C.; Navickiene, S.; Gaujac, A. Determination of Tryptamines and β-Carbolines in Ayahuasca Beverage Consumed During Brazilian Religious Ceremonies. J. AOAC Int. 2017, 100, 820–824. [Google Scholar] [CrossRef][Green Version]
- Souza, R.C.Z.; Zandonadi, F.S.; Freitas, D.P.; Tófoli, L.F.F.; Sussulini, A. Validation of an Analytical Method for the Determination of the Main Ayahuasca Active Compounds and Application to Real Ayahuasca Samples from Brazil. J. Chromatogr. B 2019, 1124, 197–203. [Google Scholar] [CrossRef]
- Santos, B.W.L.; de Oliveira, R.C.; Sonsin-Oliveira, J.; Fagg, C.W.; Barbosa, J.B.F.; Caldas, E.D. Biodiversity of β-Carboline Profile of Banisteriopsis Caapi and Ayahuasca, a Plant and a Brew with Neuropharmacological Potential. Plants 2020, 9, 870. [Google Scholar] [CrossRef] [PubMed]
- Kaasik, H.; Souza, R.C.Z.; Zandonadi, F.S.; Tófoli, L.F.; Sussulini, A. Chemical Composition of Traditional and Analog Ayahuasca. J. Psychoact. Drugs 2021, 53, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Riba, J.; Rodríguez-Fornells, A.; Urbano, G.; Morte, A.; Antonijoan, R.; Montero, M.; Callaway, J.C.; Barbanoj, M.J. Subjective Effects and Tolerability of the South American Psychoactive Beverage Ayahuasca in Healthy Volunteers. Psychopharmacology 2001, 154, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, R.G.; Grasa, E.; Valle, M.; Ballester, M.R.; Bouso, J.C.; Nomdedéu, J.F.; Homs, R.; Barbanoj, M.J.; Riba, J. Pharmacology of Ayahuasca Administered in Two Repeated Doses. Psychopharmacology 2012, 219, 1039–1053. [Google Scholar] [CrossRef]
- Rocha, J.M.; Rossi, G.N.; de Lima Osório, F.; Bouso, J.C.; de Oliveira Silveira, G.; Yonamine, M.; Campos, A.C.; Bertozi, G.; Cecílio Hallak, J.E.; Dos Santos, R.G. Effects of Ayahuasca on the Recognition of Facial Expressions of Emotions in Naive Healthy Volunteers: A Pilot, Proof-of-Concept, Randomized Controlled Trial. J. Clin. Psychopharmacol. 2021, 41, 267–274. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Osório, F.D.L.; Rocha, J.M.; Rossi, G.N.; Bouso, J.C.; Rodrigues, L.S.; Silveira, G.D.O.; Yonamine, M.; Hallak, J.E.C. Ayahuasca improves self-perception of speech performance in subjects with social anxiety disorder. J. Clin. Psychopharmacol. 2021, 41, 540–550. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Bouso, J.C.; Rocha, J.M.; Rossi, G.N.; Hallak, J.E. The Use of Classic Hallucinogens/Psychedelics in a Therapeutic Context: Healthcare Policy Opportunities and Challenges. Risk Manag. Healthc. Policy 2021, 14, 901–910. [Google Scholar] [CrossRef]
- Callaway, J.C.; McKenna, D.J.; Grob, C.S.; Brito, G.S.; Raymon, L.P.; Poland, R.E.; Andrade, E.N.; Andrade, E.O.; Mash, D.C. Pharmacokinetics of Hoasca Alkaloids in Healthy Humans. J. Ethnopharmacol. 1999, 65, 243–256. [Google Scholar] [CrossRef]
- Frecska, E.; Szabo, A.; Winkelman, M.J.; Luna, L.E.; McKenna, D.J. A Possibly Sigma-1 Receptor Mediated Role of Dimethyltryptamine in Tissue Protection, Regeneration, and Immunity. J. Neural Transm. 2013, 120, 1295–1303. [Google Scholar] [CrossRef]
- Cohen, I.; Vogel, W.H. Determination and Physiological Disposition of Dimethyltryptamine and Diethyltryptamine in Rat Brain, Liver and Plasma. Biochem. Pharmacol. 1972, 21, 1214–1216. [Google Scholar] [CrossRef]
- Barker, S.A.; Beaton, J.M.; Christian, S.T.; Monti, J.A.; Morris, P.E. Comparison of the Brain Levels of N,N-Dimethyltryptamine and Alpha, Alpha, Beta, Beta-Tetradeutero-N-N-Dimethyltryptamine Following Intraperitoneal Injection. The in Vivo Kinetic Isotope Effect. Biochem. Pharmacol. 1982, 31, 2513–2516. [Google Scholar] [CrossRef]
- Christian, S.T.; Harrison, R.; Quayle, E.; Pagel, J.; Monti, J. The in Vitro Identification of Dimethyltryptamine (DMT) in Mammalian Brain and Its Characterization as a Possible Endogenous Neuroregulatory Agent. Biochem. Med. 1977, 18, 164–183. [Google Scholar] [CrossRef]
- Busch, A.E.; Karbach, U.; Miska, D.; Gorboulev, V.; Akhoundova, A.; Volk, C.; Arndt, P.; Ulzheimer, J.C.; Sonders, M.S.; Baumann, C.; et al. Human Neurons Express the Polyspecific Cation Transporter HOCT2, Which Translocates Monoamine Neurotransmitters, Amantadine, and Memantine. Mol. Pharmacol. 1998, 54, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, B.R.; Lockett, L.; Talomsin, R.; Blackman, G.L.; McLeod, W.R. In Vivo Metabolism of 5-Methoxy-N,N-Dimethyltryptamine and N,N-Dimethyltryptamine in the Rat. Biochem. Pharmacol. 1987, 36, 1509–1512. [Google Scholar] [CrossRef]
- Takahashi, T.; Takahashi, K.; Ido, T.; Yanai, K.; Iwata, R.; Ishiwata, K.; Nozoe, S. 11C-Labeling of Indolealkylamine Alkaloids and the Comparative Study of Their Tissue Distributions. Int. J. Appl. Radiat. Isot. 1985, 36, 965–969. [Google Scholar] [CrossRef]
- Yanai, K.; Ido, T.; Ishiwata, K.; Hatazawa, J.; Takahashi, T.; Iwata, R.; Matsuzawa, T. In Vivo Kinetics and Displacement Study of a Carbon-11-Labeled Hallucinogen, N,N-[11C]Dimethyltryptamine. Eur. J. Nucl. Med. 1986, 12, 141–146. [Google Scholar] [CrossRef]
- Vitale, A.A.; Pomilio, A.B.; Cañellas, C.O.; Vitale, M.G.; Putz, E.M.; Ciprian-Ollivier, J. In Vivo Long-Term Kinetics of Radiolabeled n,n-Dimethyltryptamine and Tryptamine. J. Nucl. Med. 2011, 52, 970–977. [Google Scholar] [CrossRef]
- Blough, B.E.; Landavazo, A.; Decker, A.M.; Partilla, J.S.; Baumann, M.H.; Rothman, R.B. Interaction of Psychoactive Tryptamines with Biogenic Amine Transporters and Serotonin Receptor Subtypes. Psychopharmacology 2014, 231, 4135–4144. [Google Scholar] [CrossRef]
- Glennon, R.A.; Dukat, M.; Grella, B.; Hong, S.-S.; Costantino, L.; Teitler, M.; Smith, C.; Egan, C.; Davis, K.; Mattson, M.V. Binding of β-Carbolines and Related Agents at Serotonin (5-HT2 and 5-HT1A), Dopamine (D2) and Benzodiazepine Receptors. Drug Alcohol Depend. 2000, 60, 121–132. [Google Scholar] [CrossRef]
- Jiménez, J.H.; Bouso, J.C. Significance of Mammalian N, N-Dimethyltryptamine (DMT): A 60-Year-Old Debate. J. Psychopharmacol. 2022, 36, 905–919. [Google Scholar] [CrossRef]
- Keiser, M.J.; Setola, V.; Irwin, J.J.; Laggner, C.; Abbas, A.I.; Hufeisen, S.J.; Jensen, N.H.; Kuijer, M.B.; Matos, R.C.; Tran, T.B.; et al. Predicting New Molecular Targets for Known Drugs. Nature 2009, 462, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Strassman, R.J. Human Psychopharmacology of N,N-Dimethyltryptamine. Behav. Brain Res. 1995, 73, 121–124. [Google Scholar] [CrossRef]
- Ray, T.S. Psychedelics and the Human Receptorome. PLoS ONE 2010, 5, e9019. [Google Scholar] [CrossRef]
- Carbonaro, T.M.; Gatch, M.B. Neuropharmacology of N,N-Dimethyltryptamine. Brain Res. Bull. 2016, 126, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Sub Laban, T.; Saadabadi, A. Monoamine Oxidase Inhibitors (MAOI); StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Abu Ghazaleh, H.; Lalies, M.D.; Nutt, D.J.; Hudson, A.L. The Modulatory Action of Harmane on Serotonergic Neurotransmission in Rat Brain. Brain Res. 2015, 1597, 57–64. [Google Scholar] [CrossRef]
- Smith, K.L.; Ford, G.K.; Jessop, D.S.; Finn, D.P. Behavioural, Neurochemical and Neuroendocrine Effects of the Endogenous β-Carboline Harmane in Fear-Conditioned Rats. J. Psychopharmacol. 2013, 27, 162–170. [Google Scholar] [CrossRef]
- Morales-García, J.A.; de la Fuente Revenga, M.; Alonso-Gil, S.; Rodríguez-Franco, M.I.; Feilding, A.; Perez-Castillo, A.; Riba, J. The Alkaloids of Banisteriopsis Caapi, the Plant Source of the Amazonian Hallucinogen Ayahuasca, Stimulate Adult Neurogenesis in Vitro. Sci. Rep. 2017, 7, 5309. [Google Scholar] [CrossRef]
- Valle, M.; Maqueda, A.E.; Rabella, M.; Rodríguez-Pujadas, A.; Antonijoan, R.M.; Romero, S.; Alonso, J.F.; Mañanas, M.À.; Barker, S.; Friedlander, P.; et al. Inhibition of Alpha Oscillations through Serotonin-2A Receptor Activation Underlies the Visual Effects of Ayahuasca in Humans. Eur. Neuropsychopharmacol. 2016, 26, 1161–1175. [Google Scholar] [CrossRef]
- Preller, K.H.; Burt, J.B.; Ji, J.L.; Schleifer, C.H.; Adkinson, B.D.; Stämpfli, P.; Seifritz, E.; Repovs, G.; Krystal, J.H.; Murray, J.D.; et al. Changes in Global and Thalamic Brain Connectivity in LSD-Induced Altered States of Consciousness Are Attributable to the 5-HT2A Receptor. Elife 2018, 7, e35082. [Google Scholar] [CrossRef]
- Olbrich, S.; Preller, K.H.; Vollenweider, F.X. LSD and Ketanserin and Their Impact on the Human Autonomic Nervous System. Psychophysiology 2021, 58, e13822. [Google Scholar] [CrossRef]
- Quednow, B.B.; Kometer, M.; Geyer, M.A.; Vollenweider, F.X. Psilocybin-Induced Deficits in Automatic and Controlled Inhibition Are Attenuated by Ketanserin in Healthy Human Volunteers. Neuropsychopharmacology 2012, 37, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Kometer, M.; Schmidt, A.; Jäncke, L.; Vollenweider, F.X. Activation of Serotonin 2A Receptors Underlies the Psilocybin-Induced Effects on α Oscillations, N170 Visual-Evoked Potentials, and Visual Hallucinations. J. Neurosci. 2013, 33, 10544–10551. [Google Scholar] [CrossRef] [PubMed]
- Chagraoui, A.; Thibaut, F.; Skiba, M.; Thuillez, C.; Bourin, M. 5-HT2C Receptors in Psychiatric Disorders: A Review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 66, 120–135. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.; DeLorenzo, C.; Choudhury, S.; Parsey, R.V. The 5-HT1A Receptor in Major Depressive Disorder. Eur. Neuropsychopharmacol. 2016, 26, 397–410. [Google Scholar] [CrossRef]
- Garcia-Garcia, A.L.; Newman-Tancredi, A.; Leonardo, E.D. 5-HT(1A) [Corrected] Receptors in Mood and Anxiety: Recent Insights into Autoreceptor versus Heteroreceptor Function. Psychopharmacology 2014, 231, 623–636. [Google Scholar] [CrossRef]
- Savitz, J.; Lucki, I.; Drevets, W.C. 5-HT(1A) Receptor Function in Major Depressive Disorder. Prog. Neurobiol. 2009, 88, 17–31. [Google Scholar] [CrossRef]
- Kreiss, D.S.; Lucki, I. Effects of Acute and Repeated Administration of Antidepressant Drugs on Extracellular Levels of 5-Hydroxytryptamine Measured in Vivo. J. Pharmacol. Exp. Ther. 1995, 274, 866–876. [Google Scholar]
- Rausch, J.L.; Johnson, M.E.; Kasik, K.E.; Stahl, S.M. Temperature Regulation in Depression: Functional 5HT1A Receptor Adaptation Differentiates Antidepressant Response. Neuropsychopharmacology 2006, 31, 2274–2280. [Google Scholar] [CrossRef][Green Version]
- Giovacchini, G.; Lang, L.; Ma, Y.; Herscovitch, P.; Eckelman, W.C.; Carson, R.E. Differential Effects of Paroxetine on Raphe and Cortical 5-HT1A Binding: A PET Study in Monkeys. Neuroimage 2005, 28, 238–248. [Google Scholar] [CrossRef]
- Fricker, A.D.; Rios, C.; Devi, L.A.; Gomes, I. Serotonin Receptor Activation Leads to Neurite Outgrowth and Neuronal Survival. Brain Res. Mol. Brain Res. 2005, 138, 228–235. [Google Scholar] [CrossRef]
- Samuels, B.A.; Anacker, C.; Hu, A.; Levinstein, M.R.; Pickenhagen, A.; Tsetsenis, T.; Madroñal, N.; Donaldson, Z.R.; Drew, L.J.; Dranovsky, A.; et al. 5-HT1A Receptors on Mature Dentate Gyrus Granule Cells Are Critical for the Antidepressant Response. Nat. Neurosci. 2015, 18, 1606–1616. [Google Scholar] [CrossRef] [PubMed]
- Claustre, Y.; Benavides, J.; Scatton, B. Potential Mechanisms Involved in the Negative Coupling between Serotonin 5-HT1A Receptors and Carbachol-Stimulated Phosphoinositide Turnover in the Rat Hippocampus. J. Neurochem. 1991, 56, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.R.; Mukhin, Y.V.; Gelasco, A.; Turner, J.; Collinsworth, G.; Gettys, T.W.; Grewal, J.S.; Garnovskaya, M.N. Multiplicity of Mechanisms of Serotonin Receptor Signal Transduction. Pharmacol. Ther. 2001, 92, 179–212. [Google Scholar] [CrossRef]
- Deliganis, A.V.; Pierce, P.A.; Peroutka, S.J. Differential Interactions of Dimethyltryptamine (DMT) with 5-HT1A and 5-HT2 Receptors. Biochem. Pharmacol. 1991, 41, 1739–1744. [Google Scholar] [CrossRef]
- Chenu, F.; Mansari, M.E.; Blier, P. Long-Term Administration of Monoamine Oxidase Inhibitors Alters the Firing Rate and Pattern of Dopamine Neurons in the Ventral Tegmental Area. Int. J. Neuropsychopharmacol. 2009, 12, 475–485. [Google Scholar] [CrossRef]
- Jiang, X.L.; Shen, H.W.; Yu, A.M. Potentiation of 5-Methoxy-N,N-Dimethyltryptamine-Induced Hyperthermia by Harmaline and the Involvement of Activation of 5-HT1A and 5-HT2A Receptors. Neuropharmacology 2015, 89, 342–351. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Nutt, D.J. Serotonin and Brain Function: A Tale of Two Receptors. J. Psychopharmacol. 2017, 31, 1091–1120. [Google Scholar] [CrossRef]
- Stamper, C.E.; Hassell, J.E.; Kapitz, A.J.; Renner, K.J.; Orchinik, M.; Lowry, C.A. Activation of 5-HT(1A) Receptors in the Rat Dorsomedial Hypothalamus Inhibits Stress-Induced Activation of the Hypothalamic-Pituitary-Adrenal Axis. Stress 2017, 20, 223–230. [Google Scholar] [CrossRef]
- Zhou, J.; Cao, X.; Mar, A.C.; Ding, Y.Q.; Wang, X.; Li, Q.; Li, L. Activation of Postsynaptic 5-HT1A Receptors Improve Stress Adaptation. Psychopharmacology 2014, 231, 2067–2075. [Google Scholar] [CrossRef]
- Tafet, G.E.; Nemeroff, C.B. Pharmacological Treatment of Anxiety Disorders: The Role of the HPA Axis. Front. Psychiatry 2020, 11, 443. [Google Scholar] [CrossRef]
- Jaster, A.M.; de la Fuente Revenga, M.; González-Maeso, J. Molecular Targets of Psychedelic-induced Plasticity. J. Neurochem. 2022, 162, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Rojas, P.S.; Neira, D.; Muñoz, M.; Lavandero, S.; Fiedler, J.L. Serotonin (5-HT) Regulates Neurite Outgrowth through 5-HT1A and 5-HT7 Receptors in Cultured Hippocampal Neurons. J. Neurosci. Res. 2014, 92, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Cornea-Hébert, V.; Riad, M.; Wu, C.; Singh, S.K.; Descarries, L. Cellular and Subcellular Distribution of the Serotonin 5-HT2A Receptor in the Central Nervous System of Adult Rat. J. Comp. Neurol. 1999, 409, 187–209. [Google Scholar] [CrossRef]
- Zhang, G.; Stackman, R.W. The Role of Serotonin 5-HT2A Receptors in Memory and Cognition. Front. Pharmacol. 2015, 6, 225. [Google Scholar] [CrossRef] [PubMed]
- Masson, J.; Emerit, M.B.; Hamon, M.; Darmon, M. Serotonergic Signaling: Multiple Effectors and Pleiotropic Effects. Wiley Interdiscip. Rev.: Membr. Transp. Signal. 2012, 1, 685–713. [Google Scholar] [CrossRef]
- Schmid, C.L.; Bohn, L.M. Serotonin, But Not N-Methyltryptamines, Activates the Serotonin 2A Receptor via a β-Arrestin2/Src/Akt Signaling Complex In Vivo. J. Neurosci. 2010, 30, 13513–13524. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.A.; Vaidya, V.A. Differential Signaling Signatures Evoked by DOI versus Lisuride Stimulation of the 5-HT2A Receptor. Biochem. Biophys. Res. Commun. 2020, 531, 609–614. [Google Scholar] [CrossRef]
- Eshleman, A.J.; Forster, M.J.; Wolfrum, K.M.; Johnson, R.A.; Janowsky, A.; Gatch, M.B. Behavioral and Neurochemical Pharmacology of Six Psychoactive Substituted Phenethylamines: Mouse Locomotion, Rat Drug Discrimination and in Vitro Receptor and Transporter Binding and Function. Psychopharmacology 2014, 231, 875–888. [Google Scholar] [CrossRef]
- Grella, B.; Dukat, M.; Young, R.; Teitler, M.; Herrick-Davis, K.; Gauthier, C.B.; Glennon, R.A. Investigation of Hallucinogenic and Related Beta-Carbolines. Drug Alcohol Depend. 1998, 50, 99–107. [Google Scholar] [CrossRef]
- Grella, B.; Teitler, M.; Smith, C.; Herrick-Davis, K.; Glennon, R.A. Binding of β-Carbolines at 5-HT2 Serotonin Receptors. Bioorganic Med. Chem. Lett. 2003, 13, 4421–4425. [Google Scholar] [CrossRef]
- Colaço, C.S.; Alves, S.S.; Nolli, L.M.; Pinheiro, W.O.; de Oliveira, D.G.R.; Santos, B.W.L.; Pic-Taylor, A.; Mortari, M.R.; Caldas, E.D. Toxicity of Ayahuasca after 28 Days Daily Exposure and Effects on Monoamines and Brain-Derived Neurotrophic Factor (BDNF) in Brain of Wistar Rats. Metab. Brain Dis. 2020, 35, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Raval, N.R.; Johansen, A.; Donovan, L.L.; Ros, N.F.; Ozenne, B.; Hansen, H.D.; Knudsen, G.M. A Single Dose of Psilocybin Increases Synaptic Density and Decreases 5-HT(2A) Receptor Density in the Pig Brain. Int. J. Mol. Sci. 2021, 22, 835. [Google Scholar] [CrossRef] [PubMed]
- Farber, N.B.; Hanslick, J.; Kirby, C.; McWilliams, L.; Olney, J.W. Serotonergic Agents That Activate 5HT2A Receptors Prevent NMDA Antagonist Neurotoxicity. Neuropsychopharmacology 1998, 18, 57–62. [Google Scholar] [CrossRef]
- Nutt, D. 5HT2a Receptors—A New Target for Depression? Eur. Psychiatry 2015, 30, 35. [Google Scholar] [CrossRef]
- Palhano-Fontes, F.; Andrade, K.C.; Tofoli, L.F.; Santos, A.C.; Crippa, J.A.; Hallak, J.E.; Ribeiro, S.; de Araujo, D.B. The Psychedelic State Induced by Ayahuasca Modulates the Activity and Connectivity of the Default Mode Network. PLoS ONE 2015, 10, e0118143. [Google Scholar] [CrossRef] [PubMed]
- Downey, D.; Dutta, A.; McKie, S.; Dawson, G.R.; Dourish, C.T.; Craig, K.; Smith, M.A.; McCarthy, D.J.; Harmer, C.J.; Goodwin, G.M.; et al. Comparing the Actions of Lanicemine and Ketamine in Depression: Key Role of the Anterior Cingulate. Eur. Neuropsychopharmacol. 2016, 26, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Hesselgrave, N.; Troppoli, T.A.; Wulff, A.B.; Cole, A.B.; Thompson, S.M. Harnessing Psilocybin: Antidepressant-like Behavioral and Synaptic Actions of Psilocybin Are Independent of 5-HT2R Activation in Mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2022489118. [Google Scholar] [CrossRef]
- Vollenweider, F.X.; Vollenweider-Scherpenhuyzen, M.F.; Bäbler, A.; Vogel, H.; Hell, D. Psilocybin Induces Schizophrenia-like Psychosis in Humans via a Serotonin-2 Agonist Action. Neuroreport 1998, 9, 3897–3902. [Google Scholar] [CrossRef]
- Carhart-Harris, R.; Giribaldi, B.; Watts, R.; Baker-Jones, M.; Murphy-Beiner, A.; Murphy, R.; Martell, J.; Blemings, A.; Erritzoe, D.; Nutt, D.J. Trial of Psilocybin versus Escitalopram for Depression. N. Engl. J. Med. 2021, 384, 1402–1411. [Google Scholar] [CrossRef]
- Davis, A.K.; Barrett, F.S.; May, D.G.; Cosimano, M.P.; Sepeda, N.D.; Johnson, M.W.; Finan, P.H.; Griffiths, R.R. Effects of Psilocybin-Assisted Therapy on Major Depressive Disorder. JAMA Psychiatry 2021, 78, 481–489. [Google Scholar] [CrossRef]
- McGrew, L.; Chang, M.S.; Sanders-Bush, E. Phospholipase D Activation by Endogenous 5-Hydroxytryptamine 2C Receptors Is Mediated by Galpha13 and Pertussis Toxin-Insensitive Gbetagamma Subunits. Mol. Pharmacol. 2002, 62, 1339–1343. [Google Scholar] [CrossRef]
- Berg, K.A.; Maayani, S.; Goldfarb, J.; Scaramellini, C.; Leff, P.; Clarke, W.P. Effector Pathway-Dependent Relative Efficacy at Serotonin Type 2A and 2C Receptors: Evidence for Agonist-Directed Trafficking of Receptor Stimulus. Mol. Pharmacol. 1998, 54, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Ślifirski, G.; Król, M.; Turło, J. 5-HT Receptors and the Development of New Antidepressants. Int. J. Mol. Sci. 2021, 22, 9015. [Google Scholar] [CrossRef] [PubMed]
- Dekeyne, A.; Mannoury la Cour, C.; Gobert, A.; Brocco, M.; Lejeune, F.; Serres, F.; Sharp, T.; Daszuta, A.; Soumier, A.; Papp, M.; et al. S32006, a Novel 5-HT2C Receptor Antagonist Displaying Broad-Based Antidepressant and Anxiolytic Properties in Rodent Models. Psychopharmacology 2008, 199, 549–568. [Google Scholar] [CrossRef] [PubMed]
- Dekeyne, A.; Brocco, M.; Loiseau, F.; Gobert, A.; Rivet, J.-M.; Di Cara, B.; Cremers, T.I.; Flik, G.; Fone, K.C.F.; Watson, D.J.G.; et al. S32212, a Novel Serotonin Type 2C Receptor Inverse Agonist/A2-Adrenoceptor Antagonist and Potential Antidepressant: II. A Behavioral, Neurochemical, and Electrophysiological Characterization. J. Pharmacol. Exp. Ther. 2012, 340, 765–780. [Google Scholar] [CrossRef]
- Hansen, K.B.; Wollmuth, L.P.; Bowie, D.; Furukawa, H.; Menniti, F.S.; Sobolevsky, A.I.; Swanson, G.T.; Swanger, S.A.; Greger, I.H.; Nakagawa, T.; et al. Structure, Function, and Pharmacology of Glutamate Receptor Ion Channels. Pharmacol. Rev. 2021, 73, 298–487. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Dravid, S.M.; Yuan, H.; Traynelis, S.F. AMPA Receptors: Molecular Biology and Pharmacology. In The Curated Reference Collection in Neuroscience and Biobehavioral Psychology; Elsevier Science Ltd.: Amsterdam, The Netherlands, 2017; pp. 311–318. [Google Scholar] [CrossRef]
- Fukumoto, K.; Fogaça, M.V.; Liu, R.J.; Duman, C.H.; Li, X.Y.; Chaki, S.; Duman, R.S. Medial PFC AMPA Receptor and BDNF Signaling Are Required for the Rapid and Sustained Antidepressant-like Effects of 5-HT(1A) Receptor Stimulation. Neuropsychopharmacology 2020, 45, 1725–1734. [Google Scholar] [CrossRef] [PubMed]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Barragan, E.V.; Wilson, P.C.; Burbach, K.F.; Soltanzadeh Zarandi, S.; Sood, A.; Paddy, M.R.; et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Berthoux, C.; De Bundel, D.; Valjent, E.; Bockaert, J.; Marin, P.; Bécamel, C. Presynaptic Serotonin 2A Receptors Modulate Thalamocortical Plasticity and Associative Learning. Proc. Natl. Acad. Sci. USA 2016, 113, E1382–E1391. [Google Scholar] [CrossRef]
- Berthoux, C.; Barre, A.; Bockaert, J.; Marin, P.; Bécamel, C. Sustained Activation of Postsynaptic 5-HT2A Receptors Gates Plasticity at Prefrontal Cortex Synapses. Cereb. Cortex 2019, 29, 1659–1669. [Google Scholar] [CrossRef]
- Riga, M.S.; Lladó-Pelfort, L.; Artigas, F.; Celada, P. The Serotonin Hallucinogen 5-MeO-DMT Alters Cortico-Thalamic Activity in Freely Moving Mice: Regionally-Selective Involvement of 5-HT1A and 5-HT2A Receptors. Neuropharmacology 2018, 142, 219–230. [Google Scholar] [CrossRef]
- Scruggs, J.L.; Patel, S.; Bubser, M.; Deutch, A.Y. DOI-Induced Activation of the Cortex: Dependence on 5-HT 2A Heteroceptors on Thalamocortical Glutamatergic Neurons. J. Neurosci. 2000, 20, 8846–8852. [Google Scholar] [CrossRef]
- Riga, M.S.; Soria, G.; Tudela, R.; Artigas, F.; Celada, P. The Natural Hallucinogen 5-MeO-DMT, Component of Ayahuasca, Disrupts Cortical Function in Rats: Reversal by Antipsychotic Drugs. Int. J. Neuropharmacol. 2014, 17, 1269–1282. [Google Scholar] [CrossRef] [PubMed]
- Takei, N.; Numakawa, T.; Kozaki, S.; Sakai, N.; Endo, Y.; Takahashi, M.; Hatanaka, H. Brain-Derived Neurotrophic Factor Induces Rapid and Transient Release of Glutamate through the Non-Exocytotic Pathway from Cortical Neurons. J. Biol. Chem. 1998, 273, 27620–27624. [Google Scholar] [CrossRef] [PubMed]
- Carbonaro, T.M.; Eshleman, A.J.; Forster, M.J.; Cheng, K.; Rice, K.C.; Gatch, M.B. The Role of 5-HT2A, 5-HT 2C and MGlu2 Receptors in the Behavioral Effects of Tryptamine Hallucinogens N,N-Dimethyltryptamine and N,N-Diisopropyltryptamine in Rats and Mice. Psychopharmacology 2015, 232, 275–284. [Google Scholar] [CrossRef] [PubMed]
- González-Maeso, J.; Ang, R.L.; Yuen, T.; Chan, P.; Weisstaub, N.V.; López-Giménez, J.F.; Zhou, M.; Okawa, Y.; Callado, L.F.; Milligan, G.; et al. Identification of a Serotonin/Glutamate Receptor Complex Implicated in Psychosis. Nature 2008, 452, 93–97. [Google Scholar] [CrossRef] [PubMed]
- West, W.B.; Lou, A.; Pechersky, K.; Chachich, M.E.; Appel, J.B. Antagonism of a PCP Drug Discrimination by Hallucinogens and Related Drugs. Neuropsychopharmacology 2000, 22, 618–625. [Google Scholar] [CrossRef][Green Version]
- Bäckman, L.; Nyberg, L.; Lindenberger, U.; Li, S.-C.; Farde, L. The Correlative Triad among Aging, Dopamine, and Cognition: Current Status and Future Prospects. Neurosci. Biobehav. Rev. 2006, 30, 791–807. [Google Scholar] [CrossRef]
- Nestler, E.J.; Carlezon, W.A. The Mesolimbic Dopamine Reward Circuit in Depression. Biol. Psychiatry 2006, 59, 1151–1159. [Google Scholar] [CrossRef]
- Jin, M.; Min, C.; Zheng, M.; Cho, D.-I.; Cheong, S.-J.; Kurose, H.; Kim, K.-M. Multiple Signaling Routes Involved in the Regulation of Adenylyl Cyclase and Extracellular Regulated Kinase by Dopamine D2 and D3 Receptors. Pharmacol. Res. 2013, 67, 31–41. [Google Scholar] [CrossRef]
- Lobo, M.K.; Covington, H.E.; Chaudhury, D.; Friedman, A.K.; Sun, H.; Damez-Werno, D.; Dietz, D.M.; Zaman, S.; Koo, J.W.; Kennedy, P.J.; et al. Cell Type–Specific Loss of BDNF Signaling Mimics Optogenetic Control of Cocaine Reward. Science 2010, 330, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Undieh, A.S. Pharmacology of Signaling Induced by Dopamine D1-like Receptor Activation. Pharmacol. Ther. 2010, 128, 37–60. [Google Scholar] [CrossRef] [PubMed]
- Francardo, V.; Bez, F.; Wieloch, T.; Nissbrandt, H.; Ruscher, K.; Cenci, M.A. Pharmacological Stimulation of Sigma-1 Receptors Has Neurorestorative Effects in Experimental Parkinsonism. Brain 2014, 137, 1998–2014. [Google Scholar] [CrossRef]
- Wilke, S.A.; Lavi, K.; Byeon, S.; Donohue, K.C.; Sohal, V.S. Convergence of Clinically Relevant Manipulations on Dopamine-Regulated Prefrontal Activity Underlying Stress Coping Responses. Biol. Psychiatry 2022, 91, 810–820. [Google Scholar] [CrossRef]
- Bergamini, G.; Sigrist, H.; Ferger, B.; Singewald, N.; Seifritz, E.; Pryce, C.R. Depletion of Nucleus Accumbens Dopamine Leads to Impaired Reward and Aversion Processing in Mice: Relevance to Motivation Pathologies. Neuropharmacology 2016, 109, 306–319. [Google Scholar] [CrossRef]
- Salamone, J.D.; Correa, M.; Yohn, S.; Lopez Cruz, L.; San Miguel, N.; Alatorre, L. The Pharmacology of Effort-Related Choice Behavior: Dopamine, Depression, and Individual Differences. Behav. Process. 2016, 127, 3–17. [Google Scholar] [CrossRef]
- Liu, R.; Wang, Y.; Chen, X.; Zhang, Z.; Xiao, L.; Zhou, Y. Anhedonia Correlates with Functional Connectivity of the Nucleus Accumbens Subregions in Patients with Major Depressive Disorder. NeuroImage: Clin. 2021, 30, 102599. [Google Scholar] [CrossRef]
- Kim, H.; Sablin, S.O.; Ramsay, R.R. Inhibition of Monoamine Oxidase A by Beta-Carboline Derivatives. Arch. Biochem. Biophys. 1997, 337, 137–142. [Google Scholar] [CrossRef]
- Elhwuegi, A.S. Central Monoamines and Their Role in Major Depression. Prog. Neuro-Psychopharm Biol. Psychiatry 2004, 28, 435–451. [Google Scholar] [CrossRef]
- De Castro-Neto, E.F.; da Cunha, R.H.; da Silveira, D.X.; Yonamine, M.; Gouveia, T.L.F.; Cavalheiro, E.A.; Amado, D.; Naffah-Mazzacoratti, M.D.G. Changes in Aminoacidergic and Monoaminergic Neurotransmission in the Hippocampus and Amygdala of Rats after Ayahuasca Ingestion. World J. Biol. Chem. 2013, 4, 141. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Polit, A.; Kędracka-Krok, S.; Wędzony, K.; Maćkowiak, M.; Dziedzicka-Wasylewska, M. Hetero-Dimerization of Serotonin 5-HT2A and Dopamine D2 Receptors. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2010, 1803, 1347–1358. [Google Scholar] [CrossRef] [PubMed]
- Lambe, E.K.; Aghajanian, G.K. Prefrontal Cortical Network Activity: Opposite Effects of Psychedelic Hallucinogens and D1/D5 Dopamine Receptor Activation. Neuroscience 2007, 145, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Frankel, P.S.; Cunningham, K.A. The Hallucinogen D-Lysergic Acid Diethylamide (d-LSD) Induces the Immediate-Early Gene c-Fos in Rat Forebrain. Brain Res. 2002, 958, 251–260. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L.; Leech, R.; Williams, T.M.; Erritzoe, D.; Abbasi, N.; Bargiotas, T.; Hobden, P.; Sharp, D.J.; Evans, J.; Feilding, A.; et al. Implications for Psychedelic-Assisted Psychotherapy: Functional Magnetic Resonance Imaging Study with Psilocybin. Br. J. Psychiatry 2012, 200, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Komorowski, J.; Stepień, H. The role of the endocannabinoid system in the regulation of endocrine function and in the control of energy balance in humans. Postep. Hig. Med. Dosw. 2007, 61, 99–105. [Google Scholar]
- Lowe, H.; Toyang, N.; Steele, B.; Bryant, J.; Ngwa, W. The Endocannabinoid System: A Potential Target for the Treatment of Various Diseases. Int. J. Mol. Sci. 2021, 22, 9472. [Google Scholar] [CrossRef] [PubMed]
- Reggio, P.H. Endocannabinoid Binding to the Cannabinoid Receptors: What Is Known and What Remains Unknown. Curr. Med. Chem. 2010, 17, 1468–1486. [Google Scholar] [CrossRef]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-Mediated Control of Synaptic Transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef]
- Felder, C.C.; Joyce, K.E.; Briley, E.M.; Mansouri, J.; Mackie, K.; Blond, O.; Lai, Y.; Ma, A.L.; Mitchell, R.L. Comparison of the Pharmacology and Signal Transduction of the Human Cannabinoid CB1 and CB2 Receptors. Mol. Pharmacol. 1995, 48, 443–450. [Google Scholar]
- Mackie, K.; Lai, Y.; Westenbroek, R.; Mitchell, R. Cannabinoids Activate an Inwardly Rectifying Potassium Conductance and Inhibit Q-Type Calcium Currents in AtT20 Cells Transfected with Rat Brain Cannabinoid Receptor. J. Neurosci. 1995, 15, 6552–6561. [Google Scholar] [CrossRef]
- Bouaboula, M.; Poinot-Chazel, C.; Marchand, J.; Canat, X.; Bourrié, B.; Rinaldi-Carmona, M.; Calandra, B.; Le Fur, G.; Casellas, P. Signaling Pathway Associated with Stimulation of CB2 Peripheral Cannabinoid Receptor. Involvement of Both Mitogen-Activated Protein Kinase and Induction of Krox-24 Expression. Eur. J. Biochem. 1996, 237, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Bouaboula, M.; Desnoyer, N.; Carayon, P.; Combes, T.; Casellas, P. Gi Protein Modulation Induced by a Selective Inverse Agonist for the Peripheral Cannabinoid Receptor CB2: Implication for Intracellular Signalization Cross-Regulation. Mol. Pharmacol. 1999, 55, 473–480. [Google Scholar] [PubMed]
- Guzmán, M.; Sánchez, C.; Galve-Roperh, I. Control of the Cell Survival/Death Decision by Cannabinoids. J. Mol. Med. 2001, 78, 613–625. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Functional Expression of Brain Neuronal CB2 Cannabinoid Receptors Are Involved in the Effects of Drugs of Abuse and in Depression. Ann. N. Y. Acad. Sci. 2008, 1139, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.N.; Osorio, F.L.; Morgan, C.J.A.; Crippa, J.A.S.; Bouso, J.C.; Rocha, J.M.; Zuardi, A.W.; Hallak, J.E.C.; Santos, R.G.D. The Effects of Cannabidiol (CBD) and Delta-9-Tetrahydrocannabinol (THC) on the Recognition of Emotions in Facial Expressions: A Systematic Review of Randomized Controlled Trials. Neurosci. Biobehav. Rev. 2020, 118, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Scherma, M.; Masia, P.; Satta, V.; Fratta, W.; Fadda, P.; Tanda, G. Brain Activity of Anandamide: A Rewarding Bliss? Acta Pharmacol. Sin. 2019, 40, 309–323. [Google Scholar] [CrossRef]
- Best, A.R.; Regehr, W.G. Serotonin Evokes Endocannabinoid Release and Retrogradely Suppresses Excitatory Synapses. J. Neurosci. 2008, 28, 6508–6515. [Google Scholar] [CrossRef]
- Parrish, J.C.; Nichols, D.E. Serotonin 5-HT(2A) Receptor Activation Induces 2-Arachidonoylglycerol Release through a Phospholipase c-Dependent Mechanism. J. Neurochem. 2006, 99, 1164–1175. [Google Scholar] [CrossRef]
- Gorzalka, B.B.; Hill, M.N.; Sun, J.C. Functional Role of the Endocannabinoid System and AMPA/Kainate Receptors in 5-HT2A Receptor-Mediated Wet Dog Shakes. Eur. J. Pharmacol. 2005, 516, 28–33. [Google Scholar] [CrossRef]
- Viñals, X.; Moreno, E.; Lanfumey, L.; Cordomí, A.; Pastor, A.; de La Torre, R.; Gasperini, P.; Navarro, G.; Howell, L.A.; Pardo, L.; et al. Cognitive Impairment Induced by Delta9-Tetrahydrocannabinol Occurs through Heteromers between Cannabinoid CB1 and Serotonin 5-HT2A Receptors. PLoS Biol. 2015, 13, e1002194. [Google Scholar] [CrossRef]
- Hashimoto, K. Sigma-1 Receptor Chaperone and Brain-Derived Neurotrophic Factor: Emerging Links between Cardiovascular Disease and Depression. Prog. Neurobiol. 2013, 100, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.K.; Dhir, A. Sigma-1 Receptors in Major Depression and Anxiety. Expert Rev. Neurother. 2009, 9, 1021–1034. [Google Scholar] [CrossRef] [PubMed]
- Sałaciak, K.; Pytka, K. Revisiting the Sigma-1 Receptor as a Biological Target to Treat Affective and Cognitive Disorders. Neurosci. Biobehav. Rev. 2022, 132, 1114–1136. [Google Scholar] [CrossRef] [PubMed]
- Collina, S.; Gaggeri, R.; Marra, A.; Bassi, A.; Negrinotti, S.; Negri, F.; Rossi, D. Sigma Receptor Modulators: A Patent Review. Expert Opin. Ther. Pat. 2013, 23, 597–613. [Google Scholar] [CrossRef]
- Szabo, A.; Kovacs, A.; Frecska, E.; Rajnavolgyi, E. Psychedelic N,N-Dimethyltryptamine and 5-Methoxy-N,N-Dimethyltryptamine Modulate Innate and Adaptive Inflammatory Responses through the Sigma-1 Receptor of Human Monocyte-Derived Dendritic Cells. PLoS ONE 2014, 9, e106533. [Google Scholar] [CrossRef]
- Szabo, A.; Frecska, E. Dimethyltryptamine (DMT): A Biochemical Swiss Army Knife in Neuroinflammation and Neuroprotection? Neural Regen. Res. 2016, 11, 396–397. [Google Scholar] [CrossRef]
- Inserra, A. Hypothesis: The Psychedelic Ayahuasca Heals Traumatic Memories via a Sigma 1 Receptor-Mediated Epigenetic-Mnemonic Process. Front. Pharmacol. 2018, 9, 330. [Google Scholar] [CrossRef]
- Morales-Garcia, J.A.; Calleja-Conde, J.; Lopez-Moreno, J.A.; Alonso-Gil, S.; Sanz-SanCristobal, M.; Riba, J.; Perez-Castillo, A. N,N-Dimethyltryptamine Compound Found in the Hallucinogenic Tea Ayahuasca, Regulates Adult Neurogenesis in Vitro and in Vivo. Transl. Psychiatry 2020, 10, 331. [Google Scholar] [CrossRef]
- Barker, S.A. N,N-Dimethyltryptamine (DMT), an Endogenous Hallucinogen: Past, Present, and Future Research to Determine Its Role and Function. Front. Neurosci. 2018, 12, 536. [Google Scholar] [CrossRef]
- Duval, F.; Mokrani, M.C.; Ortiz, J.A.; Schulz, P.; Champeval, C.; Macher, J.P. Neuroendocrine Predictors of the Evolution of Depression. Dialogues Clin. Neurosci. 2005, 7, 273–282. [Google Scholar] [CrossRef]
- Joyce, P.R. Neuroendocrine Changes in Depression. Aust. N. Z. J. Psychiatry 1985, 19, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Pariante, C.M.; Thomas, S.A.; Lovestone, S.; Makoff, A.; Kerwin, R.W. Do Antidepressants Regulate How Cortisol Affects the Brain? Psychoneuroendocrinology 2004, 29, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Akaltun, İ.; Çayır, A.; Kara, T.; Ayaydın, H. Is Growth Hormone Deficiency Associated with Anxiety Disorder and Depressive Symptoms in Children and Adolescents?: A Case-Control Study. Growth Horm. IGF Res. 2018, 41, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Algahtany, M.; Sharma, S.; Fahoum, K.; Jing, R.; Zhang, S.; Kovacs, K.; Rotondo, F.; Lee, J.; Vanek, I.; Cusimano, M.D. The Role of Growth Hormone in Depression: A Human Model. Front. Neurosci. 2021, 15, 661819. [Google Scholar] [CrossRef] [PubMed]
- Jackson, I.M. The Thyroid Axis and Depression. Thyroid 1998, 8, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, T.; Crown, A.; Checkley, S.; Farmer, A.; Lightman, S. Atypical Depression in Growth Hormone Deficient Adults, and the Beneficial Effects of Growth Hormone Treatment on Depression and Quality of Life. Eur. J. Endocrinol. 2004, 151, 325–332. [Google Scholar] [CrossRef][Green Version]
- Sullivan, P.F.; Wilson, D.A.; Mulder, R.T.; Joyce, P.R. The Hypothalamic-Pituitary-Thyroid Axis in Major Depression. Acta Psychiatr. Scand. 1997, 95, 370–378. [Google Scholar] [CrossRef]
- Schindler, E.A.D.; Wallace, R.M.; Sloshower, J.A.; D’Souza, D.C. Neuroendocrine Associations Underlying the Persistent Therapeutic Effects of Classic Serotonergic Psychedelics. Front. Pharmacol. 2018, 9, 177. [Google Scholar] [CrossRef]
- Calogero, A.E.; Bernardini, R.; Margioris, A.N.; Bagdy, G.; Gallucci, W.T.; Munson, P.J.; Tamarkin, L.; Tomai, T.P.; Brady, L.; Gold, P.W.; et al. Effects of Serotonergic Agonists and Antagonists on Corticotropin-Releasing Hormone Secretion by Explanted Rat Hypothalami. Peptides 1989, 10, 189–200. [Google Scholar] [CrossRef]
- Hemrick-Luecke, S.K.; Evans, D.C. Comparison of the Potency of MDL 100,907 and SB 242084 in Blocking the Serotonin (5-HT)(2) Receptor Agonist-Induced Increases in Rat Serum Corticosterone Concentrations: Evidence for 5-HT(2A) Receptor Mediation of the HPA Axis. Neuropharmacology 2002, 42, 162–169. [Google Scholar] [CrossRef]
- Mikkelsen, J.D.; Hay-Schmidt, A.; Kiss, A. Serotonergic Stimulation of the Rat Hypothalamo-Pituitary-Adrenal Axis: Interaction between 5-HT1A and 5-HT2A Receptors. Ann. N. Y. Acad. Sci. 2004, 1018, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Owens, M.J.; Knight, D.L.; Ritchie, J.C.; Nemeroff, C.B. The 5-Hydroxytryptamine2 Agonist, (+-)-1-(2,5-Dimethoxy-4-Bromophenyl)-2-Aminopropane Stimulates the Hypothalamic-Pituitary-Adrenal (HPA) Axis. I. Acute Effects on HPA Axis Activity and Corticotropin-Releasing Factor-Containing Neurons in the Rat Brain. J. Pharmacol. Exp. Ther. 1991, 256, 787–794. [Google Scholar] [PubMed]
- Hasler, F.; Grimberg, U.; Benz, M.A.; Huber, T.; Vollenweider, F.X. Acute Psychological and Physiological Effects of Psilocybin in Healthy Humans: A Double-Blind, Placebo-Controlled Dose-Effect Study. Psychopharmacology 2004, 172, 145–156. [Google Scholar] [CrossRef]
- Schmid, Y.; Enzler, F.; Gasser, P.; Grouzmann, E.; Preller, K.H.; Vollenweider, F.X.; Brenneisen, R.; Müller, F.; Borgwardt, S.; Liechti, M.E. Acute Effects of Lysergic Acid Diethylamide in Healthy Subjects. Biol. Psychiatry 2015, 78, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Strajhar, P.; Schmid, Y.; Liakoni, E.; Dolder, P.C.; Rentsch, K.M.; Kratschmar, D.V.; Odermatt, A.; Liechti, M.E. Acute Effects of Lysergic Acid Diethylamide on Circulating Steroid Levels in Healthy Subjects. J. Neuroendocrinol. 2016, 28, 12374. [Google Scholar] [CrossRef]
- Brierley, D.I.; Davidson, C. Developments in Harmine Pharmacology—Implications for Ayahuasca Use and Drug-Dependence Treatment. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 263–272. [Google Scholar] [CrossRef]
- Miralles, A.; Esteban, S.; Sastre-Coll, A.; Moranta, D.; Asensio, V.J.; García-Sevilla, J.A. High-Affinity Binding of Beta-Carbolines to Imidazoline I2B Receptors and MAO-A in Rat Tissues: Norharman Blocks the Effect of Morphine Withdrawal on DOPA/Noradrenaline Synthesis in the Brain. Eur. J. Pharmacol. 2005, 518, 234–242. [Google Scholar] [CrossRef]
- Göckler, N.; Jofre, G.; Papadopoulos, C.; Soppa, U.; Tejedor, F.J.; Becker, W. Harmine Specifically Inhibits Protein Kinase DYRK1A and Interferes with Neurite Formation. FEBS J. 2009, 276, 6324–6337. [Google Scholar] [CrossRef]
- Jain, S.; Panuganti, V.; Jha, S.; Roy, I. Harmine Acts as an Indirect Inhibitor of Intracellular Protein Aggregation. ACS Omega 2020, 5, 5620–5628. [Google Scholar] [CrossRef]
- Weiss, M.; Buldakova, S.; Dutova, E. Interaction of the Beta-Carboline Harmaline with a GABA-Benzodiazepine Mechanism: An Electrophysiological Investigation on Rat Hippocampal Slices. Brain Res. 1995, 695, 105–109. [Google Scholar] [CrossRef]
- Berrougui, H.; Martín-Cordero, C.; Khalil, A.; Hmamouchi, M.; Ettaib, A.; Marhuenda, E.; Herrera, M.D. Vasorelaxant Effects of Harmine and Harmaline Extracted from Peganum harmala, L. Seeds in Isolated Rat Aorta. Pharmacol. Res. 2006, 54, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Orsolini, L.; Pompili, S.; Tempia Valenta, S.; Salvi, V.; Volpe, U. C-Reactive Protein as a Biomarker for Major Depressive Disorder? Int. J. Mol. Sci. 2022, 23, 1616. [Google Scholar] [CrossRef] [PubMed]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and Depression: A Review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, G.C.; Gould, T.D.; Prueitt, W.L.; Nanavati, J.; Grunebaum, M.F.; Farber, N.B.; Singh, B.; Selvaraj, S.; Machado-Vieira, R.; Achtyes, E.D.; et al. Blood-Based Biomarkers of Antidepressant Response to Ketamine and Esketamine: A Systematic Review and Meta-Analysis. Mol. Psychiatry 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Clinical Trials Investigating the Antidepressant Effects of Ayahuasca | ||||
|---|---|---|---|---|
| Reference | Design | Sample | Main Measures | Main Findings |
| Osório et al., 2015 1 [37] | Preliminary Open-Label Study | Six Volunteers | HAM-D, MADRS, BPRS and YMRS | Reduction of 82% in depressive symptoms up to 21 days after administration |
| Sanches et al., 2016 2 [40] | Open-Label | Seventeen volunteers | HAM-D, MADRS, BPRS, CADSS and YMRS | Reduction in depressive symptoms up to 21 days after administration, increased blood flow in the left nucleus accumbens, right insula and left subgenual area |
| Galvão et al., 2018 [69] | Randomized, Placebo-Controlled Trial | Seventy-one volunteers (twenty-eight of whom ingested ayahuasca) | HAM-D, MADRS and salivary cortisol | Normalization of cortisol levels in saliva 48 hours after administration without correlation with reduction in depressive symptoms |
| Palhano-Fontes et al., 2019 3 [38] | Twenty-nine volunteers (fourteen of whom ingested ayahuasca) | HAM-D, MADRS, BPRS, CADSS, HRS, MEQ30 and YMRS | Reduction in depressive symptoms up to 7 days after administration | |
| Zeifman et al., 2019 [70] | MADRS and MADRS-SI | Non-significant reduction in suicidal ideation | ||
| de Almeida et al., 2019 [71] | Seventy-three volunteers (twenty-eight of whom ingested ayahuasca) | HAM-D, MADRS and plasma BDNF | Higher plasma BDNF concentrations in the ayahuasca group 48 hours after administration when compared with placebo. Increases in BDNF correlated with reduction in depressive symptoms | |
| Galvão-Coelho et al., 2020 [36] | HAM-D, MADRS, C-reactive protein and interleukin-6 | Reduction in C-reactive protein levels correlated with reduction in depressive symptoms | ||
| Reported Concentrations (mg/mL) | Number of Samples Analyzed | ||||
|---|---|---|---|---|---|
| Reference | DMT | Harmine | Harmaline | THH | |
| McKenna et al., 1984 [5] | 0.13–0.30 | 0.15–0.34 | 0–0.20 | 0.05–0.80 | 8 |
| Callaway, 2005 [78] | 0–14.15 | 0.45–22.85 | 0–0.90 | 0.48–23.8 | 29 |
| Santos et al., 2017 [79] | 0.30–36.70 1 | 20 | |||
| Souza et al., 2019 [80] | 0.62–3.40 | 4.14–18.16 | 0.40–3.92 | 4.02–30.88 | 38 |
| Santos et al., 2020 [81] | 0.10–3.12 | 0.11–7.11 | 0.01–0.94 | 0.09–3.05 | 33 |
| Kaasik et al., 2021 [82] | 0–2.68 | 0.06v4.44 | 0–0.33 | 0.01–3.87 | 102 |
| Reference | Standard DMT Dose (mg/kg) | Ayahuasca Alkaloid Concentration (mg/mL) | Mean Dosage per Mean Body Weight of Participants (mg) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| DMT | Harmine | Harmaline | THH | DMT | Harmine | Harmaline | THH | ||
| Riba et al., 2001 1 [83] | 0.50 | 0.53 | 0.90 | 0.06 | 0.72 | 35.75 | 60.65 | 4.12 | 48.75 |
| 0.75 | 53.63 | 90.97 | 6.14 | 74.42 | |||||
| 1.00 | 71.50 | 121.28 | 8.24 | 97.50 | |||||
| Riba et al., 2003 2 [11] | 0.60 | 39.80 | 67.40 | 4.60 | 54.20 | ||||
| 0.85 | 57.40 | 95.80 | 6.50 | 77.00 | |||||
| Riba et al., 2006 [39] | 1.00 | 66.80 | 113.31 | 7.69 | 91.09 | ||||
| dos Santos et al., 2011 [1] | 1.00 | 67.00 | 113.65 | 7.72 | 91.37 | ||||
| dos Santos et al., 2012 3 [84] | 0.75 | 52.26 | 88.64 | 6.02 | 71.26 | ||||
| Sanches et al., 2016 [40] | - | 0.80 | 0.21 | ND | NA | 128.00 | 33.50 | - | - |
| Palhano-Fontes et al., 2019 [38] | - | 0.36 | 1.86 | 0.24 | 1.20 | 25.77 | 133.15 | 19.32 | 85.00 |
| Rocha et al., 2021b [85] | - | 0.67 | 0.87 | 0.27 | 0.38 | 109.36 | 79.92 | 7.38 | 50.71 |
| dos Santos et al., 2021 [86] | - | 0.68 | 0.52 | 0.14 | 0.62 | 87.44 | 66.87 | 18.00 | 79.73 |
| Total mean | 0.81 | 0.61 | 0.87 | 0.18 | 0.73 | 66.23 | 88.76 | 8.70 | 74.64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, G.N.; Guerra, L.T.L.; Baker, G.B.; Dursun, S.M.; Saiz, J.C.B.; Hallak, J.E.C.; dos Santos, R.G. Molecular Pathways of the Therapeutic Effects of Ayahuasca, a Botanical Psychedelic and Potential Rapid-Acting Antidepressant. Biomolecules 2022, 12, 1618. https://doi.org/10.3390/biom12111618
Rossi GN, Guerra LTL, Baker GB, Dursun SM, Saiz JCB, Hallak JEC, dos Santos RG. Molecular Pathways of the Therapeutic Effects of Ayahuasca, a Botanical Psychedelic and Potential Rapid-Acting Antidepressant. Biomolecules. 2022; 12(11):1618. https://doi.org/10.3390/biom12111618
Chicago/Turabian StyleRossi, Giordano Novak, Lorena T. L. Guerra, Glen B. Baker, Serdar M. Dursun, José Carlos Bouso Saiz, Jaime E. C. Hallak, and Rafael G. dos Santos. 2022. "Molecular Pathways of the Therapeutic Effects of Ayahuasca, a Botanical Psychedelic and Potential Rapid-Acting Antidepressant" Biomolecules 12, no. 11: 1618. https://doi.org/10.3390/biom12111618
APA StyleRossi, G. N., Guerra, L. T. L., Baker, G. B., Dursun, S. M., Saiz, J. C. B., Hallak, J. E. C., & dos Santos, R. G. (2022). Molecular Pathways of the Therapeutic Effects of Ayahuasca, a Botanical Psychedelic and Potential Rapid-Acting Antidepressant. Biomolecules, 12(11), 1618. https://doi.org/10.3390/biom12111618